基于SRAP標記的國蘭種質資源遺傳多樣性分析
2020-06-19 08:53:50袁媛曹彬張詠琪陳清西陳南川
熱帶作物學報 2020年5期
袁媛 曹彬 張詠琪 陳清西 陳南川


摘? 要:采用SRAP(sequence related amplified polymorphism)技術對154份國蘭(雜交蘭)種質資源的親緣關系進行分析。從168對SRAP引物中篩選出16對條帶多、帶型清晰、多態性強的引物組合對所有樣品進行擴增,共獲得874條帶,其中多態性條帶857條,多態性比率為98.1%,平均每對引物每個樣品產生5.74條帶。UPGMA聚類分析表明:Me6-Em3引物能較好地揭示154份國蘭種質間的遺傳多樣性與親緣關系,在遺傳相似系數0.818處將它們分為8個類群,遺傳相似系數變化范圍為0.772~1.000。此外,發現引物Me8-Em4、Me11-Em2和Me12-Em11可以較可靠地聯合鑒定建蘭。該研究結果可為國蘭種質資源利用及雜種后代鑒定提供參考。
關鍵詞:國蘭;種質資源;SRAP標記;遺傳多樣性
中圖分類號:S682.31? ? ? 文獻標識碼:A
Genetic Diversity Analysis of Cymbidium Germplasms Based on SRAP Markers
YUAN Yuan1, CAO Bin1, ZHANG Yongqi1, CHEN Qingxi1*, CHEN Nanchuan2
1. College of Horticulture, Fujian Agriculture and Forestry University, Fuzhou, Fujian 350002, China; 2. Fujian Bainong Ecological Technology Co., Ltd, Zhaoan, Fujian 363500, China
Abstract: SRAP (sequence related amplified polymorphism) was applied for the determination of the diversity of 154 samples of Cymbidium germplasm. 16 SRAP primer pairs were screened from 168 SRAP primers and applied to all the materials, resulting in 874 amplified DNA bands. For the amplified DNA bands, 857 were polymorphic, with a ratio of 98.1%. The average number of bands of each SRAP primer for each Cymbidium germplasm was 5.74. UPGMA cluster analysis showed that the genetic diversity and genetic relationship of the Cymbidium germplasms could be revealed by Me6-Em3 primer pair and could be classified into eight groups with a similarity coefficient 0.818 and the genetic similarity of the germplasms ranged from 0.772 to 1.000. It was found for the first time that the primer pairs Me8-Em4, Me11-Em2 and Me12-Em11 could be used together to identify C. Ensifolium reliably. Results in the present study could provide references for the utilization of Cymbidium germplasm and the identification of Cymbidium hybrid cultivars.
Keywords: Chinese Cymbidium; germplasm; SRAP marker; genetic diversity
DOI: 10.3969/j.issn.1000-2561.2020.05.012
國蘭一般指中國蘭花,是蘭科(Orchidaceae)蘭屬(Cymbidium)草本植物,傳統上分為春蘭(C. goeringii)、建蘭(C. ensifolium)、墨蘭(C. sinense)、蕙蘭(C. faberi)、寒蘭(C. kanran)、春劍(C. longibracteatum)、蓮瓣蘭(C. tortisepalum)這7大類和其他國蘭雜交資源[1-2]。除傳統的國蘭7大類分類外,有學者將建蘭、墨蘭分為建蘭組,寒蘭、春蘭、春劍分在春蘭組[3],或將春蘭歸為春蘭組,建蘭、寒蘭、春劍和墨蘭歸為蕙組等[4]。雜交蘭是蘭屬地生蘭和附生蘭種間雜交培育而成的蘭花新種類,集地生蘭的植株小巧、幽香、典雅和附生蘭的花大、色純靚麗等優異特性,花期長,有花觀花、無花觀葉,觀賞價值和經濟價值高,逐漸成為消費市場的新寵,開發潛力巨大[5]。
在國蘭的育種工作中,由于種間及種內雜交頻繁,導致許多國蘭雜交品種的遺傳背景復雜,且形態學分類在一些雜交品種的分類中存在著很大的局限性[6],這就為研究其分類和遺傳多樣性工作帶來了很大的阻礙。此外,國蘭資源命名不規范,存在“同物異名”和“同名異物”現象,且中文名稱和拉丁文學名混亂,加上信息資源管理手段落后,限制了雜交蘭的種質資源保存、交流、利用和創新。在此背景下,分子標記技術逐漸成為一種精確、快速鑒定國蘭(雜交蘭)之間親緣關系的重要手段,包括SRAP、RAPD、AFLP、ISSR等[7-11]。其中,SRAP(sequence related amplified polymorphism)分子標記技術因高效、簡便、不需預知物種序列信息、引物設計簡單等特點[12],能提供的多態性信息比SSR分子標記更有效[13],比AFLP 分子標記更加優良[14],更適合國蘭鑒定[15],已應用于建蘭[16]、野生蘭[17]、雜交蘭[18]、寒蘭[19]等國蘭的研究中。
本研究采用SRAP 分子標記技術對154份國蘭(雜交蘭)樣品的親緣關系進行鑒定,并從中篩選3對引物作為聯合鑒定建蘭的標記引物,以期為種質資源鑒定、雜交育種中的親本選配和種質資源的合理利用提供科學依據與技術基礎。
1? 材料與方法
1.1? 材料
供試的154份國蘭樣品由福建百秾生態科技有限公司提供,樣品編號和雜交類型詳見表1。選取無病蟲害,生長正常的植株,取新鮮幼嫩葉片3~5片,保存在?80 ℃冰箱中。
1.2? 方法
1.2.1? 提取DNA? 使用高效植物基因組DNA提取試劑盒(天根生化科技有限公司)提取葉片DNA,使用微量紫外分光光度計測定濃度(UV- Vis Spectrophotometer Q5000,Quawell),用無菌ddH2O將DNA稀釋至50 ng/μL工作液,?20 ℃冰箱中保存備用。
1.2.2? SRAP引物篩選? 根據龔理等[20]在中國蘭SRAP-PCR研究中篩選引物的結果,選擇正向引物12條、反向引物14條(表2),共168對引物組合(鉑尚生物技術上海有限公司)。以提取的2個親緣關系近的國蘭材料K41和K180的DNA樣本為模板,分別用這168對引物組合進行PCR擴增,擴增產物用1.5%瓊脂糖凝膠在105 V恒壓電泳45 min,在凝膠成像儀下觀察、拍照保存分析,最后選出擴增結果條帶多、帶型清晰、多態性強的引物組合16對。
1.2.3? SRAP-PCR擴增反應? 用篩選的16對引物分別對154個國蘭葉片DNA模板進行SRAP- PCR擴增反應(儀器為Bio-Rad S1000TM Thermal Cycler PCR)。PCR反應體系共25 μL,包含2×Taq Master Mix(北京全式金生物技術有限公司)12.5 μL、DNA模板1 μL、正向引物(10 μmol/L)0.5 μL、反向引物(10 μmol/L)0.5 μL、無菌ddH2O 10.5 μL。PCR擴增程序為:94 ℃預變性5 min,反應前5個循環為94 ℃變性1 min,35 ℃退火1 min,72 ℃延伸2 min;后35個循環為94 ℃變性1 min,50 ℃退火1 min,72 ℃延伸2 min,循環結束后再72 ℃延伸10 min。反應結束后取10 μL PCR產物用1.5%瓊脂糖凝膠進行電泳檢測,電壓105 V,電泳時間45 min,電泳結束后在凝膠成像系統上(Bio-Rad SYSTEM Gel Doc XR+)采集圖像,拍照保存。
1.2.4? 建蘭鑒別引物的核驗? 通過16對引物擴增的圖譜中,獲得3對能較集中聚類建蘭的引物,推測它們能在將來鑒定建蘭中發揮一定作用,為核驗它們聯合鑒定建蘭的功能,隨機選取32個不同類別的國蘭樣品和1個建蘭種質(90號)作為對照,分別使用這3對引物組合對這33個國蘭樣品進行SRAP分析,制作聚類樹狀圖,然后統計3張聚類樹狀圖中與90號建蘭聚類的國蘭種質,再將這些種質取交集,查詢交集種質的來源是否為建蘭,從而驗證這3組引物鑒定建蘭的可靠性。
1.3? 數據處理
采用人工計數的方式讀取SRAP-PCR擴增的條帶,以DL2000 Marker為標準,在同一引物、同一位點有條帶記為“1”,無條帶記為“0”,統計結果用Excel 2010軟件制作(0,1)矩陣。通過NTSYS-pc 2.10軟件的SAHN Clustering分析(0,1)矩陣,使用UPGMA算法得到遺傳相似性系數(GS),并由軟件自動生成聚類樹狀圖。依次制作16對引物的擴增聚類圖譜,并從中篩選出能特異性鑒別建蘭的引物。
2? 結果與分析
2.1? 國蘭DNA提取和檢測
154份國蘭葉片DNA經過電泳檢測,條帶清晰完整(圖1);用紫外分光光度計檢測,濃度均在50 ng/μL以上;OD260/230比值1.8~2.0,OD260/280比值1.74~2.0,表明DNA完整性、濃度、純度均
符合實驗要求。
2.2? 國蘭SRAP多態性分析
從168對SRAP引物組合中,篩選出16對擴增條帶數目多、條帶清晰的引物組合,分別為Me5- Em14、Me6-Em3、Me7-Em3、Me8-Em2、Me8-Em4、Me8-Em5、Me8-Em8、Me8-Em13、Me8-Em15、Me9-Em3、Me11-Em2、Me11-Em4、Me11-Em15、Me12-Em9、Me12-Em11、Me12-Em12,用于全部國蘭樣品的SRAP-PCR擴增反應。
用篩選出的16對引物對154份國蘭種質進行SRAP-PCR擴增。圖2為其中一對引物的擴增圖譜。16對引物共擴增出54個位點,共獲得874條具有重復性的DNA帶,其中多態性條帶857條,多態性比率為98.1%。平均每對引物擴增每個樣品獲得5.74條。表明SRAP標記在國蘭種質中具有較為豐富的多態性,能較好地揭示國蘭材料間的親緣關系。
2.3? 遺傳相似性分析
154份國蘭種質遺傳相似系數(GS)在0.772~ 1.000區間內,說明大部分國蘭種質之間遺傳距離相對較近。12號‘K10-秋香與14號‘K44、100號‘福隆與101號‘雪蘭之間的遺傳相似系數最高,為1.000,表明它們的遺傳距離最近;67號‘金華山與12號‘K10-秋香的遺傳相似系數最低,為0.772,表明它們的遺傳距離較遠。
2.4? 國蘭樣品SRAP聚類分析
根據篩選出的16對多態性引物擴增得到SRAP擴增結果,利用NTSYS軟件進行UPGMA聚類分析,得出154份國蘭材料的親緣關系聚類圖(圖3)。在遺傳相似系數為0.818處將154份國蘭材料分為8個大類,第Ⅰ類是67號;第Ⅱ類是126號;第Ⅲ類包括46、48、89、93、47、87、102、88和115號材料,這些材料大部分含有春蘭的基因;第Ⅳ包括137號;第Ⅴ類包括128號;第Ⅵ類包括65、66、71和119號4份材料,其中65號‘花開富貴與66號‘S04-62聚類到一起,71號‘K33a-3和119號‘K30黃金小神童聚類到一起;第Ⅶ類包括105、45、104、106、110、112、118、108、109、111和113號材料;第Ⅷ類為剩下的所有國蘭材料。
在此基礎上,第Ⅷ類在相似系數為0.825處可劃分為A、B、C 3個亞類(圖3)。A類在相似系數0.860處劃分為6類,其中A1類包括1、5、9、12、14、15、16、7、86、68、84、97、132、82、91、99、100、101、103、6、27、73、125、146、29、142、83、85、127、145、26、74和43號材料;A2類包括79、81、143、78、95、94和96號材料;A3類包括69、70、72和90號材料;A4類包括2、124、3、77、13、18、25、130、24、75、4、11、129、22、8、10和23號材料,主要為建蘭或墨蘭后代;A5類包括17、148、44、98、147、49、133、19、135、76、20、50、138、80、144、136、21、121、123和141號材料;A6類包括56、60、57、59和131號材料。B類在相似系數0.860處可劃分為4類,B1類包括54號和153號兩份材料;B2類包括28、31、63、107、114、92和116號材料,除未知雜交類型的材料外,其余大部分為大花蕙蘭與其他類型蘭花的雜交后代;B3類包括30、32、37、51、38、40、41、150、33、34、39、149、58、122、62、35、154、151、42、36和64號材料;B4類為117號和120號。C類包括53、61、52、152、55、139、134和140號材料。
2.5? 聯合鑒別建蘭引物的核驗
在16對引物中,有3對引物的聚類分析結果顯示建蘭材料較為集中地聚集在一起,推測它們對建蘭的鑒定作用更顯著,進而推測聯合使用3對引物可以從不同國蘭材料中鑒別出建蘭及其后代。為了核驗這種推測,本研究使用3對引物Me8-Em4、Me11-Em2和Me12-Em11對33個國蘭樣品進行PCR擴增,其中90號建蘭‘金荷作為鑒別參照。在Me8-Em4鑒別聚類圖中(圖4),GS變化范圍為0.73~0.97,在GS為0.826時,與90號‘金荷聚為一類的樣品有:40、120、91、110、119、122、62、147、113、63、64和115號。
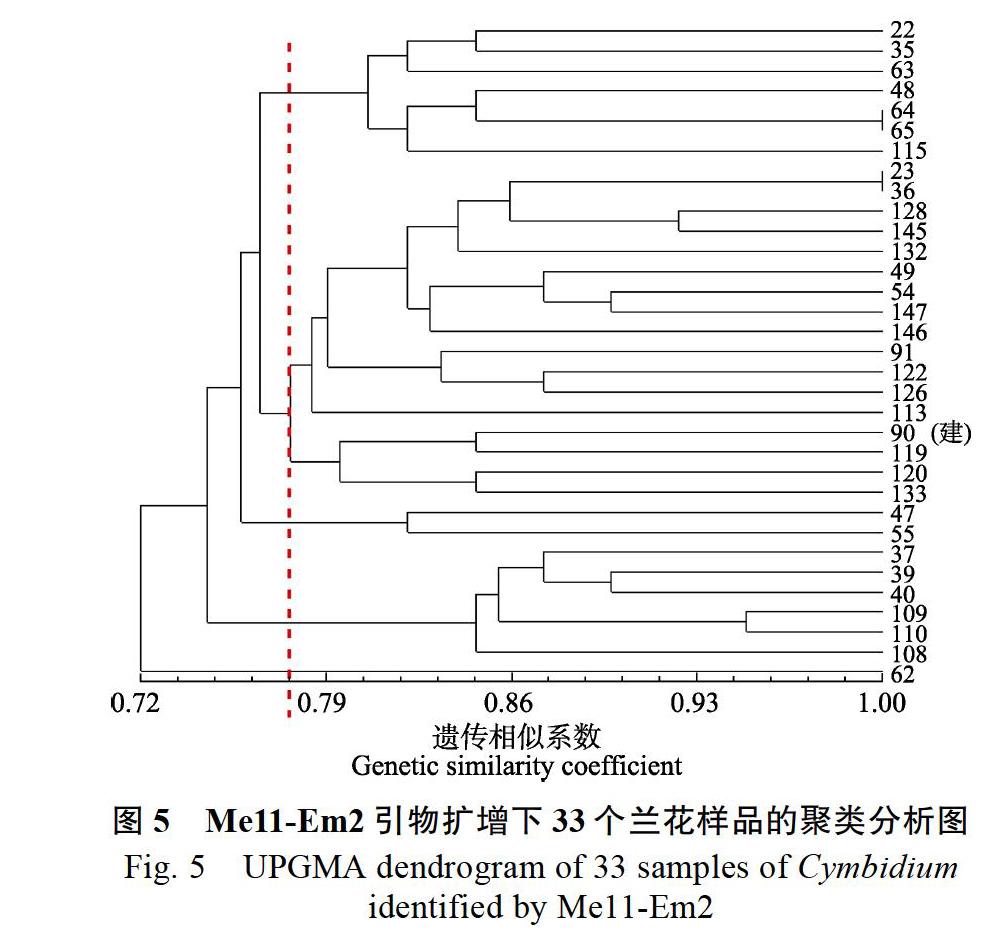
在Me11-Em2鑒別聚類圖(圖5)中,GS變化范圍為0.72~1.00,在GS為0.769時,與90號‘金荷聚為一類的樣品有:23、36、128、145、132、49、54、147、146、91、122、126、113、119、120和133號。
在Me12-Em11鑒別聚類圖(圖6)中,GS變化范圍為0.74~1.00,在GS為0.788時,與90號‘金荷聚為一類的樣品有:54、55、64、91、109、110、120和122號。
33個國蘭種質在3對不同引物的聚類分析下均與90號建蘭‘金荷聚類的是120、91和122號。通過查詢這3種國蘭的種質來源,發現三者確實為建蘭,在表型上也具有建蘭的特點,如葉帶型前部有鋸齒、花序直立并通常短于葉、總狀花序、花瓣狹長或橢圓等(圖7)。
3? 討論
3.1? SRAP技術能較為準確地鑒定國蘭的親緣關系
SRAP分子標記技術由于具備了AFLP分子標記的多態性高、可靠性強以及可重復性好的特點,同時具備了RAPD分子標記的隨機性、方便性和引物設計的簡便性等多方面的優勢,備受國內外研究者青睞[6],在花卉、草坪草、農作物、果樹中的應用中均發現其是一個較為穩定和準確的標記系統[21-27]。
本研究的結果也證實,根據已有的材料來源數據,SRAP分析得到的聚類結果能較真實地反應供試材料的親緣關系,如A4類中10號‘K24-1福韻丹霞與22號‘K26有相同的父本。A5中20號‘S1309-3和147號‘K02-68的父本都來自于‘K21紫妍氏,而‘K21紫妍氏與19號‘K21-1紫妍氏又有著較近的親緣關系,二者均為紅花,唇瓣帶胭脂紅斑,為大卷舌。A6類中57號‘K56紫妍氏三星蝶和59號‘K52綠翡翠爪藝都是墨蘭和大花蕙蘭雜交后代芽變而來,不同的是,前者是芽變花藝品,而后者是芽變葉藝品。B3類中150號‘(K26×C683)-7、34號‘(K26×C705)-11、62號‘(K26×C683)-83種雜交蘭的親本之一均為‘K26福韻紅霞,且35號‘K24福韻丹霞與‘K26福韻紅霞為兄弟株。C類中53號‘泗港水老葉無葉藝和61號‘泗港水聚類到一組,前者的老葉藝退化,而后者有葉藝。這個結果說明各種質間的親緣關系與來源、花色、花型、葉型、葉藝等均有一定的相關性,與鐘淮欽等[28]、牛田等[29]、馬紅勃等[30]分析國蘭花親緣關系的研究結果一致。79號建蘭材料‘一品梅和81號墨蘭材料‘C150-2、3號建蘭材料‘C428綠梅和77號墨蘭材料‘C387遺傳相似系數均為0.939,表現出較近的親緣關系,這與前人研究發現建蘭、墨蘭和寒蘭三者的親緣關系比較近,而春蘭與它們的親緣關系較遠[4, 17, 31]的結論一致。
盡管大部分結果與實際相吻合,但聚類分析出現一些目前尚無法解釋的結果,如97號墨蘭材料‘C387青葉達摩和132號春蘭材料‘C625帝冠聚為一簇;100號建蘭材料‘福隆和101號春蘭材料‘雪蘭聚為一簇,但它們在花型、葉型上的表現完全不同;在Me8-Em4聚類圖譜中,86號春劍雜交種和109號建蘭聚到一起,而目前并沒有研究證明春劍和建蘭有較近的親緣關系。這些結果可能是由于供試材料遺傳背景較為復雜,或植株出現芽變,或引物不夠特異等,其原因有待進一步研究。
3.2? SRAP技術可為雜種后代鑒定提供依據
在漫長且復雜的選育過程中,有些品種可能會由于記錄不夠完善導致丟失或不確定其分類類型或雜交類型,可通過SRAP進行聚類分析,推測其遺傳背景。例如A3類中70號‘C456-1和90號‘金荷均為建蘭,推測69號‘DF705B和72號‘L1120與建蘭親緣關系很近;65號‘花開富貴為春蘭雜交品種,119號‘K30黃金小神童為建蘭和大花蕙蘭雜交品種,推測66號‘S04-62可能含有春蘭基因,71號‘K33a-3可能含有建蘭或大花蕙蘭基因;26號‘K21-2先與墨蘭和大花蕙蘭的雜交后代74號‘K42 金吉利聚為一樹,再與同樣為墨蘭和大花蕙蘭的雜交后代的43號‘K32-3聚為一樹,推測26號‘K21-2也可能為墨蘭和大花蕙蘭的雜交后代;A4類中8號‘K06黃金三結義、10號‘K24-1福韻丹霞和22號‘K26福韻紅霞三者表現出較近的親緣關系,而10號‘K24-1福韻丹霞和22號‘K26福韻紅霞均為大花蕙蘭與墨蘭的雜交后代,且父本來源相同,推測8號‘K06黃金三結義極有可能為大花蕙蘭與墨蘭的雜交種;在A6類的五個品種中,56號‘K29黃金小神童爪藝、57號‘K56紫妍氏三星蝶和59號‘K52綠翡翠爪藝三者均為大花蕙蘭與其他類型國蘭的雜交后代,推測60號‘K40-1和131號‘K58也可能攜帶大花蕙蘭基因;在Me8-Em4聚類圖譜中,當GS為0.952時,70號‘C456-1先和75號‘L1101、79號‘一品梅、91號‘C1103聚到一起,再與76號‘C459小桃紅、94號‘DF402和125號‘C11大國冠聚到一起,其中除75號和125號分類未知外,其余70、79、91、76和94號均為建蘭,由此推測75號和125號可能也是建蘭或者含有建蘭親本的雜交蘭。當GS為0.968時,27號‘S457-11、61號‘泗港水、142號‘GS1001-1和146號‘幸姬與126號‘K31-2玉鳳之華聚類到一起。27號為建蘭,61號為墨蘭,二者聚到一起,說明二者具有一定的相似性且親緣關系相近,這與唐源江等[32]應用SRAP分子標記技術研究國蘭種質資源遺傳關系聚類結果相同,也與文李等[33]利用RAPD分子標記技術得出建蘭與墨蘭親緣關系最近,且兩者具有更強的親和性的結果相符。142號與146號為墨蘭種間雜交,126號為墨蘭和大花蕙蘭雜交,它們聚到一起,推測142號含有墨蘭基因。這些結果可為品種間的分類、雜種后代鑒定和雜交育種提供理論依據。
3.3? 基于SRAP技術利用多對引物聯合鑒定建蘭
根據UPGMA聚類結果,從16對引物中選擇出聚類較為清晰、明顯,且能較集中聚類建蘭的3對引物,即Me8-Em4、Me11-Em2和Me12-Em11。雖然這3對引物單獨使用時無法將所有建蘭及其后代聚集在一起來準確鑒定建蘭,但推測聯合使用3對引物將有望增加鑒定建蘭的準確性。為驗證這一猜想,使用3對引物分別對包括90號建蘭在內的33個蘭花種質(隨機選擇,包括春蘭、墨蘭、建蘭、寒蘭或其后代)進行聚類分析,取3對引物中和建蘭聚類到一起的交集品種,即可推斷其為建蘭,如3組引物中均與90號建蘭‘金荷聚類在一起的是120、91和122號,通過追溯他們的來源,發現它們的確屬于建蘭,表明利用這3對引物聯合鑒別建蘭確實具有一定的準確性和可靠性。相比通過單一引物對鑒定國蘭種質的方法,這種多對引物聯合鑒定的方法具有一定創新性,也為鑒定其他國蘭種質提供新的思路。鑒于本研究僅選取了33種國蘭材料,利用這3對引物聯合鑒定建蘭的嚴謹性還需要結合形態評價、增加樣本量等方法進行深入的研究。
4? 結論
本研究利用SRAP標記技術,對154份國蘭材料的親緣關系進行分析,從聚類結果來看,154份國蘭材料間表現出的遺傳背景比較復雜,具有豐富的遺傳多樣性,在遺傳相似系數0.818處可將154份供試材料劃分為8個類群,再對8個大類作細分,最終可將全部供試材料分為18類。SRAP標記在這些材料的親緣關系分析中表現的可信度較高,且可根據聚類結果推定未知材料的來源。此外,本研究發現,聯合3對SRAP引物Me8-Em4、Me11-Em2和Me12-Em11在鑒定建蘭及其后代上具有一定的可靠性和準確性。從本研究的結果來看,SRAP技術可為雜交蘭品種改良、分類命名及雜種后代鑒定等提供依據,多對引物聯合鑒定建蘭也具備一定的準確性和可靠性,需要進一步增大樣品數量以確認其嚴謹性。
參考文獻
敖素燕, 范義榮, 段艷皊, 等. 31個國蘭品種的數量性狀分析[J]. 江蘇農業科學, 2014, 42(9): 131-134.
嚴? 華, 張冬梅, 羅玉蘭, 等. 38種國蘭親緣關系的ISSR分析[J]. 分子植物育種, 2010, 8(4): 736-741.
Puy D D, Cribb P. The genus Cymbidium[M]. London, Portland, Oregan: Christophor Helm Timber Press, 1988.
吳應祥. 中國蘭花[M]. 2版. 北京: 中國林業出版社, 1993.
陸? 然. 雜交蘭: 生產漸成規模 產銷優勢明顯[J]. 中國花卉園藝, 2013(1): 23-24.
龔? 理. 部分國蘭品種(系)的SRAP遺傳多樣性研究[D]. 福州: 福建農林大學, 2014,
甘? 娜, 譚向紅, 陳其兵, 等. 應用RAPD標記和細胞質基因組PCR-RFLP技術研究大花蕙蘭的遺傳多樣性[J]. 園藝學報, 2006, 33(2): 349-355.
肖文芳, 李? 佐, 尤? 毅, 等. 蘭屬雜交蘭綠翡翠組培變異的RAPD檢測與鑒定[J]. 廣東農業科學, 2014, 41(21): 135-139.
田韋韋, 王彩霞, 田? 敏, 等. 文心蘭體細胞無性系變異的倍性檢測和CE-AFLP分析[J]. 核農學報, 2017, 31(2): 241-247.
張亞楠, 楊柏云, 熊冬金, 等. 寒蘭轉錄組SSR信息分析及其分子標記開發[J]. 南昌大學學報(理科版), 2017, 41(3): 249-254.
江亞雯, 孫小琴, 羅火林, 等. 基于ISSR標記的江西野生寒蘭居群遺傳多樣性研究[J]. 園藝學報, 2017, 44(10): 1993-2000.
Li G, Quiros C F. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in Brassica[J]. Theoretical and Applied Genetics, 2001, 103(2-3): 455-461.
Ahmad R, Potter D, Southwick S M. Genotyping of peach and nectarine cultivars with SSR and SRAP molecular markers[J]. Journal of the American Society for Horticultural Science, 2004, 129(2): 204-210.
Ferriol M, Picó B, Nuez F. Genetic diversity of a germplasm collection of Cucurbita pepo using SRAP and AFLP markers[J]. Theoretical and Applied Genetics, 2003, 107(2): 271-282.
隨粉粉. 福建國蘭種質資源遺傳多樣性研究[D]. 福州: 福建農林大學, 2010.
張? 芬, 李? 達, 呂長平, 等. 蘭花SRAP指紋圖譜的構建[J]. 湖南農業科學, 2011(3): 129-132.
楊光穗, 任? 羽, 王榮香. 利用SRAP技術分析海南野生蘭屬植物的親緣關系[J]. 熱帶農業科學, 2011, 31(9): 1-3.
何俊蓉, 蔣? 彧, 孫淑霞, 等. 13個國蘭品種遺傳關系的SRAP分析[J]. 分子植物育種(網絡版), 2011, 9(1): 1619-1625.
蹇? 黎, 朱利泉. 寒蘭品種類型的SRAP分子鑒定[J]. 中國農業科學, 2010, 43(15): 3184-3190.
龔? 理, 黃添毅, 王? 芳, 等. 國蘭雜交品種(系)SRAP- PCR反應體系優化及引物快速篩選[J]. 熱帶作物學報, 2014, 35(5): 925-932.
孫榮喜. 國槐種子形態變異與品種/無性系SRAP分子識別研究[D]. 北京: 中國林業科學研究院, 2011.
薛丹丹, 郭海林, 鄭軼琦, 等. 結縷草屬植物雜交后代雜種真實性鑒定——SRAP分子標記[J]. 草業學報, 2009, 18 (1): 72-79.
謝立峰, 李? 寧, 李? 燁, 等. 茄子種質遺傳多樣性及群體結構的SRAP分析[J]. 植物學報, 2019, 54(1): 58-63.
赫? 衛, 張? 慧, 董延龍, 等. 辣椒種質資源形態學性狀相關性、主成分與聚類分析[J]. 北方園藝, 2018(4): 9-17.
白? 羽, 丁海麥, 石松利. 瀕危植物蒙古扁桃SRAP反應體系優化[J]. 江蘇農業科學, 2019, 47(3): 46-48.
童? 俊, 毛? 靜, 董艷芳, 等. 鳶尾屬部分種質資源遺傳多樣性的SRAP分析[J]. 湖北農業科學, 2019, 58(4): 88-92.
林榕燕, 葉秀仙, 鐘淮欽, 等. 基于SRAP分子標記的石斛蘭種質資源遺傳多樣性分析[J]. 福建農業學報, 2018, 33(5): 469-473.
鐘淮欽, 林榕燕, 黃敏玲, 等. 雜交蘭種質資源遺傳多樣性的SRAP分析[J]. 福建農業學報, 2016, 31(11): 1193-1197.
牛? 田, 張? 林, 王厚新, 等. 利用SRAP標記分析春蘭種質資源遺傳多樣性[J]. 農學學報, 2014, 4(8): 53-58.
馬紅勃, 賴鋆英, 許旭明, 等. 基于SRAP標記的大花蕙蘭種質資源遺傳多樣性分析[J]. 植物遺傳資源學報, 2011, 12(4): 551-556.
孫彩云, 張明永, 葉秀粦, 等. 中國蘭屬植物種間及品種間親緣關系的RAPD分析[J]. 園藝學報, 2005, 32(6): 1121- 1124.
唐源江, 曹雯靜, 吳坤林. 基于SRAP標記的國蘭種質資源遺傳多樣性分析及分子身份證構建[J]. 中國農業科學, 2015, 48(9): 1795-1806.
文? 李, 葉慶生, 王小菁, 等. 利用RAPD技術分析蘭屬(Cymbidium)品種間的親緣關系[J]. 應用與環境生物學報, 2001(1): 29-32.
