廣西凡納濱對蝦源副溶血弧菌耐藥性及其整合子—基因盒檢測
2020-06-22 02:37:32賀曉晨韓書煜黎姍梅胡大勝黃德生吳偉鋒梁靜真胡庭俊
西南農業學報 2020年4期
關鍵詞:耐藥
賀曉晨,韓書煜,黎姍梅,胡大勝,黃德生,吳偉鋒,黃 鈞,梁靜真*,胡庭俊*
(1.廣西大學動物科學技術學院/廣西水生動物病害診斷實驗室,廣西 南寧 530004;2.廣西水產技術推廣總站,廣西 南寧 530022;3.合浦縣水產技術推廣站,廣西 北海 536100)
【研究意義】凡納濱對蝦(Litopenaeusvannamei)又名南美白對蝦,具有肉嫩味美、營養豐富、適鹽范圍廣、生長迅速和抗病能力強等優點,是全球主要對蝦養殖品種之一。但凡納濱對蝦弧菌性疾病在其養殖業迅猛發展的同時常暴發流行,導致養殖經濟損失巨大,嚴重制約凡納濱對蝦產業的可持續發展。副溶血弧菌(Vibrioparahaemolyticus)是一種廣泛分布于海水或內陸咸水的嗜溫嗜鹽革蘭氏陰性桿菌[1],屬于魚、蝦和貝等水生動物的條件性致病菌[2],可引起對蝦紅體、爛鰓及早期死亡綜合征等疾病,甚至造成對蝦大量死亡[3]。在生產中,由于養殖戶使用抗生素的方式方法不科學及養蝦塘受含有抗生素畜禽養殖廢水的污染,部分養蝦塘中的弧菌已產生耐藥性,嚴重威脅對蝦食用質量安全。因此,檢測分析凡納濱對蝦源副溶血弧菌的耐藥性及相關耐藥基因,對指導弧菌病防控和揭示其耐藥機制具有重要意義。【前人研究進展】Bakeraustin等[4]、Lopatek等[5]、Fattel等[6]分別對美洲、歐洲和亞洲的海水環境或水生動物源副溶血弧菌進行了耐藥性檢測。國內眾多學者也對我國20多個省(區)各種來源的副溶血弧菌進行了耐藥性調查[3,7-10]。以上研究結果表明,不同地區的副溶血弧菌耐藥性存在明顯差異,且部分菌株存在多重耐藥現象。細菌產生多重耐藥的主要原因是細其可通過某些途徑獲得外源耐藥基因[11]。整合子(根據整合酶不同可分為I、II和III型整合子)是一個能捕獲、整合、表達外源耐藥基因的位點特異性重組系統,能隨可移動遺傳組件(質粒和轉座子等)在細菌種內或種間進行水平轉移[12]。在細菌中以I型整合子最常見[13],其結構通常包含5′端保守區(5′-CS)、3′端保守區(3′-CS)及二者間的可變區,其中,5′-CS含有整合酶基因int1,大部分I型整合子的3′-CS含有季銨鹽耐藥基因qacEΔ1和磺胺類耐藥基因sul1,可變區通常可整合β-內酰胺類耐藥基因bla、氨基糖苷類耐藥基因aac和aad等各種耐藥基因盒[14]。國內外大量研究表明,不同來源弧菌的整合子攜帶率不同,其基因盒的攜帶情況也存在地域差異[14-19]。【本研究切入點】迄今,關于水產養殖源副溶血弧菌整合子分布的研究主要集中在我國海南和華東地區及秘魯和韓國等地區的分離株上[14-19],針對廣西凡納濱對蝦源副溶血弧菌中整合子—基因盒的研究鮮見報道。【擬解決的關鍵問題】以K-B紙片擴散法檢測2013-2018年104株廣西凡納濱對蝦源副溶血弧菌的耐藥性,采用PCR檢測分離株的整合子—基因盒攜帶情況,并分析整合子—基因盒與弧菌耐藥表型的相關性,為凡納濱對蝦副溶血弧菌病防控及該菌耐藥分子機制的深入研究提供基礎數據。
1 材料與方法
1.1 菌株來源
104株受試副溶血弧菌由廣西水生動物病害診斷實驗室于2013-2018年分離自廣西養殖發病的凡納濱對蝦,所有菌株均已進行API生化鑒定和16S rRNA分子鑒定。其中,36株分離自廣西北海市,命名為BH01~BH36;35株分離自廣西欽州市,命名為QZ01~QZ35;33株分離自廣西防城港市,命名為FCG01~FCG33。
1.2 主要試劑
PCR擴增引物參考姚小娟[14]的方法進行設計(表1),委托英濰捷基(上海)貿易有限公司合成。

表1 整合子—基因盒PCR擴增的引物序列、退火溫度及產物長度
注:CS 為保守區。
Note:CS is indicated as conserved segment.
2×TaqMasterMix和細菌基因組DNA快速提取試劑盒購自北京艾德萊生物科技有限公司。克隆載體pMD18-T和大腸桿菌(Escherichiacoli)DH5α感受態細胞購自寶日醫生物技術(北京)有限公司。瓊脂糖購自美國Invi-trogen公司。
試驗用藥敏紙片共16種,其中,新霉素(NEO,30 μg/片)、復方新諾明(SXT,磺胺甲惡唑/甲氧芐啶,23.75/1.25 μg/片)、諾氟沙星(NOR,10 μg/片)、環丙沙星(CLX,5 μg/片)、氧氟沙星(OFX,5 μg/片)、左氧氟沙星(LVX,5 μg/片)、恩諾沙星(ENR,5 μg/片)、多西環素(DOX,30 μg/片)和頭孢拉定(RAD,30 μg/片)等9種購自杭州天和微生物試劑有限公司,其余7種參考劉杰[20]的方法自制,分別為氟苯尼考(FFC,30 μg/片)、甲砜霉素(TAP,30 μg/片)、鹽酸沙拉沙星(SAR,5 μg/片)、磺胺二甲嘧啶(SM2,25 μg/片)、磺胺間甲氧嘧啶(SMM,25 μg/片)、磺胺對甲氧嘧啶(SMD,25 μg/片)和復方磺胺嘧啶(SD,25 μg/片)。所有藥敏紙片均置于4 ℃冷藏備用。
1.3 試驗方法
1.3.1 藥敏試驗 采用K-B紙片擴散法測定104株凡納濱對蝦源副溶血弧菌對16種抗菌藥物的敏感性,試驗結果根據美國臨床實驗室標準協會(CLSI)判定標準進行判定。
1.3.2 整合子—基因盒PCR檢測 按試劑盒說明提取細菌總DNA,并進行PCR擴增。PCR反應體系25.0 μl:2×TaqMasterMix 12.5 μl,上、下游引物各1.0 μl,DNA模板1.0 μl,ddH2O 9.5 μl。擴增程序:94 ℃預變性3 min;94 ℃ 30 s,退火30 s,72 ℃ 90 s,進行35個循環;72 ℃延伸10 min。采用1 %瓊脂糖凝膠電泳檢測PCR擴增產物。將回收純化后的PCR擴增產物與克隆載體pMD18-T連接,并轉化大腸桿菌DH5α感受態細胞。將陽性克隆送至深圳華大基因科技有限公司測序。
1.4 統計分析
采用SPSS 20.0對不同地區整合子—基因盒檢出率的差異性及整合子—基因盒與耐藥表型的相關性進行統計分析。
2 結果與分析
2.1 廣西凡納濱對蝦源副溶血弧菌的藥敏試驗結果
從圖1可看出,凡納濱對蝦源副溶血弧菌對FFC、LVX、OFX和NOR的敏感性較高,敏感率分別為82.7 %、79.8 %、73.1 %和71.2 %;對SMM、SM2、SD和SMD表現出較強的耐藥性,耐藥率分別為100.0 %、99.0 %、93.3 %和83.7 %;對其他抗菌藥物的敏感率在7.7 %~63.5 %。說明104株凡納濱對蝦源副溶血弧菌對16種抗菌藥物表現出不同程度的敏感性,其中對FFC、LVX、OFX和NOR較敏感,對SMM、SM2、SD和SMD較耐藥。
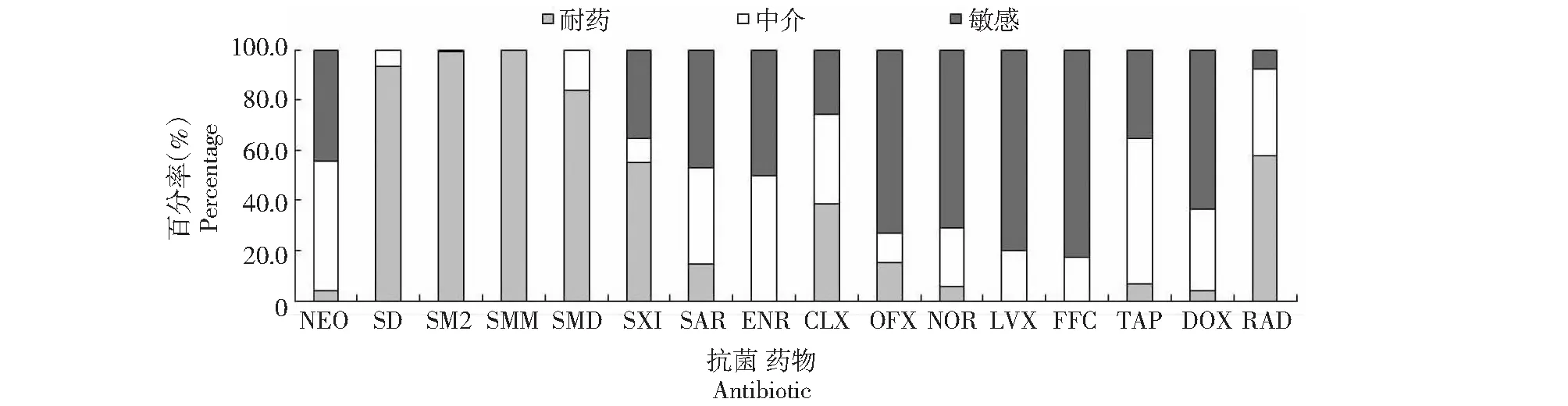
圖1 廣西凡納濱對蝦源副溶血弧菌對16種抗菌藥物的敏感性Fig.1 Susceptibility of V. parahaemolyticus isolated from L. vannamei in Guangxi towards 16 kinds of antibiotics
2.2 廣西凡納濱對蝦源副溶血弧菌的多重耐藥情況及耐藥譜
從圖2可看出,2013年的分離株主要為二重~五重耐藥,無八重及以上耐藥菌株,2014年以后,每年五重以上耐藥菌株所占比例均超過90.0 %,至2018年,新增九重耐藥菌株,八重和九重耐藥菌株數量合計占當年菌株總數的55.6 %。說明2013-2018年廣西凡納濱對蝦源副溶血弧菌多重耐藥狀況有明顯加重趨勢。
由表2可知,104株廣西凡納濱對蝦源副溶血弧菌的耐藥譜共20種,耐藥譜型豐富度為19.2 %。其中,優勢耐藥譜為SD/SM2/SMM/SMD/SXT/RAD,共17株,占菌株總數的16.3 %;二重耐藥株3株,占2.9 %;三重耐藥株8株,占7.7 %;四重耐藥株6株,占5.8 %;五重耐藥株33株,占31.7 %;六重耐藥株21株,占20.2 %;七重耐藥株14株,占總數的13.5 %;八重耐藥株16株,占15.4 %;九重耐藥株3株,占2.9 %。說明廣西凡納濱對蝦源副溶血弧菌具有一定的耐藥譜型豐富性。
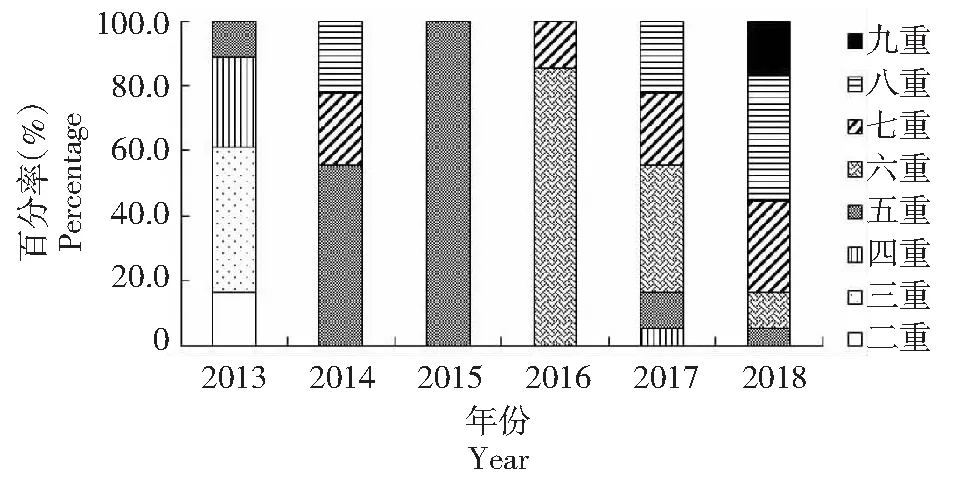
各年菌株數分別為:2013-2015、2017和2018年均為18株,2016年為14株 The strain amounts in each year are as follows:18 isolates in the year 2013-2015,2017 and 2018,14 isolates in the year 2016圖2 廣西凡納濱對蝦源副溶血弧菌的多重耐藥狀況Fig.2 Multidrug resistance status of V. parahaemolyticus isolated from L. vannamei in Guangxi
2.3 整合子—基因盒的檢出情況
經PCR檢測發現,104株廣西凡納濱對蝦源副溶血弧菌菌株中int1基因陽性菌株有67株,檢出率為64.4 %;經反復PCR檢測,所有菌株均未檢出int2和int3基因;sul1基因和qacEΔ1基因陽性菌株數分別有26和29株,檢出率分別為25.0 %和27.9 %;int1、sul1和qacEΔ1基因均為陽性的菌株有23株,即典型的I型整合子總檢出率為22.1 %。部分菌株int1、sul1和qacEΔ1基因的PCR擴增電泳結果見圖3。
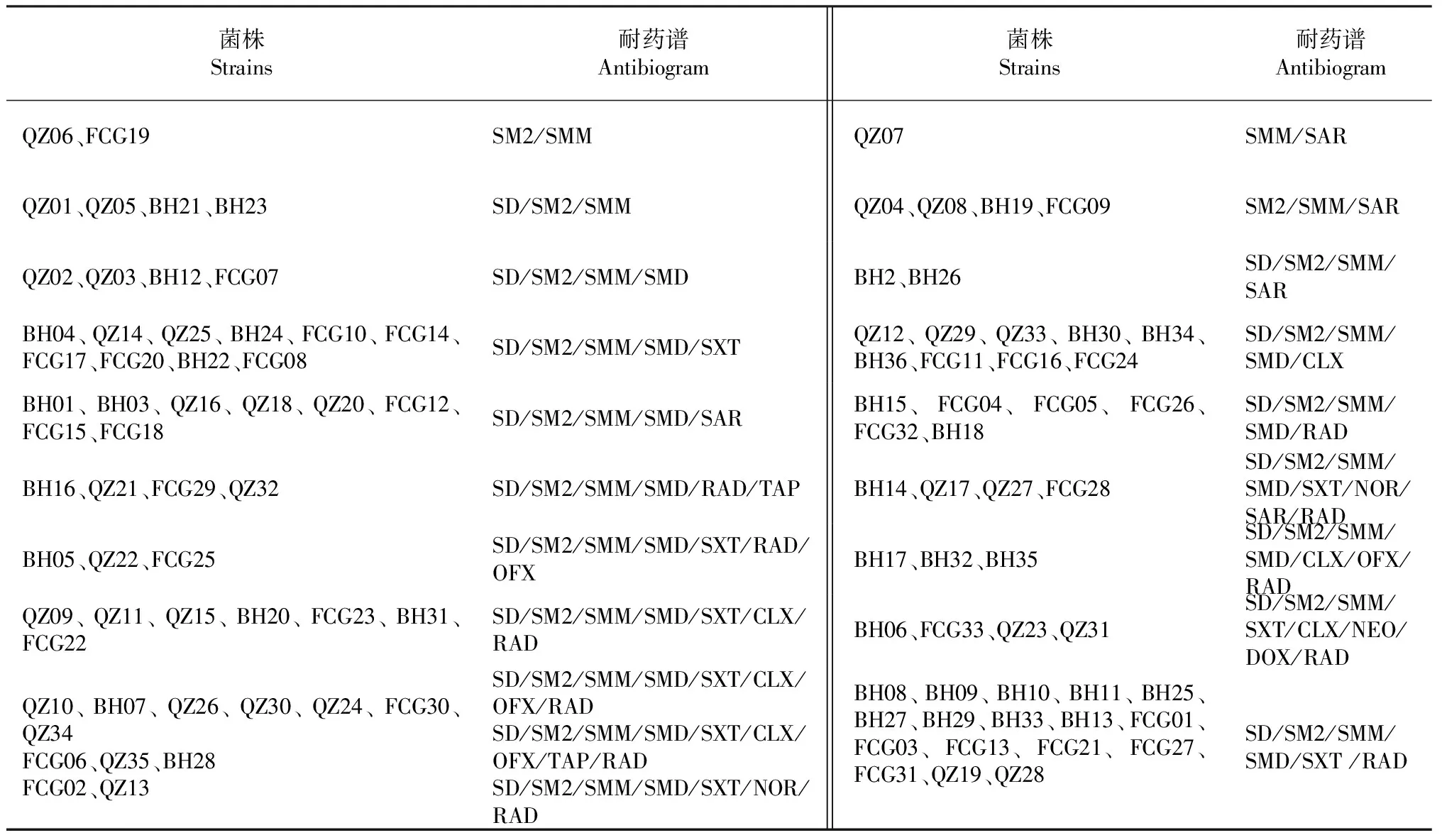
表2 廣西凡納濱對蝦源副溶血弧菌菌株的耐藥譜
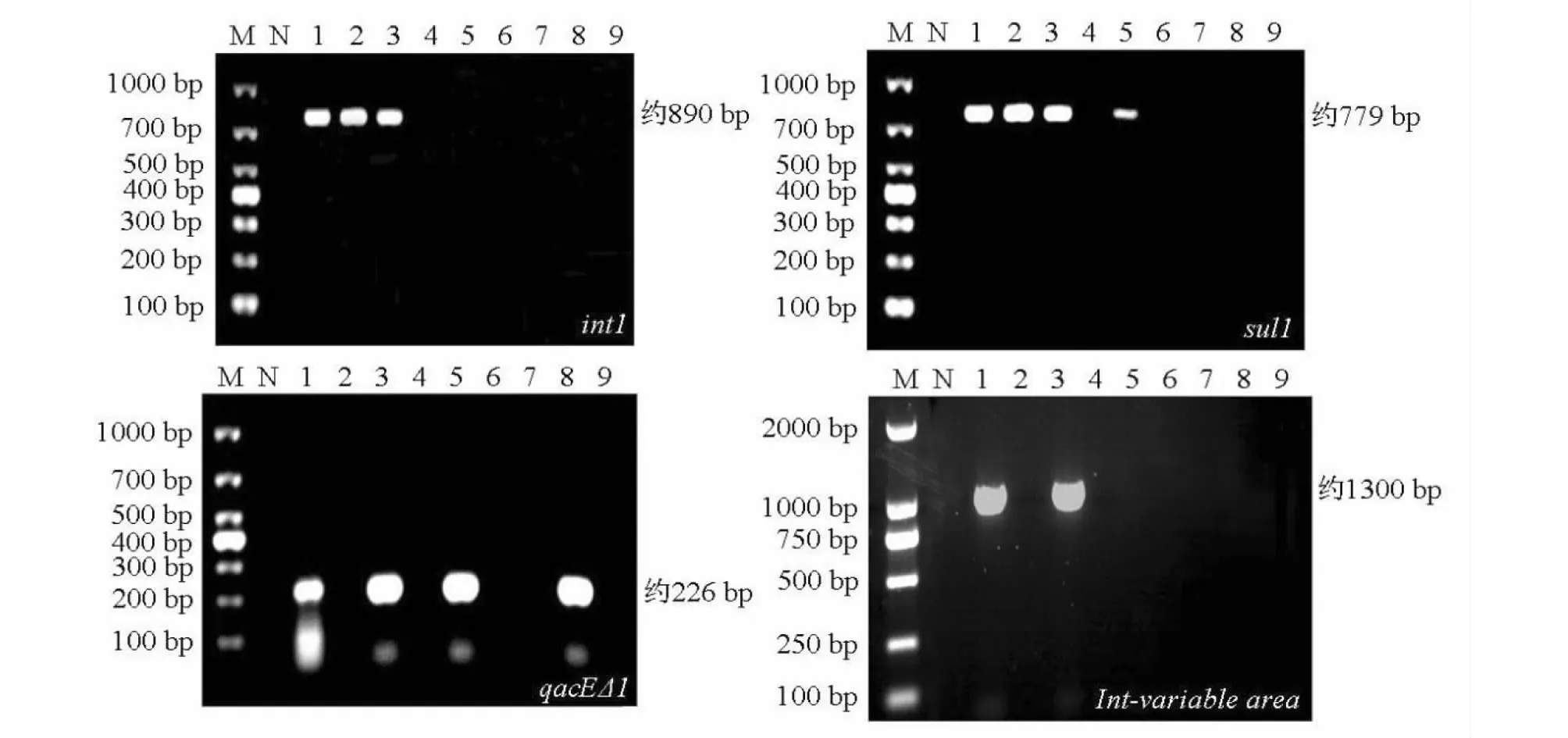
M:DL1000/2000 DNA Marker;N:陰性對照;1~9:被檢菌株M:DL1000/2000 DNA Marker;N:Negative control;1-9:Strains detected圖3 廣西凡納濱對蝦源副溶血弧菌菌株的整合子—基因盒PCR擴增電泳結果Fig.3 PCR amplification results of integron-gene cassettes of V. parahaemolyticus strains isolated from L. vannamei in Guangxi
對int1、sul1和qacEΔ1基因(I型整合子保守區)均為陽性的23株菌株進行I型整合子可變區PCR擴增,結果表明,在菌株BH15、BH18、BH25、FCG05、FCG18和QZ17中可檢測到片段長度約1300 bp的I型整合子可變區片段(圖3),檢出率為26.1 %。對該6株菌株可變區片段進行克隆和測序,將測序結果與GenBank中的對應序列進行比對分析,檢出blaCTX-M-2-aadA1(BH15、BH18、QZ17、FCG18)和blaVIM-60-aadA1-aacA(FCG05、BH25)2種基因盒。
由表3可知,凡納濱對蝦源副溶血弧菌int1基因在廣西北海市、欽州市和防城港市分離株中的檢出率依次為36.1 %、68.6 %和90.9 %,三者間差異顯著(P<0.05,下同);qacEΔ1基因在廣西北海市、欽州市和防城港市分離株中的檢出率依次為33.3 %、42.9 %和6.1 %,三者間差異極顯著(P<0.01);sul1基因和I型整合子可變區在這3個市分離株中的檢出率無顯著差異(P>0.05,下同)。說明I型整合子—基因盒在廣西凡納濱對蝦源副溶血弧菌菌株中的流行程度具有一定差異性。
2.4 I型整合子及基因盒與菌株耐藥性的相關性分析
由表4可知,int1陽性廣西凡納濱對蝦源副溶血弧菌菌株對TAP的耐藥率顯著高于int1陰性菌株;由于未檢測到相應的敏感、中介或耐藥菌株,凡納濱對蝦源副溶血弧菌對SMM、ENR、LVX、FFC的耐藥表型與int1基因的相關性無法計算;副溶血弧菌對其余抗菌藥物的耐藥表型與int1基因無顯著相關性。說明廣西凡納濱對蝦源副溶血弧菌菌株對大部分抗菌藥物的耐藥表型與I型整合子無明顯的相關性。
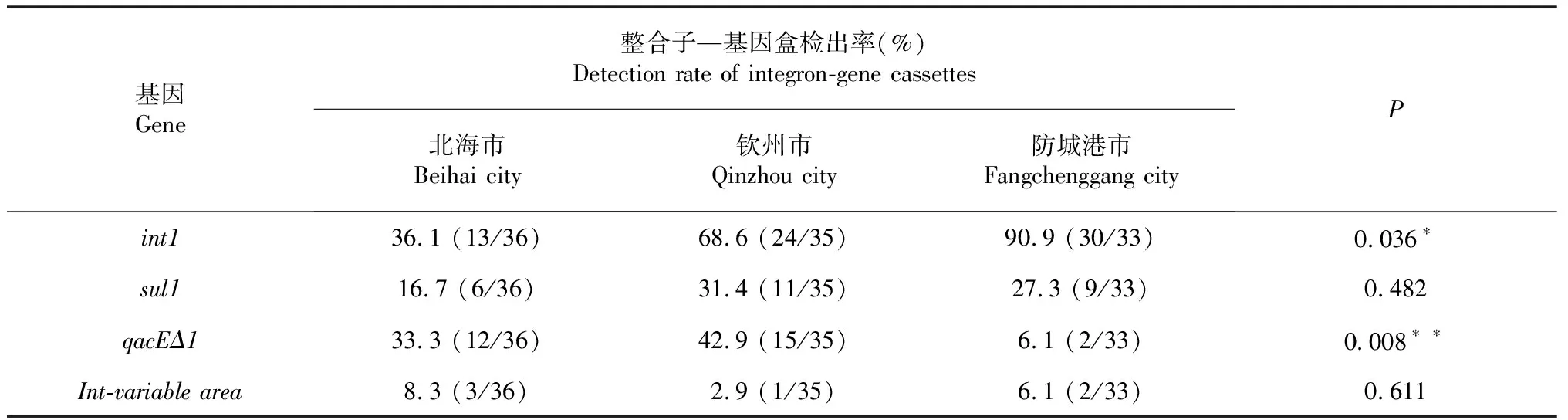
表3 廣西3個市凡納濱對蝦源副溶血弧菌菌株的整合子—基因盒檢出率比較
注:括號里的數字為攜帶相應基因的菌株數與受試菌株總數之比;*表示差異顯著(P<0.05),**表示差異極顯著(P<0.01)。
Note:Figures in the brackets indicate the amounts of strains harboring the relevant genes versus the total amounts of strains detected;* represented significant difference(P<0.05),**represented extremely significant difference(P<0.01).
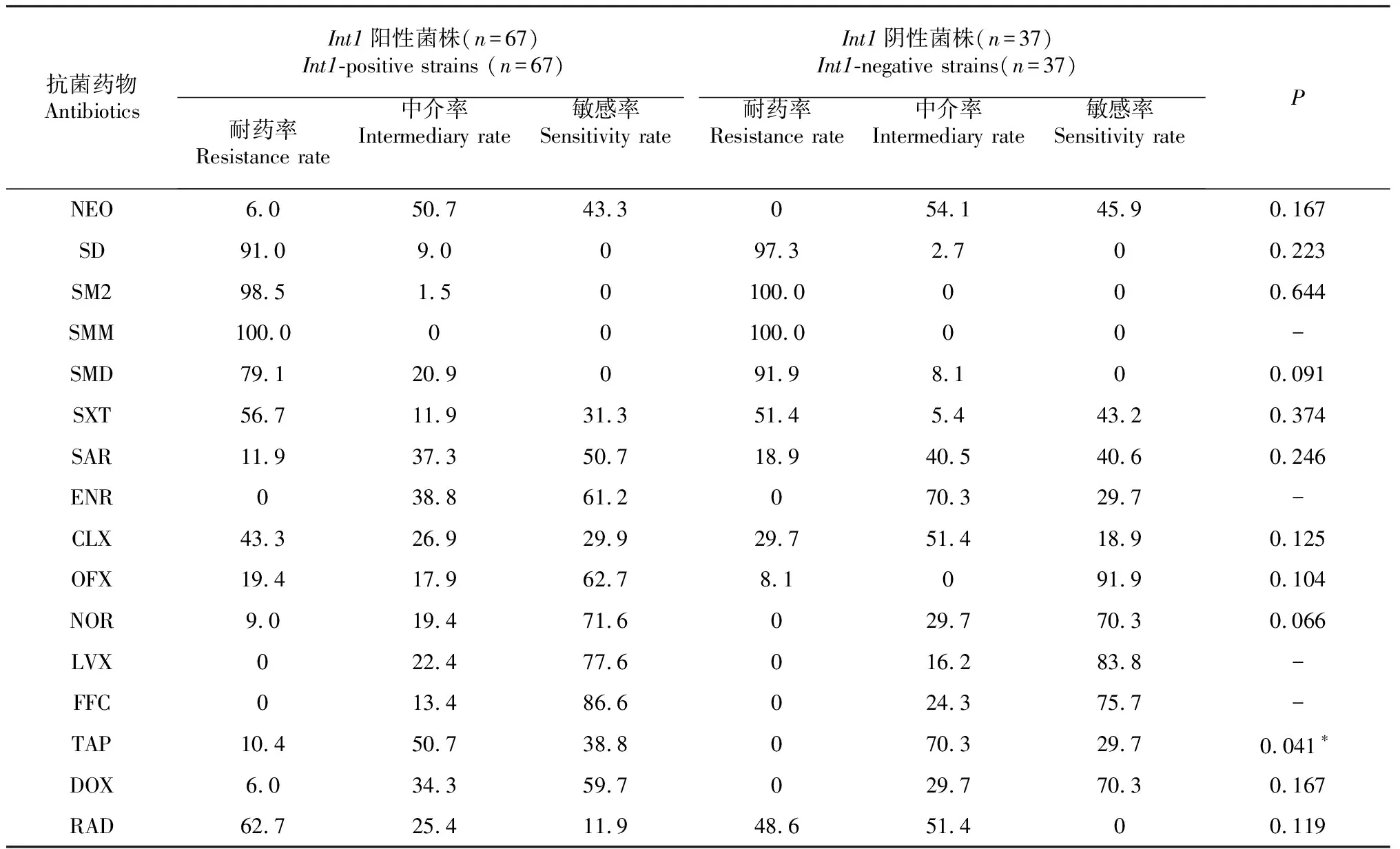
表4 int1陽性和int1陰性廣西凡納濱對蝦源副溶血弧菌菌株的耐藥性比較
注:-表示無法計算,下同;*表示差異顯著(P<0.05)。
Note:- indicates that the value could not be calculated,the same as below. * indicates significant difference (P<0.05).

表5 整合子相關基因陽性菌株和陰性菌株的耐藥性比較
在I型整合子可變區檢出基因盒的6株分離株(BH15、BH18、QZ17、FCG18、FCG05和BH25)均為五重或五重以上耐藥菌株(表2)。由表5可知,在int1、sul1和qacEΔ1基因均為陽性的23株廣西凡納濱對蝦源副溶血弧菌中,RAD的耐藥表型與基因盒blaCTX-M-2和blaVIM-60、5種磺胺類抗菌藥物的耐藥表型與sul1基因間的相關性均不顯著,NEO的耐藥表型與基因盒aadA1和aacA的相關性無法計算。說明在廣西凡納濱對蝦源副溶血弧菌菌株中各基因盒與相應抗菌藥物耐藥表型也無明顯的相關性。
3 討 論
3.1 廣西凡納濱對蝦源副溶血弧菌的耐藥性
抗菌藥物不規范使用導致水產養殖源弧菌多重耐藥性日益加重[5,18]。2013-2018年,廣西凡納濱對蝦源副溶血弧菌多重耐藥狀況有明顯加重趨勢,八重及八重以上耐藥菌株所占比例由2013年的無攀升到2018年的55.6 %。據調查顯示,廣西凡納濱對蝦養殖戶在養蝦過程中極少購買或使用抗生素,推測副溶血弧菌多重耐藥性的加重可能與養殖水體受醫院、生活或畜禽養殖污水中的抗生素污染或耐藥基因的傳播有關[3,21]。本研究中,受試菌株對大部分磺胺類的耐藥率均較高,與天津和浙江等地分離株的藥敏試驗結果相似[22-23]。磺胺類藥物是一種價格低廉的人獸共用藥,已廣泛用于臨床治療和畜禽養殖等,但長期不規范使用促使其頻繁滲入環境。目前,我國地表水環境中磺胺類藥物總體濃度遠高于其他國家[24],推測本研究分離株對磺胺類的耐藥性主要受到擴散至環境中的磺胺類所誘導。本研究結果表明,廣西凡納濱對蝦源副溶血弧菌對氟苯尼考敏感率(82.7 %)最高,但低于劉杰等[7](95.81 %)和黃偉德等[3](96.7 %)的檢測結果,可能與2017-2018年采集的菌株對氟苯尼考中介率相對較高有關,需給予高度重視并進一步調查具體原因。建議在對蝦養殖過程中:①盡可能不使用抗菌藥物進行疾病防控;②當弧菌病暴發必須使用抗菌藥物時,應根據藥敏結果選擇敏感抗菌藥物治療;③要避免低濃度或長期使用同種藥物,因為低濃度、長期使用同種藥物會誘導細菌產生耐藥性[25];④根據本研究的藥敏試驗結果,盡管氟苯尼考敏感率有所下降,其仍可考慮作為廣西凡納濱對蝦副溶血弧菌病防治的備選藥物。
3.2 廣西凡納濱對蝦源副溶血弧菌整合子—基因盒攜帶情況
本研究結果表明,I型整合子在廣西凡納濱對蝦源副溶血弧菌中具有一定的流行性,I型整合酶基因int1檢出率為64.4 %,低于Dahanayake等[19](90 %)、姚小娟[14](89.62 %)的檢出率,但高于劉旭[16](5.6 %)、李林桂[15](2.8 %)和Sulca[18](0)等的檢出率;I型整合子3′-CS的sul1基因和qacEΔ1基因檢出率分別為25.0 %和27.9 %,均高于其他研究報道的水產養殖源弧菌檢出率[14-16];I型整合子可變區的基因盒包括β-內酰胺類耐藥基因blaCTX-M-2和blaVIM-60及氨基糖苷類耐藥基因aadA1和aacA。其他水產養殖源弧菌的基因盒攜帶情況,江蘇、浙江、福建和海南分離株的基因盒以arr-3、dfr16、dfrA27和aadA2為主[14-15],趙姝等[17]在半滑舌鰨(Cynoglossussemilaevis)源弧菌中檢出dfr16、aadA2、arr-3和dfrA27等基因盒,韓國的菲律賓簾蛤(Ruditapesphilippinarum)源弧菌主要攜帶blaCTX-M、blaTEM和aadA2等基因盒[19]。本研究中,int1和qacEΔ1基因在廣西北海市、欽州市和防城港市菌株中的檢出率差異顯著,可能與不同地區蝦塘受含有int1基因和qacEΔ1基因的污水污染程度不同有關。已有報道,從醫院污水、豬魚混養池水中分離獲得的細菌均攜帶有I型整合子及相關基因盒,且污水中的I型整合子很難去除[26-27]。I型整合子的流行加劇了廣西凡納濱對蝦源副溶血弧菌產生多重耐藥的可能性,實行生態養殖、嚴格規范使用抗生素、避免養殖用水受污染等應是目前控制廣西凡納濱對蝦源副溶血弧菌中I型整合子及相關耐藥基因傳播的有效措施。
3.3 凡納濱對蝦源副溶血弧菌整合子—基因盒與耐藥表型的相關性
根據相關性分析結果可知,廣西凡納濱對蝦源副溶血弧菌int1陽性菌株對甲砜霉素的耐藥率顯著高于int1陰性株,說明凡納濱對蝦源副溶血弧菌對甲砜霉素的耐藥機制與I型整合子存在密切聯系。除甲砜霉素外,int1基因與其余抗菌藥物的耐藥表型間均未檢測到顯著相關性,與已報道的人醫臨床分離大腸桿菌[28-29]和銅綠假單胞菌(Pseudomonasaeruginesa)[30]存在差異。究其原因可能是蝦塘副溶血弧菌所受的抗生素選擇壓力比醫院臨床細菌小,其整合子捕獲和積累耐藥基因盒的概率也相對較小[28]。本研究中5種磺胺類耐藥表型與sul1基因間也無顯著相關性,應與sul1基因位于I型整合子的3′末端、其表達可能受到啟動子種類、強弱和距離的影響[31],以及分離株還可能存在其他磺胺類耐藥機制有關。qacEΔ1基因屬于季銨鹽類耐藥基因,本研究中攜帶qacEΔ1基因的凡納濱對蝦源副溶血弧菌是否對季銨鹽消毒劑耐藥尚需進一步探究證實;在int1、sul1和qacEΔ1基因均為陽性的23株凡納濱對蝦源副溶血弧菌中,基因盒blaCTX-M-2和blaVIM-60與頭孢拉定耐藥表型、基因盒aadA1和aacA與新霉素耐藥表型間無顯著相關性,但出現部分攜帶aadA1和aacA的菌株對新霉素敏感、攜帶blaCTX-M-2的菌株對頭孢拉定敏感的現象。類似現象在大腸桿菌的人類臨床分離株[28]和腹瀉犬源分離株中也有報道[32],可能是由耐藥基因的低水平表達或基因沉默所引起[28]。影響基因盒表達水平的因素包括可變區啟動子Pc和二級啟動子P2的強弱及基因盒與啟動子的距離遠近等,基因盒離啟動子越近則表達水平越高,反之則越低[31]。盡管本研究中凡納濱對蝦源副溶血弧菌對大部分抗菌藥物的耐藥表型與int1基因間未檢測到顯著相關性、基因盒與相應抗菌藥物的耐藥表型間相關性也不顯著,但整合子—基因盒的存在給廣西凡納濱對蝦副溶血弧菌疾病防控帶來的風險仍需引起高度重視。為了盡可能控制凡納濱對蝦源副溶血弧菌多重耐藥狀況加劇,建議在對蝦養殖過程中應加強對弧菌整合子—基因盒的監控,對整合子—基因盒的消除方法進行深入探討,嚴格控制抗生素的使用,避免養殖用水污染,以提高凡納濱對蝦副溶血弧菌病的防控效率。
4 結 論
廣西凡納濱對蝦源副溶血弧菌對16種抗菌藥物表現出不同程度的敏感性,其中對氟苯尼考的敏感性最高,可考慮將該抗菌藥物作為廣西凡納濱對蝦副溶血弧菌病防治的備選藥物。凡納濱對蝦源副溶血弧菌對大部分抗菌藥物的耐藥表型與int1基因間未檢測到顯著相關性、基因盒與相應抗菌藥物的耐藥表型間的相關性也不顯著,但I型整合子的流行加劇了廣西凡納濱對蝦源副溶血弧菌產生多重耐藥的可能性,因此應加強對弧菌整合子—基因盒的監控。
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
