萊州灣游泳動物群落結構研究
2020-06-28 00:24:40張煥君劉淑德劉元進周全利
海洋科學 2020年6期
張煥君, 劉淑德, 劉元進, 涂 忠, 周全利, 陳 瑋, 孫 偉, 李 凡
萊州灣游泳動物群落結構研究
張煥君1, 劉淑德2, 劉元進1, 涂 忠2, 周全利1, 陳 瑋1, 孫 偉1, 李 凡1
(1. 山東省海洋資源與環境研究院 山東省海洋生態修復重點實驗室, 山東 煙臺 264006; 2. 山東省水生生物資源養護管理中心, 山東 煙臺 264003)
本文根據2010和2011年春季(5月)、夏季(8月)、秋季(10月)和冬季(12月)進行的8航次調查數據, 研究了萊州灣游泳動物群落格局及其與環境因子關系。結果顯示, 調查共捕獲游泳動物85種, 2011年生物量略高于2010年; 生物量季節間差別較大, 各季節生物量排序為夏季、秋季、冬季和春季; 不同年間相同季節生物量差異多不顯著(秋季除外); 除2010年春季外, 其余季節均為魚類生物量最高, 甲殼類次之, 頭足類最低; 從全年調查平均值來看, 萊州灣西側游泳動物生物量(60.2 kg/h)明顯高于東側(33.4 kg/h)。根據8航次31個主要種類生物量聚類分析, 萊州灣20調查站位可劃分為4組: 組I由位于調查海域東南的3個站位組成, 組II由位于調查海域東部及中部的8個站位組成, 組III由調查海域西北部(黃河口南側)的3個站位組成, 組IV由西部6個站位組成。組I浮游植物豐度、浮游動物生物量、游泳動物生物量和游泳動物多樣性指數在各組中均最低, 而底棲動物生物量最高; 組III和組IV的浮游植物豐度、浮游動物生物量以及游泳動物生物量高于組I和組II。ANOSIM分析表明, 各聚類組間群落結構雖有不同, 但差異并不顯著。DCCA分析表明, 溫度、鹽度、底棲動物生物量以及浮游植物豐度是影響游泳動物群落的重要因子。研究表明, 目前萊州灣游泳動物群落總生物量處于較低水平, 特別是春季生物量下降尤為嚴重; 生物量的季節分布發生了較大的改變, 春季生物量極低, 夏季生物量相對較高; 底層魚類比例下降的趨勢有所逆轉, 頭足類比例明顯增加。萊州灣游泳動物群落格局異質性較低, 可能與萊州灣生境異質性低以及人類劇烈干擾密切相關。
生物量; 聚類; 非度量多維標度排序; 去趨勢典范對應分析
萊州灣位于山東半島西北、渤海南部, 有黃河、濰河、小清河等多條河流注入。黃河是影響本區域的主要河流, 每年為萊州灣帶來了大量的淡水, 入海口附近海域具有鹽度低, 溫度適中, 營養鹽豐富的特點, 使萊州灣附近海域成為眾多主要經濟生物的產卵場、索餌場和育幼場, 是我國沿海漁業資源生產力最高的海域之一, 俗有“北方漁倉”之美譽。受長期的過度捕撈和環境污染影響, 萊州灣乃至整個黃、渤海的漁業資源已嚴重衰退, 面臨幾近枯竭的嚴峻形勢, 漁業資源群落結構也發生了巨大的變化[1-3]。
萊州灣游泳動物主要包括魚類、頭足類和一些大型甲殼類, 它們中的許多種類既是海洋生態系統中的高級消費者, 也是漁業開發的對象。研究游泳動物群落格局及其變化對于了解萊州灣生態系統特征以及游泳動物資源保護具有重要現實意義。有關萊州灣及渤海游泳動物群落組成、優勢種變化等的研究開展較多[2, 4-10], 這些研究未涉及游泳動物群落格局的研究。
本研究于2010年和2011年對萊州灣漁業資源進行了8個航次的調查, 探究了萊州灣游泳動物生物量及其時空變化, 揭示了萊州灣游泳動物群落結構及其與環境因子的關系, 以期為近海漁業資源保護、漁業經濟可持續發展以及生態修復等提供基礎資料。
1 材料與方法
1.1 調查站位和調查時間
本研究于2010年和2011年對萊州灣附近海域進行了8航次調查, 調查時間分別為5月(春季)、8月(夏季)、10月(秋季)、12月(冬季), 在119°05′—120°00′E、37°12′—37°40′N區域內設置了20個站位(圖1), 采用單船底拖網, 船只功率260 kW, 網口周長30.6 m, 囊網網目20 mm, 拖曳時網口寬度8 m。每站位拖網時間為1 h, 拖網船速為3 kn。
1.2 樣品分析方法
樣品分析按照《海洋調查規范(GB/T 12763.6—2007)》的要求實施, 調查均在白天進行, 調查人員記錄每個站位的漁獲量, 漁獲量少于20 kg的站位, 全部樣品均留樣分析; 漁獲物20~40 kg的站位, 選取大型個體和稀有物種后, 剩余部分隨機抽取1/2樣品留樣; 漁獲量超過40 kg的站位, 選取大型個體和稀有物種后, 剩余部分隨機抽取20 kg樣品留樣。樣品冷凍保存后, 帶回實驗室進行分類鑒定和生物學測定。2010年冬季航次中SL16站未取得有效數據, 計算中未使用該航次此站數據。將119°30′E以西、以東海域分別定義為萊州灣西側海域、東側海域。
環境因子測定均按照《海洋調查規范》(GB/T 12763—2007)、《海洋監測規范》(GB 17378—2007)規定的方法進行。水溫、鹽度、氨、硝酸鹽、亞硝酸鹽、磷酸鹽的分析, 分別采用水溫表法、鹽度計法、鋅-鎘還原法、萘乙二胺分光光度法、次溴酸鹽氧化法、磷鉬藍分光光度法, 均為表、底層海水平均值; 葉綠素為表層海水數據。浮游植物使用淺水III型浮游生物網采集, 浮游動物使用淺水I型浮游生物網采集, 底棲動物使用抓斗采泥器(取樣面積0.05 m2, 每站取樣5次)采集。
1.3 數據處理
(1) 數據標準化
根據實際拖網時間和物種的可捕系數, 對調查數據進行標準化。中上層魚類可捕系數為0.3, 中底層魚類取0.5, 底層魚類取0.8[11]。
(2) 群落結構的相似性
通過Bray–Curtis相似性指數計算各航次物種組成的差異, 分析萊州灣游泳動物隨時間的變化規律。計算公式[12]為:
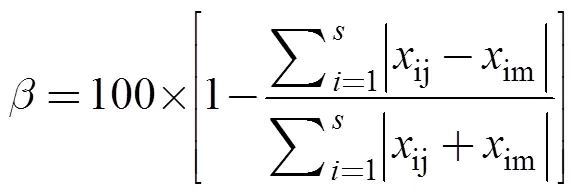
式中ij,im分別為第個種類在第個月份和第個月份的單位時間的生物量。
(3) 聚類與非度量多維標度分析
根據全年調查數據計算各種類生態優勢度(IRI), 僅保留IRI≥10的種類用于數據分析。2010年有29種(占生物量的99.7%), 2011年有24種(占生物量的99.3%), 合計共31種。為平衡群落中不同物種的作用, 生物量的數值進行平方根處理, 建立種類×站位矩陣, 利用PRIMER 6軟件非加權組平均法進行等級聚類和非度量多維標度(NMDS), NMDS的優劣使用協強系數判定, 協強系數在(0, 0.01)之間, 結果完全可信; 在(0.01, 0.05)之間, 結果可信; (0.05, 0.10)之間, 基本可信; (0.10, 0.20)之間, 結果有一定參考價值, 可用二維圖表示; >0.20, 結果幾乎是任意的, 不能在二維圖上展示[13]。
采用相似性百分比分析(SIMPER)探究不同物種對群落相異性和相似性的貢獻率。采用相似性分析檢驗(ANOSIM)進行顯著性檢驗,=0, 表示完全相同; 在(0, 0.5)之間, 表示差異不顯著; 在(0.5, 0.75)之間, 表示雖有重疊, 但可以清楚分開; 在(0.75, 1)之間, 表示差異顯著;=1, 表示完全不同。顯著性水平為<0.05[14]。
(4) DCCA
使用DCCA 排序來解釋物種與環境間的內在聯系, 揭示游泳動物對不同環境因子(生物的和非生物的)的適應性特征[15]。
(5) 數據處理
不同站點間生物量的比較采用單因素方差分析(one-way ANOVA), 并作雙尾t檢驗, 以<0.05為顯著相關。
2 結果與分析
2.1 種類組成
本研究8航次調查共捕獲游泳動物85種, 魚類55種, 屬于8目29科49屬; 甲殼類25種, 屬于2目22科25屬; 頭足類5種, 屬于3目3科3屬, 見表1。
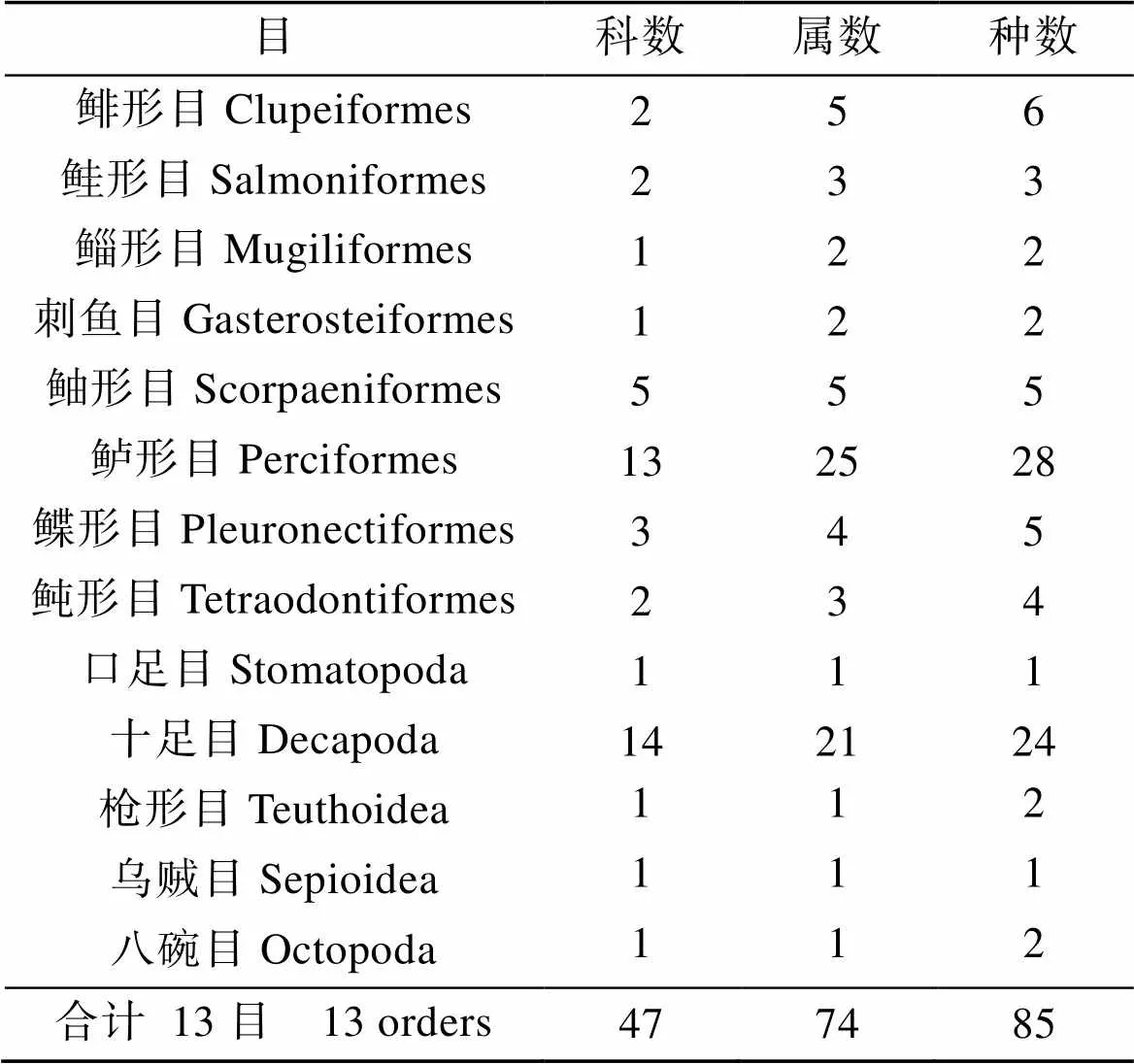
表1 2010—2011年萊州灣調查游泳動物組成
2.2 生物量及組成的季節變化
2011年調查海域相對生物量(52.3 kg/h)高于2010年(41.3 kg/h), 但年間差異不顯著(= –1.186,>0.05)。生物量季節間差別較大, 根據相同季節2年平均值計算, 各季節生物量排序為夏季>秋季>冬季>春季, 生物量均值季節差異顯著(ANOVA,<0.001)。夏季生物量與相同年份其他3季生物量差異均顯著, 春季與冬季生物量差異不顯著, 2010年秋季與春、冬季差異不顯著, 而2011年秋季與春、冬季差異則為顯著。相同季節生物量雖有差別, 春、夏、秋季生物量的年間差異均不顯著, 僅秋季存在顯著差異(圖2a)。
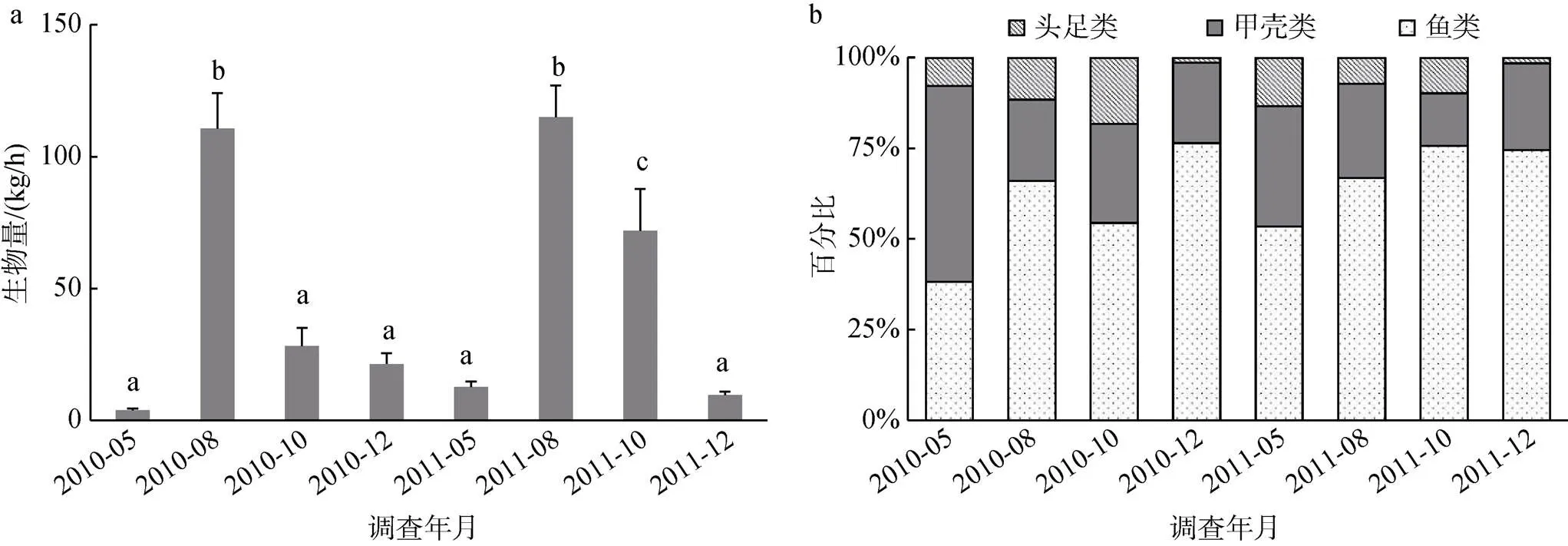
圖2 萊州灣游泳動物相對生物量(kg/h)的均值(+標準誤差)。
注: a: 相對生物量(kg/h); b: 生物量百分比(%); 相同小寫字母代表處理間差異不顯著(>0.05)
除2010年春季甲殼類生物量高于魚類, 其他季節均為魚類生物量最高(生物量百分比均超過50%), 甲殼類次之, 頭足類最低。生物量季節間的變化主要是由于魚類生物量的季節變化導致的, 冬季魚類生物量比例達到全年最高, 而春季最低, 甲殼類和頭足類生物量的季節變化幅度較小(圖2b)。
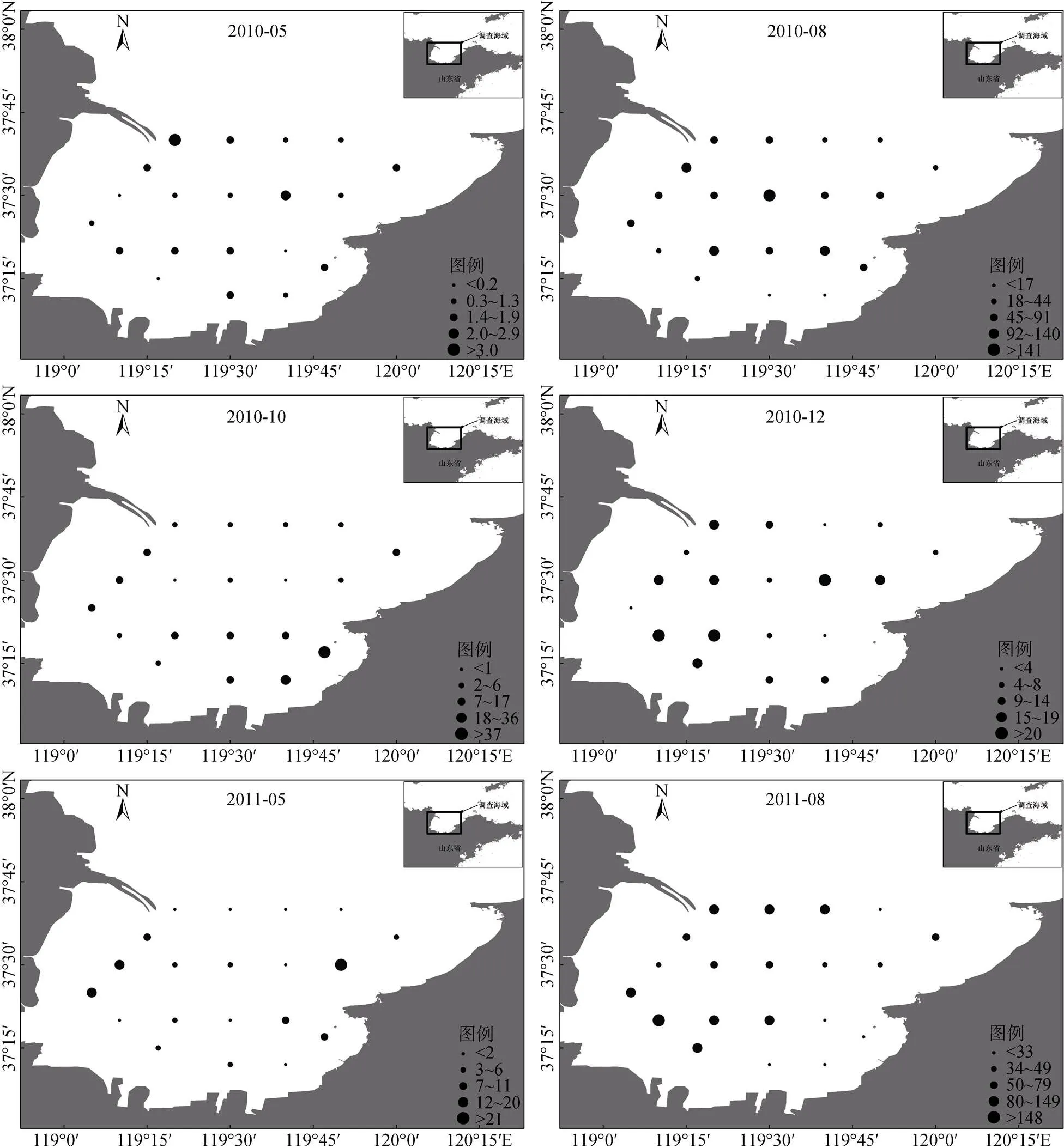
2.3 生物量空間分布
從游泳動物生物量空間分布可以看出(圖3), 在多數季節灣底水域生物量偏低, 而黃河口南側海域生物量較高, 存在一定的異質性。從全年調查平均值來看, 萊州灣西側游泳動物生物量(60.2 kg/h)明顯高于東側(33.4 kg/h;=4.597,<0.001)。
2.4 群落劃分
根據8航次31個主要種類生物量聚類, 萊州灣20調查站位可劃分為4組, 協強系數(2D stress=0.14), 表明所有NMDS排序結果在二維空間上是可信的。組I由位于調查海域東南的3個站位組成, 組II位于調查海域東部及中部的8個站位組成, 組III由調查海域西北部(黃河口南側)的3個站位組成, 組IV由西部6個站位組成(圖4)。組I浮游植物豐度、浮游動物生物量、游泳動物生物量、游泳動物多樣性指數在各組中均最低, 而底棲動物生物量最高; 組III和組IV的浮游植物豐度、浮游動物生物量、游泳動物生物量高于組I和組II(表2)。鮻()、石鰈()在組I中比例明顯高于其他3組; 短蛸()、日本褐蝦()、黃鰭刺蝦虎魚()在組II中比例明顯高于其他3組; 短吻紅舌鰨()、方氏錦鳚()、葛氏長臂蝦()、脊尾白蝦()、青鱗小沙丁魚()、日本鼓蝦()在組III的比例明顯高于其他3組; 斑鰶(、赤鼻棱鳀()、黃鯽()、脊尾白蝦、口蝦蛄()、皮氏叫姑魚()、中國明對蝦()在組IV中比例明顯高于其他三組。斑尾刺蝦虎魚()、槍烏賊(sp.)、鲬()、日本蟳()、疣背深額蝦()在各組中比例較為接近。
ANOSIM分析表明, 所有組間值均小于0.5, 顯著水平也多在可接受范圍(<0.5%, 除組III&組IV外), 表明組間群落結構雖有不同, 但差異并不顯著(表3)。SIMPER分析表明, 斑尾刺蝦虎魚、槍烏賊和日本蟳對4個組的組內相似性貢獻均較高; 矛尾蝦虎魚()、口蝦蛄和短吻紅舌鰨對組II、組III和組IV組內相似性貢獻較高; 斑鰶對組I、組III和組IV的組間相似性貢獻較高。日本褐蝦對組I相似性較高, 而葛氏長臂蝦、脊尾白蝦和青鱗小沙丁魚對組IV相似性貢獻較高(表4)。
2.5 調查站位和環境因子DCCA排序
圖5為各調查站位游泳動物生物量(圓圈)與環境因子DCCA排序圖。溫度、鹽度、底棲動物生物量以及浮游植物豐度是影響群落的重要因子, 無機磷對群落影響較小。由表5可以看出, 與環境第一排序軸相關系數較高的是溫度(–0.939 8)和鹽度(–0.321 5), 與環境第二排序軸密切相關的是浮游植物豐度(0.477 8)和底棲動物生物量(–0.406 4)。
3 討論
3.1 生物量及組成的變化
群落總生物量(Total biomass)的變化是反映群落功能變化的一個最簡單、最直接的指標[16]。與歷史調查結果相比, 目前萊州灣游泳動物生物量發生了明顯變化。
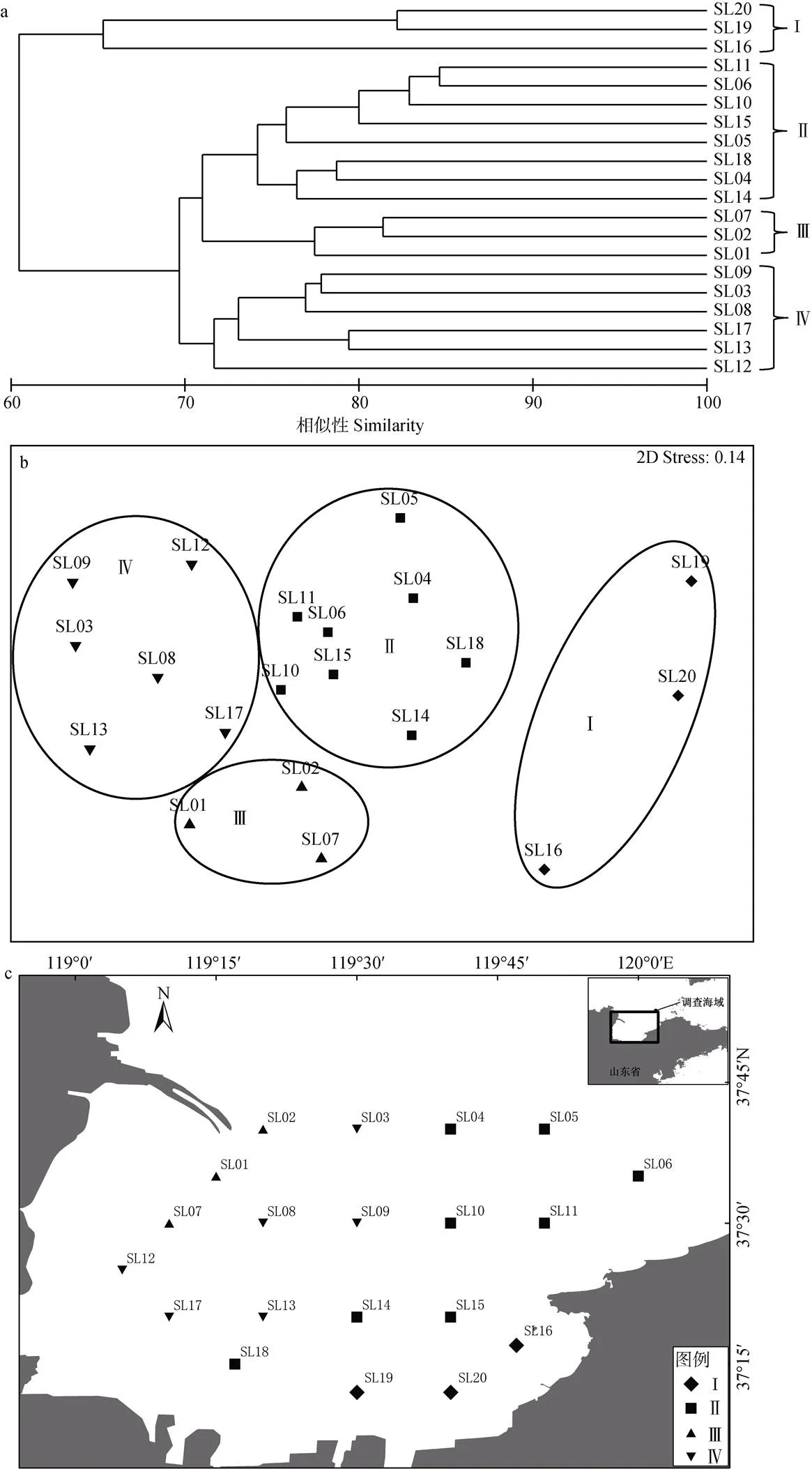
圖4 萊州灣游泳動物調查站位聚類圖(a)、NMDS排序圖(b)和空間分布圖(c)
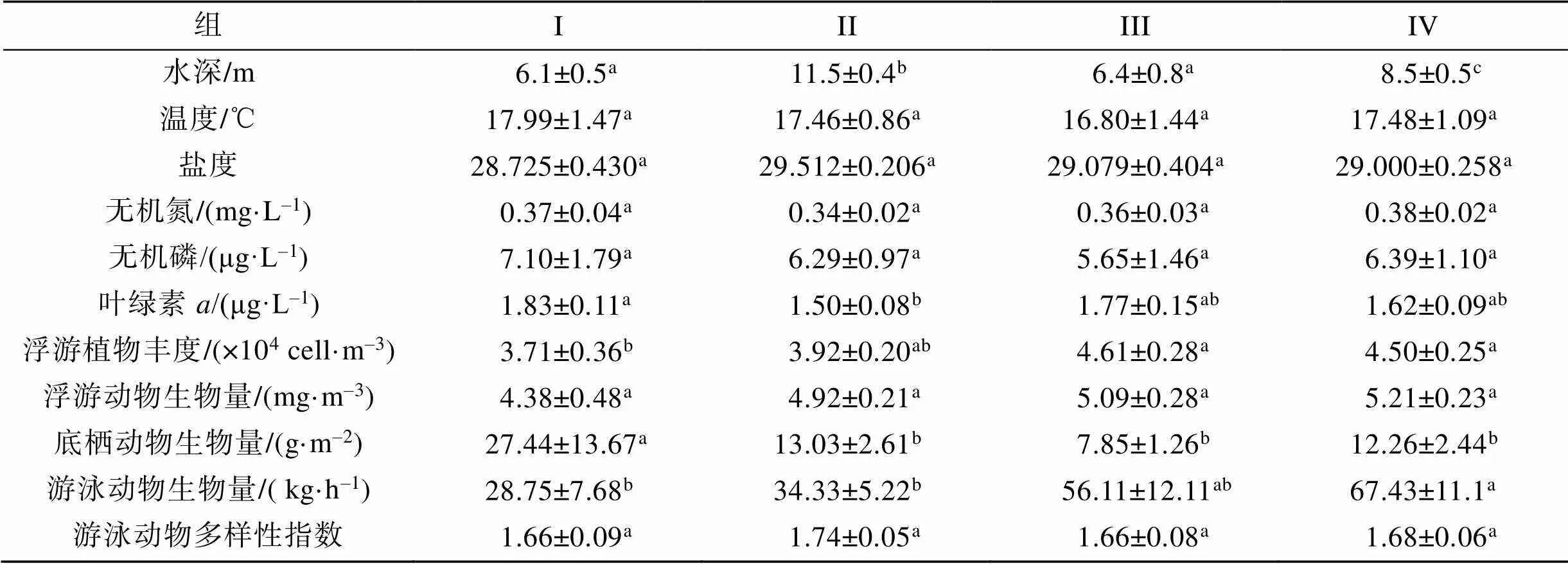
表2 萊州灣調查站位組間差別
注: 同一行比較, 相同字母為差異不顯著, 有不同字母為差異顯著(<0.05)
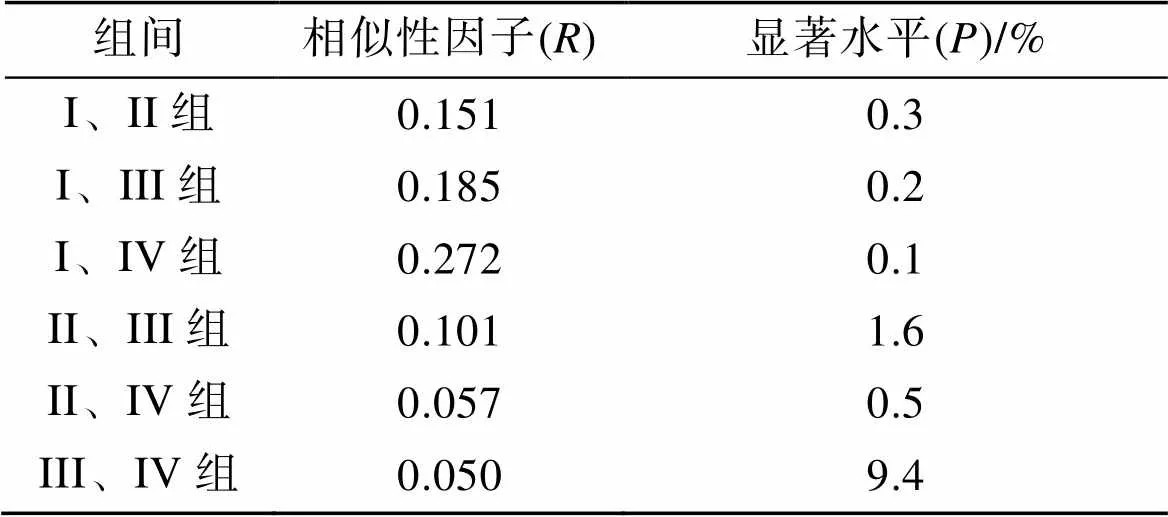
表3 萊州灣游泳動物群落單因子相似性分析
總生物量整體上處于較低水平, 其中只有夏季能達到2 t/km2, 其余季節均小于1 t/km2; 春季生物量下降尤為嚴重, 僅0.2 t/km2左右。相關研究表明, 渤海春季魚類生物量由1959年的186 kg/h降到了2010年的不足1 kg/h[8]。春季資源狀況不佳, 也導致春季幾乎沒有漁船在萊州灣生產。萊州灣游泳動物生物量自1959至1998年間的4次調查呈逐次下降趨勢, 特別是春季生物量下降幅度大[2]。生物量的急劇下降與主要經濟物種資源衰退有直接關系, 渤海海域主要漁業資源表現出小型化和低齡化的發展趨勢[9]。張波等[17]研究發現, 餌料生物優勢種的演替也是導致漁業資源生物量和種類變動的重要原因, 還導致其生態位發生變化。另外, 海域環境惡化引起魚類繁殖能力、幼體存活率和種群增長率的急劇下降, 嚴重影響了漁業資源的補充量, 進而導致生物量的降低。
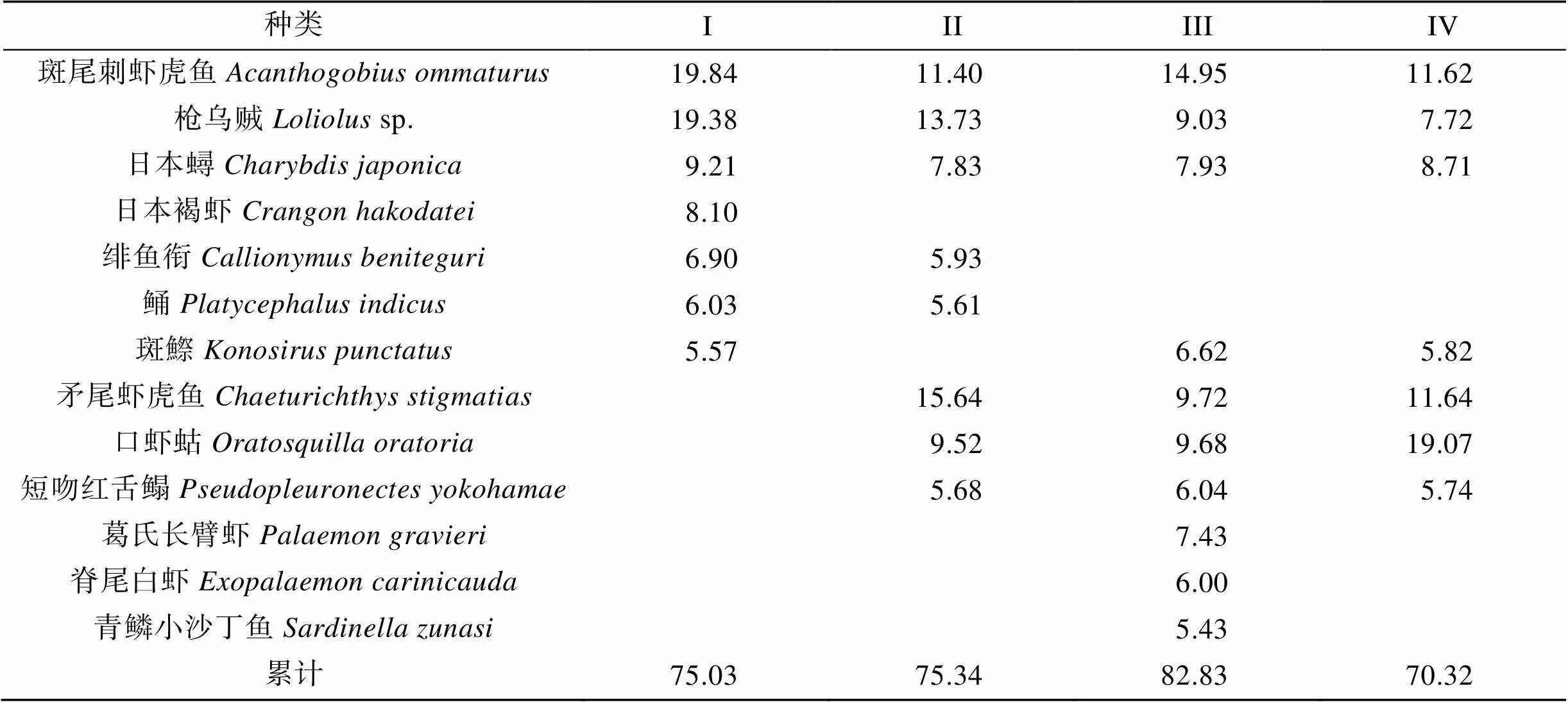
表4 根據SIMPER分析得到的群落類型中對平均相似性的貢獻率>5%的主要特征種類

圖5 萊州灣環境因子與游泳動物生物量的DCCA排序
注: Depth: 水深; Temp: 溫度; Sal: 鹽度; Din: 無機氮濃度; Dip: 無機磷濃度; Chl: 葉綠素濃度; Phyto: 浮游植物豐度; Zoo: 浮游動物生物量; Benth: 底棲動物生物量; ○為調查站位游泳動物生物量
與1959—1998年調查結果[2, 5]對比后發現, 目前萊州灣游泳動物生物量低于1992年之前的歷次調查, 但高于1998年調查值。相關研究表明, 2003年、2006年、2008年萊州灣春季漁業資源均高于1998年春季[9], 21世紀后, 萊州灣游泳動物資源較20世紀90年代末期有所恢復。這可能與自上世紀80年代末陸續實施的多項漁業資源管理措施有一定關系, 如1988年拖網漁業退出渤海, 1995年開始的伏季休漁, 以及藍點馬鮫、中國明對蝦、海蜇()等單種漁業資源專項管理措施等, 同時自2005年山東省開始實施漁業資源修復行動計劃以來, 對資源修復起到了一定作用。但目前生物量主要依靠幼魚和低營養級種類(相同的初級生產力可以支撐更高的生物量)支撐, 資源仍遠未達到完全修復的程度。相關研究[8]也表明增殖放流種類資源量遠低于歷史最高資源量, 資源恢復尚有很大空間。
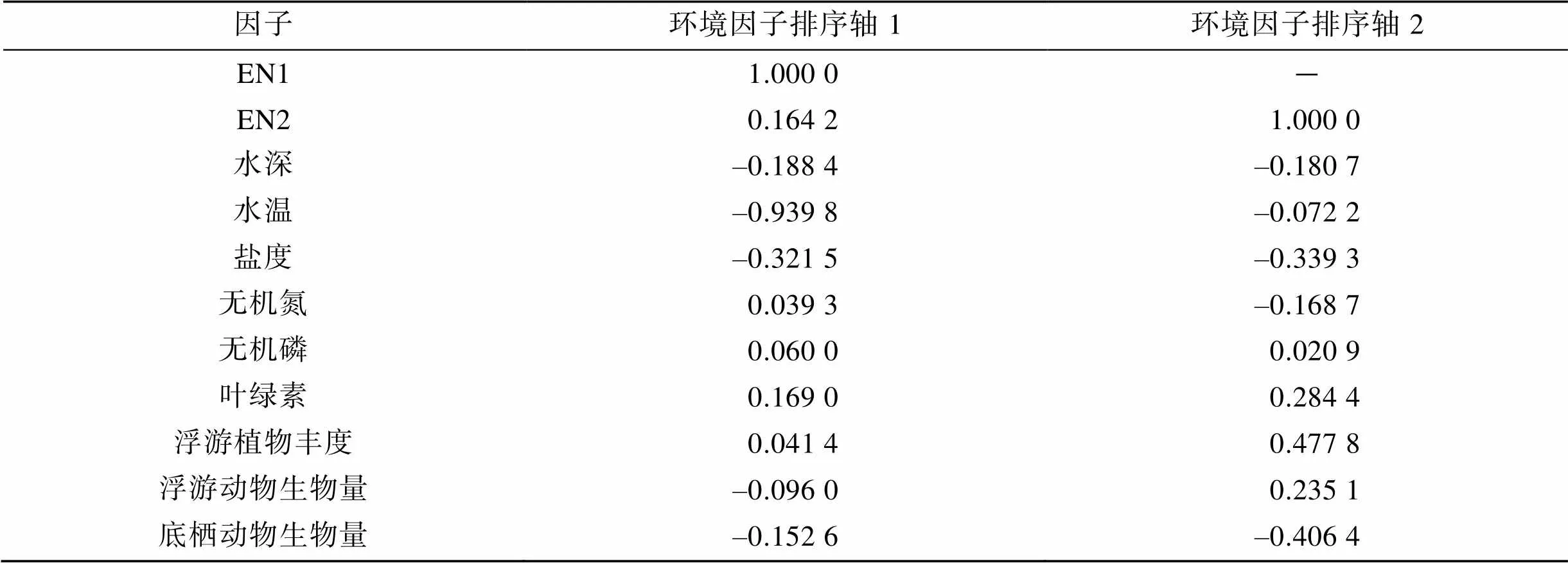
表5 環境因子與前兩個排序軸的相關系數
本次調查結果顯示, 萊州灣游泳動物生物量的季節分布發生了較大程度的改變, 夏季最高, 春季最低。1959年春季生物量明顯高于夏、秋季, 春季生物量主要由帶魚()、小黃魚()的產卵群體貢獻[2], 而目前春季帶魚、小黃魚生物量比例幾乎可以忽略(調查未捕獲帶魚, 小黃魚在2011年春季航次中出現, 占生物量比例的0.42%), 而這一時期萊州灣主要游泳動物如斑鰶和青鱗小沙丁魚等尚未大量進入萊州灣, 同期多數斑尾刺蝦虎魚個體在繁殖后已經死亡, 造成春季生物量較低。夏季是目前一年中生物量最高的季節, 這要由當年生幼魚貢獻(得益于伏季休漁對當年生幼魚的保護)。目前秋季生物量明顯低于夏季, 可能與休漁結束后大量個體被捕撈, 以及部分種類洄游離開有關。萊州灣漁業賴以成名的春汛、秋汛早已消失, 目前萊州灣漁業生產主要集中在夏季、秋季, 目標種類主要為增殖放流資源種類——中國明對蝦、三疣梭子蟹()和海蜇, 其它種類基本不能形成專捕漁業(未發表數據)。
底層魚類生物量比例下降趨勢有所逆轉, 小型無脊椎動物比例升高。自1959年至1998年, 萊州灣中上層魚類比例呈不斷上升趨勢, 底層魚類比例則不斷下降[2]。而目前中上層魚類比例較1998年有所下降, 底層魚類比例有所上升, 各類別生物量比例與1992年較為接近。與歷史調查值相比, 目前頭足類比例處于較高水平, 槍烏賊(火槍烏賊和日本槍烏賊)在某些季節甚至成為優勢種。2011和2011年各類別生物量比例有所差異, 但主要是青鱗小沙丁魚在2年間生物量的巨大差異所導致的。
3.2 萊州灣游泳動物群落異質性
游泳動物群落時空分布的異質性與海水環境、餌料生物、洄游、海底地貌等因素均密切相關[10, 18-19]。本文DCCA分析表明, 溫度是影響萊州灣游泳動物時空分布的最主要因素, 這主要是由于萊州灣游泳動物種類主要以暖溫種為主, 隨著季節變化, 種類和生物量因洄游魚類進出而發生較大改變; 底棲動物生物量也是影響游泳動物群落的重要因子之一, 張波等的研究也證實底棲動物在目前萊州灣生態系統的食物關系中起關鍵作用[17]; 而水深并不是影響萊州灣游泳動物群落的主要環境因子, 這與萊州灣水深較淺、水深梯度變化較小(最深水深僅18 m, 大部分水深在10 m以內)有關。
根據主要種類生物量聚類, 萊州灣游泳動物群落可劃分為4個組。組I位于萊州灣東南部, 底質以粉砂質砂為主, 處于扇貝筏式養殖區和單環刺螠漁業作業區邊緣, 與其他組相比, 組I種類數、生物量以及Shannon-Wiener多樣性明顯偏低, 李少文等的研究表明該區域底棲生物種類數和多樣性也明顯偏低, 而次級生產力明顯偏高, 表明游泳動物、底棲生物群落受到了明顯干擾[20]。組II的8個站位主要位于灣東北部(龍口周邊海域), 該組站位底質以礦質粉砂為主且復雜多變, 水深較深, 海水環境質量較好(2010年、2011年山東省海洋環境公報), 組II是游泳動物多樣性最高的組, 黑棘鯛()、許氏平鲉()和大瀧六線魚()等戀礁魚類以及喜穴居的蛸類(長蛸、短蛸)等對生境要求高的種類在主要出現于該組。組III的3個站位位于離黃河口最近的區域, 脊尾白蝦、青鱗小沙丁魚偏好在此聚集, 底棲動物生物量較低, 底棲動物食性的石鰈、緋魚銜()生物量明顯低于其他組; 組IV由靠近小清河口的2個站位和萊州灣中央的4個站位組成, 底質以粉砂質黏土為主, 海水富營養化最嚴重(2010年、2011年山東省海洋環境公報), 在各組中游泳動物生物量最高, 但多樣性偏低, 該組浮游動物生物量較高, 斑鰶、赤鼻棱鳀等浮游動物食性魚類生物量高。
ANOSIM分析表明聚類各組之間雖有差異, 但差異不顯著(表4), 總體而言, 萊州灣游泳動物群落的時空分布異質性處于較低水平。這與黃海山東海域(魚類群落可明顯區分為近岸淺水區和遠岸深水區2個不同的組[6])和北部灣(魚類群落各季節均可劃分為5個較為穩定的組[10])明顯不同。萊州灣游泳動物群落異質性較低, 可能與萊州灣自然環境、氣候變化和人為影響密切相關。
1) 與黃海山東海域和北部灣等海域相比, 萊州灣面積較小(僅6 215.4 km2), 水深變化梯度小; 作為半封閉海灣-渤海的一部分, 萊州灣水交換周期長, 約為0.5年[21]。
2) 作為影響萊州灣最大的河流—黃河, 受上游大型水利工程建設以及沿岸工農業用水的影響, 入海徑流量不斷減少, 并發生數次斷流, 雖然自2002年實施調水調沙以來, 黃河斷流已不復存在, 但徑流量仍較小, 這也造成萊州灣淡水水體面積較小[22], 整個灣內鹽度較高且鹽度梯度較小。渾濁度高、營養豐富的低鹽水體生境, 不僅可為漁業資源早期生活史階段提供足夠的食物, 同時也是肉食性魚類的天然屏障, 為幼魚良好的育幼場和庇護場。低鹽水體的減少和大型水利工程對洄游通道的破壞, 加劇了生境的單一程度, 導致河口種和河道洄游種基本絕跡, 降低了游泳動物群落格局異質性。
3) 萊州灣是傳統捕撈作業區, 捕撈壓力大, 過度捕撈導致群落特有種滅絕,-選擇型種類成為優勢種, 這些種類在單一生境下廣為分布。同時, 對底棲環境破壞嚴重的網具底拖網、弓子網等被大量使用, 加劇了底質荒漠化和生境同質化程度。近岸灘涂、淺海養殖以及圍填海、旅游開發、海岸工程對海岸帶的侵占, 導致巖礁、海草場等特有生境消失。而海水富營養加劇, 氮磷比不斷升高, 改變了浮游植物群落結構, 食物鏈朝向不利于漁業資源種類、有利于大型水母的方向發展[23], 大型水母暴發會大量攝食魚卵仔魚, 選擇壓力, 加劇了-選擇種的生存壓力, 進一步降低游泳動物群落異質性。
4 結論
綜上, 自然環境特點、氣候變化和人為影響導致萊州灣生境異質性程度較低, 對生境要求較高的種類(如河道洄游種、河口種、軟骨魚類、巖礁魚類等)不利, 從而造就了目前萊州灣較低的游泳動物群落異質性。根據生態學基本原理, 豐富的小生境將增加群落格局異質性和生物多樣性, 合理發展海洋牧場, 包括集魚型魚礁、海草場和海藻場, 可提高生境異質性程度, 對生物多樣性保護和資源修復具有積極意義。
[1] Jin X S, Tang Q S. Changes in fish species diversity and dominant species composition in the Yellow Sea[J]. Fisheries Research, 1996, 26(3-4): 337-352.
[2] 金顯仕, 鄧景耀. 萊州灣漁業資源群落結構和生物多樣性變化[J]. 生物多樣性, 2000, 8(1): 65-72. Jin Xianshi, Deng Jingyao. Variations in community structure of fishery resources and biodiversity in the Laizhou Bay, Shandong[J]. Biodiversity Science, 2000, 8(1): 65-72.
[3] 山東省海洋與漁業廳. 山東省海洋環境公報[R]. 濟南: 山東省海洋與漁業廳, 2010. Shandong Provincial Department of Oceans and Fisheries. Marine Conditions of Shandong[R]. Jinan: Shan-dong Provincial Department of Oceans and Fisheries, 2010.
[4] 金顯仕, 鄧景耀. 萊州灣春季漁業資源及生物多樣性的年間變化[J]. 海洋水產研究, 1999, 20(1): 6-12. Jin Xianshi, Deng Jingyao. Yearly variations of fishery resources and biodiversity in the Laizhou Bay[J]. Progress in Fishery Sciences, 1999, 20(1): 6-12.
[5] 鄧景耀, 金顯仕. 萊州灣及黃河口水域漁業生物多樣性及其保護研究[J]. 動物性研究, 2000, 21(1): 76-82. Deng Jingyao, Jin Xianshi. Study on fishery biodiversity and its conservation in Laizhou Bay and Yellow River estuary[J]. Zoological Research, 2000, 21(1): 76-82.
[6] 王平, 焦燕, 任一平, 等. 萊州灣、黃河口水域春季近岸漁獲生物多樣性特征的調查研究[J]. 海洋湖沼通報, 1999, (1): 40-44. Wang Ping, Jiao Yan, Ren Yiping, et al. Investigation of biodiversity from spring catch in coastal waters of Laizhou Bay and Huanghe estuary[J]. Transactions of Oceanology and Limnology, 1999, (1): 40-44.
[7] 陳大剛, 沈謂銓, 劉群, 等. 萊州灣及黃河口水域地理學特征與魚類多樣性[J]. 中國水產科學, 2000, 7(3): 46-52. Chen Dagang, Shen Weiquan, Liu Qun, et al. The geographical characteristics and fish species diversity in the Laizhou Bay and Yellow River estuary[J]. Journal of fishery sciences of China, 2000, 7(3): 46-52.
[8] 單秀娟, 金顯仕, 李忠義, 等. 渤海魚類群落結構及其主要增殖放流魚類的資源量變化[J]. 漁業科學進展, 2012, 33(6): 1-9. Shan Xiujuan, Jin Xianshi, Li Zhongyi, et al. Fish com-munity structure and stockdynamics of main releasing fish species in the Bohai Sea[J]. Progress in Fishery Sciences, 2012, 33(6): 1-9.
[9] Jin X S, Shan X J, Li X S, et al. Long-term changes in the fishery ecosystem structure of Laizhou Bay, China[J]. Science China Earth Sciences, 2013, 56: 366-374.
[10] Li F, Zhang H J, Lü Z B, et al. Species composition and community diversity of nekton in Laizhou Bay, China[J]. Biodiversity Science, 2013, 21: 537-546.
[11] 詹秉義. 漁業資源評估[M]. 北京: 中國農業出版社, 2000. Zhan Bingyi. Fisheries Resources Assessment[M]. Beijing: Agriculture Press of China, 2000.
[12] Bray T R, Curtis J T. An ordination of the upland forest communities of southern Wisconsin[J]. Ecological Monographs, 1957, 27(4): 325-349.
[13] 張濤, 莊平, 章龍珍, 等. 長江口中華鱘自然保護區底層魚類的群落結構特征[J]. 生態學報, 2011, 31(6): 1687-1694. Zhang Tao, Zhuang Ping, Zhang Longzhen, et al. Community structure of demersal fish in Nature Reserve of Acipenser sinensis in Yangtze River estuary[J]. Acta Ecologica Sinica, 2011, 31(6): 1687-1694.
[14] Clarke K R, Warwick R M. A further biodiversity index applicable to species lists: variation in taxonomic distinctness[J]. Marine Ecology Progress Series, 2001, 216: 265-278.
[15] 張金屯. 數量生態學(第二版)[M]. 北京: 科學出版社, 2011. Zhang Jintun. Quantitative Ecology (Second Edition)[M]. Beijing: Science Press, 2011.
[16] 程家驊, 姜亞洲. 捕撈對海洋魚類群落影響的研究進展[J]. 中國水產科學, 15(2): 359-366. Cheng Jiahua, Jiang Yazhou. Methods for evaluating fishing effects on fish community[J]. Journal of Fishery Sciences of China, 15(2): 359-366.
[17] 張波, 吳強, 金顯仕. 萊州灣魚類群落的營養結構及其變化[J]. 漁業科學進展, 2013, 34(2): 1-9. Zhang Bo, Wu Qiang, Jin Xianshi. Feeding ecology of fish assemblages and its variations in the Laizhou Bay[J]. Progress in Fishery Sciences, 2013, 34(2): 1-9.
[18] 李圣法, 程家驊, 嚴利平. 東海大陸架魚類群落的空間結構[J]. 生態學報, 2007, 27(11): 4377-4386. Li Shengfa, Cheng Jiahua, Yan Liping. Spatial struc-tures of fish communities on the continental shelf of the East China Sea[J]. Acta Ecologica Sinica, 2007, 27(11): 4377-4386.
[19] 任一平, 徐賓鐸, 葉振江, 等. 青島近海春、秋季漁業資源群落結構特征的初步研究[J]. 中國海洋大學學報(自然科學版), 2005, 35(6): 792-798. Ren Yiping, Xu Binduo, Ye Zhenjiang, et al. Preliminary study on community structure of fishery resources during spring and autumn in the coastal waters of Qingdao[J]. Periodical of Ocean University of China, 2005, 35(6): 792-798.
[20] 李少文, 劉元進, 李凡, 等. 萊州灣大型底棲動物功能群現狀[J]. 生態學雜志, 2013, 32(2): 380-388. Li Shaowen, Liu Yuanjin, Li Fan, et al. Macrobenthic functional groups in Laizhou Bay, East China[J]. Chinese Journal of Ecology, 2013, 32(2): 380-388.
[21] 魏皓, 田恬, 周鋒, 等. 渤海水交換的數值研究–水質模型對半交換時間的模擬[J]. 青島海洋大學學報 (自然科學版), 2002, 32(4): 519-525. Wei Hao, Tian Tian, Zhou Feng, et al. Numerical study on the water exchange of the Bohai Sea: simulation of the half -life time by dispersion model[J]. Periodical of Ocean University of Qingdao, 2002, 32(4): 519-525.
[22] 肖純超, 張龍軍, 楊建強. 2004—2009年黃河口近岸海域低鹽區面積的變化趨勢研究[J]. 中國海洋大學學報 (自然科學版), 2012, 42(6): 40-41. Xiao Chunchao, Zhang Longjun, Yang Jianqiang. Study on the change of low-salinity area in the Yellow River Estuary and its coastal zone from 2004 to 2009[J]. Periodical of Ocean University of China, 2012, 42 (6): 40-41.
[23] 許思思, 宋金明, 李學剛, 等. 漁業捕撈對渤海漁業資源及生態系統影響的模型研究[J]. 資源科學, 2011, 33(6): 1153-1162. Xu Sisi, Song Jinming, Li Xuegang, et al. Modeling of fishing effects on fishery resources and ecosystems of the Bohai Sea[J]. Resources Science, 2011, 33(6): 1153-1162.
Community structure of nektons in Laizhou Bay, China
ZHANG Huan-jun1, LIU Shu-de2, LIU Yuan-jin1, TU Zhong2, ZHOU Quan-li1, CHEN Wei1, SUN Wei1, LI Fan1
(1. Shandong Marine Resources and Ecology Research Institute, Shandong Provincial Key Laboratory of Restoration for Marine Ecology, Yantai 264006, China; 2. Shandong Hydrobios Resources Conservation and Management Center, Yantai 264003, China)
Based on the data of 8 cruises in spring (May), summer (August), Autumn(October) and winter (December) in 2010 and 2011, the community pattern of nekton and its relationship with environmental factors in Laizhou Bay were studied. Overall, a total of 85 species of nekton were caught, the values of nekton biomass were higher in 2011 than in 2010. The biomass had a great difference between seasons, and the order of the biomass was summer, autumn, winter and spring. The biomass of the same season had no significant difference in different years except autumn. In all surveys except in the spring of 2010, the biomass of fishes was highest, followed by that of crustaceans and cephalopods. Nekton biomass was significantly higher in the western half of the bay (60.2 kg/h) than in the eastern half (33.4 kg/h). We grouped the biomass values of 31 main species of nekton captured at the 20 stations by cluster analysis: group I comprised the southeastern areas (three stations); group II comprised the eastern and middle areas (eight stations); group III comprised the northwestern areas (three stations); and group IV comprised the western areas (six stations). Group I had the lowest phytoplankton abundance, zooplankton biomass, and nekton biomass and diversity, but had the highest macrobenthos biomass. Phytoplankton abundance, zooplankton biomass, and nekton biomass were higher in groups III and IV than in groups I and II. The analysis of similarities revealed different community structures among the four groups; however, there were no significant differences. Detrended canonical correspondence analysis showed that temperature, salinity, benthic biomass, and phytoplankton abundance are the significant factors affecting nekton communities. The nekton community of Laizhou Bay has changed remarkably as compared with its previous survey values: total nekton biomass was at relatively low levels, especially in spring; seasonal distributions of biomass had become even lower in spring and higher in summer; and a downward trend in the proportion of demersal fishes had somewhat reversed. The nekton community now appears more homogeneous, possibly because of an increasingly homogeneous environment and the severe effects of human activities.
biomass; cluster analysis; NMDS; DCCA
Dec. 6, 2019
S931
A
1000-3096(2020)06-0110-12
10.11759/hykx20191206001
2019-12-06;
2020-03-05
山東省漁業資源增殖效果評價項目(SD-XGPJ-2012-2); 山東省重點研發計劃(2018GHY115014)
[Evaluation on the effect for fishery enhancement and releasing of Shandong Province, No. SD-XGPJ-2012-2; Key Research and Development Program of Shandong Province, No. 2018GH115014]
張煥君(1979-), 男, 漢族, 山東威海人, 碩士研究生, 副研究員, 主要從事漁業資源與生態研究, 電話: 18153518135, E-mail: hjzhang815@163.com;李凡,通信作者, 電話: 18153518186, E-mail: lifan811230@126.com
(本文編輯: 楊 悅)
