大興安嶺北部針葉林土壤團聚體酶活性研究
2020-07-14 08:51:08朱家琪滿秀玲張頔劉思琪
森林工程 2020年4期
朱家琪 滿秀玲 張頔 劉思琪
摘 要:以大興安嶺北部典型針葉林(興安落葉松林、樟子松林)為研究對象,通過團聚體脲酶、蔗糖酶、酸性磷酸酶活性以及酶活性幾何平均數的測定與分析,探明該地區兩種主要針葉林土壤團聚體酶活性的差異,以及隨團聚體粒徑大小不同酶活性的變化特征。結果表明:①興安落葉松林0~5 cm土層和樟子松林0~10 cm土層以大團聚體占優勢,隨著土層加深大團聚體含量下降,兩種林型生長季中期大團聚體含量高于生長季初期和末期。②兩種林型團聚體酶活性均表現為隨粒徑減小而增加的一致性規律,以小于0.25 mm微團聚體酶活性最高,且隨著土層加深,團聚體酶活性隨之降低。興安落葉松林團聚體脲酶、蔗糖酶活性低于樟子松林,酸性磷酸酶活性高于樟子松林;兩種林型團聚體脲酶以生長季初期和末期活性最高,蔗糖酶和酸性磷酸酶均以7—8月生長旺季活性最高。③土壤團聚體酶活性幾何平均數表現為樟子松林高于興安落葉松林,隨土層降低而遞減,且隨粒徑增大酶活性平均數降低,兩種林型酶活性幾何平均數在生長季中期數值較高。
關鍵詞:大興安嶺;興安落葉松林;樟子松林;水穩性團聚體;酶活性
Abstract:Taking the typical coniferous forest (Larix gmelinii forest, Pinus sylvestris var. mongolica forest) in the north of Daxingan Mountains as the research object, the activity of aggregate urease, invertase, acid phosphatase and the geometric mean of enzyme activity were measured and analyzed, in order to find out the difference of enzyme activity between the two main coniferous forests in this area, and the change characteristics of enzyme activity with the particle size of aggregates. The results showed that: ①Large aggregates dominated the 0-5 cm soil layer of Larix gmelinii forest and 0~10 cm soil layer of Pinus sylvestris var. mongolica forest. As the soil deepened, the content of large aggregates decreased, and the content of large aggregates in the middle growth season of the two forest types was higher than that in the early and late growth season. ②The enzyme activity of aggregates of the two types of forest showed a consistent rule of increasing with the decrease of particle size, with the enzyme activity of <0.25 mm microaggregates being the highest, and the enzyme activity of aggregates decreased with the deepening of the soil layer. The activity of the aggregate urease and invertase in Larix gmelinii forest was slightly lower than that in Pinus sylvestris var. mongolica forest, and the activity of acid phosphatase was higher that in Pinus sylvestris var. mongolica forest. The aggregate urease of the two types of forest was the highest at the beginning and end of the growing season, while the activity of invertase and acid phosphatase was the highest in the peak growing season from July to August. ③The geometric mean of the enzyme activity of soil aggregates showed that the Pinus sylvestris var. mongolica forest was higher than that of Larix gmelinii forest, which decreased with the decrease of soil layer, and the mean of enzyme activity decreased with the increase of particle size. The geometric mean of enzyme activity of the two forest types was higher in the middle growing season.
Keywords:Daxingan Mountains; Larix gmelinii forest; Pinus sylvestris var. mongolica forest; soil water stable aggregate; enzyme activity
0 引言
土壤酶是微生物養分轉化過程中的催化物質,其中土壤脲酶能夠參與尿素的分解,其酶促反應能夠得到氨,是植物氮素營養的主要來源[1],蔗糖酶能夠以有機質為底物,將高分子的化合物分解成為植物可利用的營養物質,是參與土壤碳循環的重要酶,而磷酸酶能夠促進土壤有機磷的礦化,有助于植物對有機磷的吸收[2]。脲酶、蔗糖酶與磷酸酶直接參與土壤碳氮磷的重要循環過程,對于生境的變化十分敏感,因此可以作為土壤質量的生物指標[3]。研究證明,土壤酶受溫濕度、微生物活性等因素的綜合影響[4-5],而不同粒徑大小的土壤團聚體中通氣狀況與水分含量等因素不同,造成了團聚體中養分轉換及酶活性的差異,因此研究團聚體中酶的活性能夠更好地揭示土壤中養分轉換的生物過程。目前,在土壤團聚體酶活性的研究中,有許多研究者對團聚體脲酶、蔗糖酶和酸性磷酸酶進行了研究,對于植被恢復過程以及施肥對團聚體酶的作用研究較多[6-7],也有不少研究涉及有機碳含量對酶活性的影響[8-9],而森林生態系統下,土壤酶受到土壤、氣候等多種因素相互作用,共同調控土壤微生物的群落結構,從而對酶的活性及釋放造成影響[10]。因此,不同森林類型中團聚體酶活性差異的相關研究對深入了解森林生態系統養分循環具有積極的作用。大興安嶺北部林區是我國唯一的寒溫帶森林,也是我國主要的林業生產基地,地帶性植被為明亮針葉林,即興安落葉松(Larix gmelinii)林,同時也分布有樟子松(Pinus sylvestris var. mongolica)林,除此之外還有白樺(Betula platyphylla)林、山楊(Populus davidiana)林等次生林[11]。但大興安嶺地區土層較薄,生長季短,對于該地區團聚體內酶活性的實測數據還十分欠缺。本研究以大興安嶺北部地區天然針葉林為研究對象,探討該地區土壤團聚體內酶活性的差異,為本地區土壤酶活性的研究提供科學參考。
1 材料與研究方法
1.1 研究區概況
研究區設于黑龍江漠河森林生態系統國家定位觀測研究站,位于大興安嶺北部漠河縣北極村內,地理坐標為:122°06′~122°27′ E,53°17′~53°30′ N。本區屬寒溫帶大陸性季風氣候,年平均氣溫-4.9 ℃,年降水量約430 mm,降水多集中在夏季。全年冰雪覆蓋時間最長可達200 d。地帶性土壤為棕色針葉林土,另外局部地區還有草甸土、沼澤土分布,是我國多年凍土的主要分布區。森林植被系歐亞大陸寒溫帶明亮針葉林,以興安落葉松為優勢建群種,此外還有樟子松林、白樺林和山楊林等森林類型,林下灌木主要有興安杜鵑(Rhododendron dauricum)、杜香(Ledum palustre)和越桔(Vaccinium vitis-idaea)等。
1.2 樣地設置和研究方法
在前期踏查的基礎上,選擇本區典型針葉林為研究對象,即興安落葉松林和樟子松林,在每一林形設置3塊調查樣地,樣地規格為20 m×30 m,并對樣地進行基本情況調查,見表1和表2。2018年5—10月每月初取樣一次,在每一樣地每月按照“S”形設置采樣點,挖掘土壤剖面,利用土刀分別采集0~5、5~10 、10~20 cm 3個土層的土壤樣品,剔除石塊及動植物殘體,室溫風干后,采用濕篩法篩分出:>1.0、0.5~1.0、0.25~0.5、0.053~0.25、<0.053 mm 5個粒徑的水穩性團聚體。在取土的同時,用環刀法采集原狀土樣以測定土壤容重與孔隙度。
土壤團聚體的測定方法:利用TTF-100型土壤團聚體分析儀,稱取50 g風干土樣,置于1 mm篩子上,使用蒸餾水浸潤5 min,以90次/min的頻率振蕩10 min,沖洗收集各級篩層上的團聚體于鋁盒中,<0.053 mm的水穩性團聚體則需要在桶內靜置沉淀72 h,隨后棄去上清液,將團聚體轉移至鋁盒內,在60 ℃下烘干,稱重,計算水穩性團聚體的質量分數。
脲酶采用苯酚鈉-次氯酸鈉比色法測定,用24 h 后1 g土壤中銨態氮的毫克數表示;蔗糖酶采用3,5-二硝基水楊酸比色法測定,用24 h后1 g土壤中葡萄糖的毫克數表示[12];酸性磷酸酶采用對硝基苯磷酸鹽法測定,用1 h后土壤所釋放酚的毫克數表示[13]。
土壤酶活性的幾何平均數(Mg)計算公式[14]為:
1.3 數據分析
利用SPSS 20.0對數據進行統計分析,采用單因素方差分析,并用LSD法對不同森林類型、不同土層、不同粒徑之間進行差異性檢驗,采用SigmaPlot 12.5數學軟件進行繪圖。
2 結果與分析
2.1 土壤水穩性團聚體組成特征
由圖1可知,隨月份變化,興安落葉松林0~5 cm土層以>0.5 mm粒徑團聚體占絕對優勢,且至8月出現最高值84.48%;0.053~0.25 mm和<0.053 mm粒徑團聚體含量均在6月達到最高值為16.14%和24.47%。5~10 cm和10~20 cm土層均以<0.25 mm微團聚體(0.053~0.25 mm;<0.053 mm)為主,呈波動式上升以生長季后期含量較高,分別在9月和10月達到最高值79.94%和93.49%;>0.25 mm大團聚體含量較低,其中7—10月10~20 cm土層沒有得到>1 mm粒徑團聚體。樟子松林0~5 cm土層>1 mm粒徑團聚體在5—9月差異不顯著(P>0.05),平均值為53.76%;0.5~1 mm和0.25~0.5 mm粒徑團聚體均在6月達到最高值26.91%和7.97%;微團聚體含量較低,波動為12.55%~27.59%。5~10 cm土層>1 mm粒徑團聚體呈波動式上升趨勢,在10月出現最高值43.09%;0.5~1 mm粒徑團聚體以6月值最高為27.03%,大團聚體約占團聚體總量的58%以上,與微團聚體差異減小。10~20 cm土層以微團聚體為主,其中0.053~0.25 mm粒徑團聚體5月值最低,為19.33%,6—10月波動為39.39%~58.07%;<0.053 mm粒徑團聚體波動為12.05%~38.22%。
總之,興安落葉松林大團聚體含量低于樟子松林,興安落葉松林0~5 cm土層和樟子松林0~10 cm土層大團聚體約占團聚體總量的60%以上,其他土層以微團聚體含量較高,兩種針葉林生長季中期大團聚體含量相對初期和末期更高。
2.2 土壤水穩性團聚體酶活性
2.2.1 脲酶活性
脲酶是影響土壤中氮轉換的關鍵酶,是植物氮素營養重要的驅動力,在土壤碳氮循環中起著重要作用[15-16]。由圖2可知,興安落葉松林0~5 cm土層,>1 mm粒徑團聚體脲酶活性在6月出現最高值3.29 mg/g;0.5~1 mm粒徑團聚體脲酶生長季中期活性較高,平均達到4.12 mg/g;微團聚體脲酶活性以生長季初期和末期較高,波動范圍為4.10~6.46 mg/g。5~10 cm土層大團聚體脲酶活性呈波動變化趨勢,>1 mm和0.5~1 mm粒徑團聚體分別在10月和5月出現最高值2.11 mg/g和2.74 mg/g;微團聚體9、10月脲酶活性顯著高于其他月份,波動范圍為3.54~4.99 mg/g。10~20 cm土層0.5~1 mm粒徑團聚體脲酶活性以8、9月較高,分別為2.19 mg/g和2.10 mg/g;0.25~0.5 mm 粒徑團聚體在生長季前期脲酶活性較高,波動范圍為1.72~1.93 mg/g;微團聚體生長季初期和末期脲酶活性最高,平均為2.68 mg/g。樟子松林0~5 cm土層>1 mm和0.5~1 mm粒徑團聚體由5月最高值4.62 mg/g和6.70 mg/g波動式下降至10月活性最低;0.25~0.5 mm粒徑團聚體生長季后期脲酶活性較低,各月份間相差較小,波動范圍為3.23~3.82 mg/g;微團聚體以5月脲酶活性最高為10.83 mg/g。5~10 cm土層>1 mm粒徑團聚體脲酶活性由5月最高值3.58 mg/g波動式下降至9月最低值1.33 mg/g后上升;0.5~1 mm和0.25~0.5 mm粒徑團聚體脲酶活性均呈波動式下降趨勢,5月活性最高為3.63 mg/g和7.67 mg/g;0.053~0.25 mm和<0.053 mm粒徑團聚體也以5月脲酶活性最高為5.24 mg/g和7.39 mg/g。10~20 cm土層,>0.25 mm各粒徑團聚體以5月和10月脲酶活性較高,且均在5月出現最高值,波動范圍為 3.19~4.23 mg/g;0.053~0.25 mm和<0.053 mm粒徑團聚體生長季初期脲酶活性顯著高于其他時期,在5月活性最高為5.37 mg/g和5.66 mg/g。總體上,樟子松林團聚體脲酶活性高于興安落葉松林(P<0.05),兩種針葉林均表現為隨土層加深團聚體脲酶活性降低,樟子松林團聚體脲酶隨粒徑的減小活性增加,興安落葉松林隨粒徑減小團聚體脲酶活性呈波動式增加趨勢,以微團聚體脲酶活性較高,興安落葉松林生長季初期和末期活性較高,樟子松林5月團聚體脲酶活性顯著高于其他月份(P<0.05)。
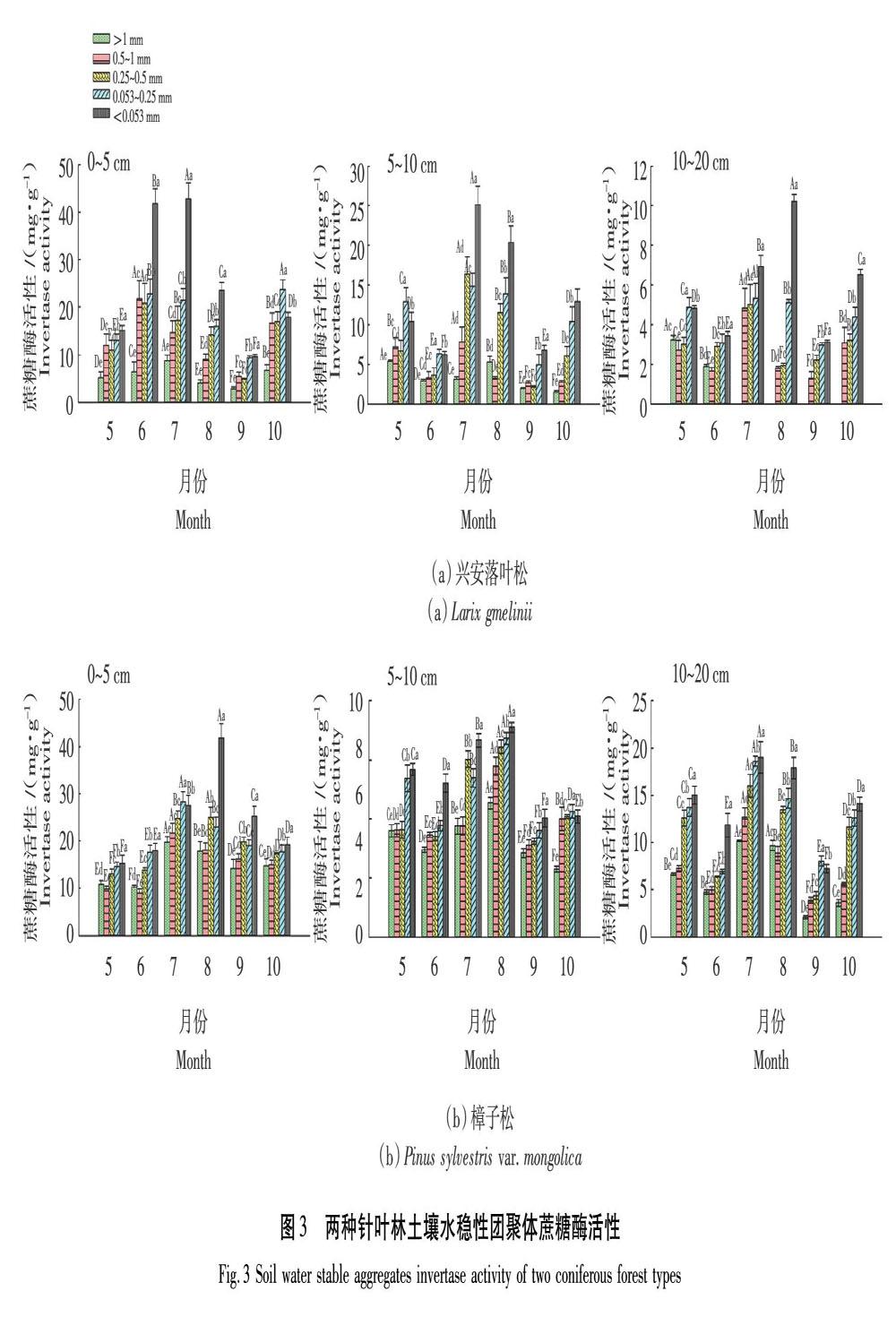
2.2.2 蔗糖酶活性
蔗糖酶能夠參與土壤有機碳的循環,其活性能夠反映有機碳積累轉換的規律[6]。由圖3可知興安落葉松林0~5 cm土層>1 mm和0.5~1 mm粒徑團聚體8、9月蔗糖酶活性顯著低于其他月份,且分別以7月和6月活性最高為8.74 mg/g和21.77 mg/g;0.053~0.25 mm和<0.053 mm粒徑團聚體蔗糖酶以6—8月活性較高,分別波動范圍為16.00~22.73 mg/g和23.59~42.79 mg/g。5~10 cm土層>1 mm和0.5~1 mm粒徑團聚體蔗糖酶活性呈波動變化趨勢,分別在5月和7月達到最高值5.38 mg/g和7.83 mg/g;<0.5 mm各粒徑團聚體蔗糖酶以7月和8月活性較高,且顯著高于其他月份,波動范圍為11.53~25.11 mg/g。10~20 cm土層0.5~1 mm和0.25~0.5 mm粒徑團聚體蔗糖酶以7月活性顯著高于其他月份分別為4.83 mg/g和5.01 mg/g,其他月份則波動范圍為1.30~3.22 mg/g;微團聚體蔗糖酶以生長季中期活性較高,波動范圍為5.13~10.23 mg/g。樟子松林0~5 cm土層各粒徑團聚體蔗糖酶活性均呈上升趨勢在生長季中期出現最高值后下降,其中>1 mm和0.5~1 mm粒徑團聚體蔗糖酶以7月活性最高為19.74 mg/g和21.72 mg/g;<0.053 mm粒徑團聚體8月蔗糖酶活性顯著高于其他月份為41.75 mg/g。5~10 cm土層各粒徑團聚體均以8月活性最高,其中>1 mm和0.5~1 mm粒徑團聚體8月蔗糖酶活性顯著高于其他月份分別為22.70 mg/g和28.92 mg/g,其他月份變化范圍則為11.50~20.01 mg/g;<0.5 mm各粒徑團聚體均表現為生長季中期活性較高,波動范圍為26.94~35.52 mg/g。10~20 cm土層各粒徑團聚體蔗糖酶活性呈波動變化趨勢,且均在7月達到最高值,>0.25 mm各粒徑團聚體7月蔗糖酶活性范圍為10.17~16.03 mg/g,0.053~0.25 mm和<0.053 mm粒徑團聚體分別為18.60 mg/g和19.01 mg/g。總之,樟子松林團聚體蔗糖酶活性高于興安落葉松林(P<0.05),2種針葉林團聚體蔗糖酶活性隨土層加深逐漸降低,且隨粒徑的減小活性增加;在生長季期間呈波動變化趨勢,興安落葉松林0~5 cm土層以6、7月活性較高,5~20 cm土層和樟子松林整個土壤剖面以7、8月活性較高。
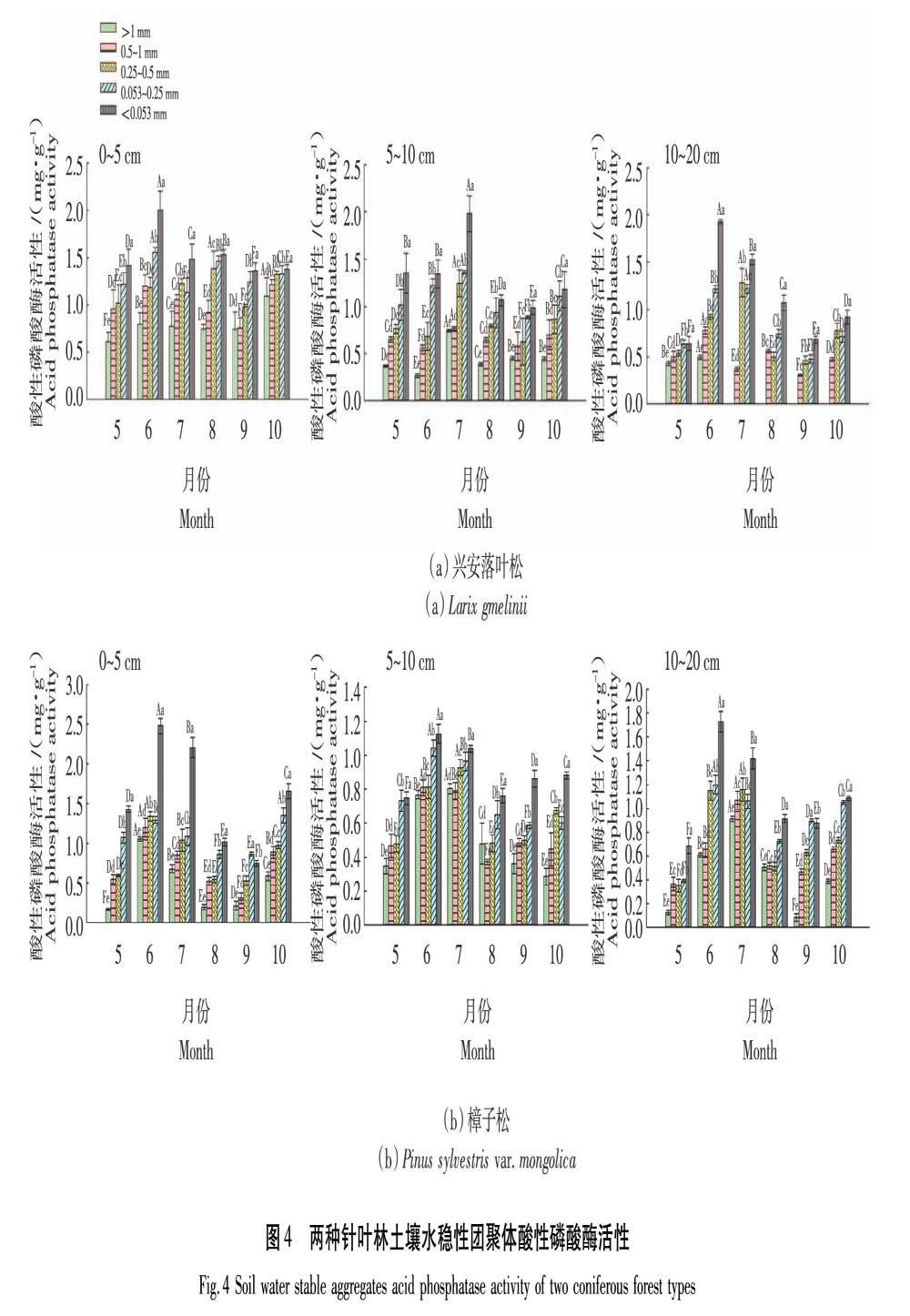
2.2.3 酸性磷酸酶活性
磷酸酶可以促進磷酸酯的水解反應,將土壤有機磷轉化后供根系吸收,其活性影響著土壤有機磷的生物有效性[17]。由圖4可知,興安落葉松林0~5 cm土層>1 mm和0.5~1 mm粒徑團聚體酸性磷酸酶活性呈波動變化趨勢,在10月出現最高值1.10 mg/g和1.22 mg/g;0.25~0.5 mm粒徑團聚體酸性磷酸酶活性呈上升趨勢至8月最高值1.39 mg/g后降低;<0.25 mm各粒徑微團聚體酸性磷酸酶活性呈波動變化趨勢,波動范圍為1.22~2.00 mg/g。5~10 cm土層各粒徑團聚體酸性磷酸酶活性均先波動上升至7月出現最高值后下降,10月再次上升;7月>1 mm和0.5~1 mm粒徑團聚體酸性磷酸酶活性差異較小,平均達到0.76 mg/g,其他粒徑團聚體波動范圍為1.25~1.98 mg/g。10~20 cm土層0.5~1 mm粒徑團聚體6月酸性磷酸酶活性顯著高于其他月份為0.78 mg/g,0.053~0.25 mm和<0.053 mm粒徑團聚體也以6—8月酸性磷酸酶活性較高,波動范圍為0.75~1.93 mg/g。樟子松林0~5 cm 土層大團聚體酸性磷酸酶6月活性最高達到1.20 mg/g,且顯著高于其他月份;0.053~0.25 mm粒徑團聚體酸性磷酸酶活性呈波動式上升趨勢,10月活性最高為1.35 mg/g;<0.053 mm粒徑團聚體6、7月活性較高,分別為2.48 mg/g和2.21 mg/g,其他月份則波動范圍為0.74~1.66 mg/g。5~10 cm土層>1 mm和0.5~1 mm粒徑團聚體酸性磷酸酶活性先分別上升至7月和6月出現最高值0.80 mg/g和0.81 mg/g后,波動式下降至10月活性最低;0.053~0.25 mm和<0.053 mm粒徑團聚體在6月酸性磷酸酶活性最高,分別為1.04 mg/g和1.13 mg/g 。10~20 cm土層各粒徑團聚體呈波動變化趨勢,其中>0.25 mm各粒徑團聚體在7月酸性磷酸酶活性最高,波動范圍為0.91~1.16 mg/g;0.053~0.25 mm和<0.053 mm粒徑團聚體酸性磷酸酶6月活性最高,分別為1.20 mg/g和1.73 mg/g,且10~20 cm土層部分團聚體酸性磷酸酶活性高于5~10 cm土層。興安落葉松林團聚體酸性磷酸酶活性高于樟子松林(P<0.05);2種林型團聚體酸性磷酸酶活性隨土層下降而降低,但相對脲酶和蔗糖酶降幅較小,10~20 cm土層部分粒徑酸性磷酸酶活性高于5~10 cm土層,隨著粒徑的減小活性增加,且以6、7月活性較高。
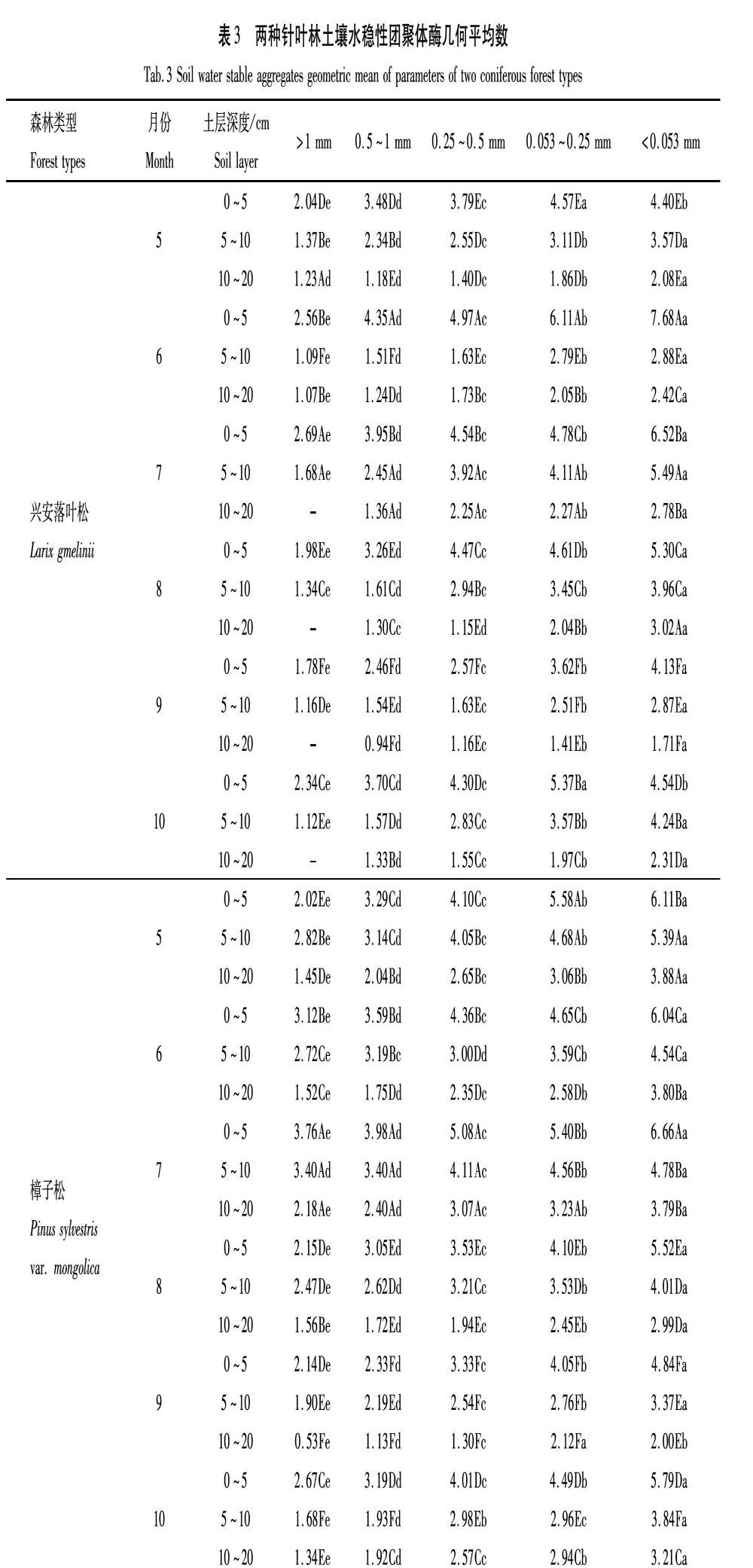
2.3 土壤團聚體酶活性幾何平均數
由表3可知,土壤酶活性的幾何平均數是指示土壤生物質量,綜合評價酶活性的指標,可將多個生物性指標簡化為單一的值來反映土壤環境的變化[18]。興安落葉松林和樟子松林均以微團聚體酶活性幾何平均數較高,隨粒徑的增大而逐漸降低。興安落葉松林0~5 cm土層>1 mm粒徑團聚體酶活性幾何平均數在7月出現最高值2.69,其他粒徑均在6月值最高,變化范圍為4.35~7.68;5~10 cm和10~20 cm土層均以7月團聚體酶活性幾何平均數最高,分別波動范圍為1.68~5.49和1.36~2.79。樟子松林0~5 cm土層0.053~0.25 mm粒徑團聚體5月團聚體酶活性幾何平均數最高為5.58,與7月差異較小,而其他粒徑均在7月達到最高值,波動范圍為3.76~6.66;5~10 cm土層>0.25 mm粒徑團聚體酶活性幾何平均數以7月值最高,變化范圍為3.40~4.11,0.053~0.25 mm和<0.053 mm粒徑團聚體則以5月酶活性幾何平均數最高為4.68和5.39;10~20 cm土層<0.053 mm粒徑團聚體5月酶活性幾何平均數最高為3.88,其他粒徑在7月達到最高值,波動范圍為2.18~3.23。
總之,樟子松林團聚體酶活性幾何平均數高于興安落葉松林, 2種針葉林團聚體酶活性幾何平均數大致以生長季中期較高,且隨著土層的加深數值顯著降低,隨粒徑增大數值降低。
3 討論
土壤團聚體是土壤成分的基本單元,其含量與分布決定了土壤儲存養分及穩定性等能力的高低[19-20]。本研究中,樟子松林大團聚體含量高于興安落葉松林,且兩種林型皆表現為隨著土層的加深,大團聚體含量逐漸降低,這是由于有機質能夠為土壤團聚體的形成提供膠結物質,而土壤表層匯集大量的枯枝落葉,為土壤提供大量的腐殖質和碳源,使得上層土壤含有大量的有機質,增加了上層土壤中的大團聚體含量[21-22],而樟子松林土壤有機碳和全氮含量高于興安落葉松林,因此大團聚體含量也相對較高[23]。本研究兩種林型以7—9月生長旺季大團聚體含量相對較高,可能是由于生長季初期和末期仍受凍融作用影響,土壤處于凍結狀態,冰晶的膨脹破壞了土壤顆粒間原有的聯結狀態,使大粒徑的團聚體受到破壞[24],且有研究認為低溫會使土壤微生物休眠甚至死亡,降低微生物活動速率,凋落物分解緩慢,對土壤有機碳的補充降低,使得大團聚體含量降低[25]。且7—9月為生長旺季,植物生長旺盛,土壤水氣狀況良好,對養分的輸送起到重要作用,有利于團聚體的膠結。
土壤酶主要來源于微生物殘留以及植物根系的分泌物,影響著土壤養分的轉化循環,能夠反映土壤營養物質的動態變化[26]。土壤團聚體對酶活性的分布具有重要的影響,不同粒徑團聚體中酶的活性存在差異[27]。本研究中,脲酶、蔗糖酶和酸性磷酸酶隨著粒徑的減小活性增大,以<0.25 mm粒徑團聚體酶活性最高。有研究表明,大團聚體不利于酶活性的積累[28],此外小團聚體會先于大團聚體形成,隨后膠結形成大團聚體,包裹在大團聚體中的小團聚體具有較穩定的有機物、酶和底物,隨著微生物的生長繁殖,促進酶與底物相互作用,提高小團聚體中酶的活性[6]。而且土壤中的脲酶和蔗糖酶大多是由細菌分泌產生,而有研究表明土壤黏粒中細菌多樣性與豐富度最高[29],有助于小團聚體中脲酶和蔗糖酶活性的增強。
不同森林類型下土壤溫濕度狀況及土壤養分與微生物種類的差異均會導致酶活性的不同[30],本研究中樟子松林土壤團聚體酶幾何平均數、脲酶和蔗糖酶的活性均高于興安落葉松林,可能是因為樟子松林土壤養分含量相對較高,導致酶的底物增加,進而誘導酶活性的增強[31],而興安落葉松林酸性磷酸酶活性高于樟子松林,可能是因為落葉松林凋落物中含有不易分解的木質素、樹脂等,分解產生強酸性的富里酸水,導致土壤酸性物質的富集,而酸性磷酸酶在酸性土壤中更占優勢,從而使得興安落葉松林酸性磷酸酶相對樟子松林活性更高[32-33]。兩種林型土壤團聚體酶活性幾何平均數以及3種酶活性均隨著土層的加深而降低,由于表層土壤根系豐富,擁有大量凋落物,導致表層土壤養分含量高,易分解物質較多,酶底物充足,有利于酶活性的提高。另一方面,上層土壤的水熱通氣狀況較好,有利于微生物的生長繁殖,而土壤微生物代謝產酶能力較強,所以上層土壤具有較高的酶活性,隨著土層的加深,土壤根系分布與養分含量減少,且微生物的生長環境惡化,因此土壤酶活性逐漸降低[34-35]。蔗糖酶和酸性磷酸酶均表現為生長季中期活性較高,夏季較高的土壤溫度與良好的通氣狀況有利于凋落物的分解及細根的周轉,導致土壤酶底物有效性增強,從而促進酶的活性。有研究認為,低溫會使土壤酶處于低活性水平甚至失活,但本研究在生長季初期和末期溫度較低的時期,脲酶活性相對于生長季中期更高,原因可能是大興安嶺地區存在凍融現象,凍融交替過程中增加了嗜冷生物,低溫時期仍存活的嗜冷生物為脲酶提供了底物,使得其在生長季初期和末期活性仍維持在較高水平[36]。
4 結論
大興安嶺北部兩種針葉林土壤隨著土層的加深大團聚體含量下降,微團聚體含量增加,生長季中期大團聚體含量高于生長季初期和末期,但興安落葉松林土壤大團聚體含量高于樟子松林。樟子松林團聚體脲酶和蔗糖酶活性高于興安落葉松林,而興安落葉松林團聚體酸性磷酸酶活性更高;興安落葉松林團聚體脲酶生長季初期和末期活性均較高,樟子松林則以5月團聚體脲酶活性顯著高于其他月份,兩種針葉林團聚體蔗糖酶和酸性磷酸酶活性均以生長季中期較高;隨著土層的加深3種團聚體酶活性均表現出逐漸降低的趨勢,且均以微團聚體酶活性更高。兩種針葉林團聚體酶活性幾何平均數大致以生長季中期較高,且隨著土層的加深數值降低。總體而言,樟子松林團聚體酶活性幾何平均數高于興安落葉松林。
【參 考 文 獻】
[1]王玉功,劉婧晶,劉貽熙,等.苯酚-次氯酸鈉比色法測定土壤脲酶活性影響因素的研究[J].土壤通報,2019,50(5):1166-1170.
WANG Y G, LIU J J, LIU Y X, et al. Effective factors of urease activities in soil by using the phenol-sodium hypochlorite clorimetric method[J]. Chinese Journal of Soil Science, 2019, 50(5):1166-1170.
[2]馬亞娟,徐福利,王渭玲,等.氮磷提高華北落葉松人工林地土壤養分和酶活性的作用[J].植物營養與肥料學報,2015,21(3):664-674.
MA Y J, XU F L, WANG W L, et al. Increase of soil nutrients and enzymatic activity by adding nitrogen and phosphorus to Larix principis-rupprechtii plantation[J]. Journal of Plant Nutrition and Fertilizers, 2015, 21(3):664-674.
[3]李瑞瑞,盧藝,王益明,等.氮添加對墨西哥柏人工林土壤碳氮磷化學計量特征及酶活性的影響[J].生態學雜志,2019,38(2):384-393.
LI R R, LU Y, WANG Y M, et al. Effects of N addition on C, N and P stoichiometry and soil enzyme activities in Cupressus lusitanica Mill. plantation[J]. Chinese Journal of Ecology, 2019, 38(2):384-393.
[4]劉超,趙光影,宋艷宇,等.氣候變化背景下濕地土壤酶活性研究進展[J].中國農學通報,2019,35(33):91-97.
LIU C, ZHAO G Y, SONG Y Y, et al. Soil enzyme activity in wetland under the background of climate change: research progress[J]. Chinese Agricultural Science Bulletin, 2019, 35(33): 91-97.
[5]陳立明,滿秀玲.小興安嶺谷地云冷杉林土壤酶活性的異質性[J].森林工程,2010,26(1):1-6,11.
CHEN L M, MAN X L. Heterogeneity of soil enzyme activity in Abies and Picea asperata forest in low-lying land of Xiaoxinganling Mountain[J]. Forest Engineering, 2010, 26(1): 1-6, 11.
[6]齊思明,韓瑛,陳祥偉.植被恢復對典型黑土表層土壤團聚體水解酶活性的影響[J].東北林業大學學報,2017,45(6):42-46.
QI S M, HAN Y, CHEN X W. Effect of vegetation restoration on soil aggregates enzyme activity in typical black soil region[J]. Journal of Northeast Forestry University, 2017, 45(6): 42-46.
[7]柳開樓,韓天富,胡惠文,等.紅壤旱地玉米開花期土壤酶活性對長期施肥的響應[J].植物營養與肥料學報,2018,24(6):1610-1618.
LIU K L, HAN T F, HU H W, et al. Response of soil enzyme activity in flowering stages of maize to long-term fertilization in red soil[J]. Journal of Plant Nutrition and Fertilizers, 2018, 24(6):1610-1618.
[8]馬瑞萍,安韶山,黨廷輝,等.黃土高原不同植物群落土壤團聚體中有機碳和酶活性研究[J].土壤學報,2014,51(1):104-113.
MA R P, AN S S, DANG T H, et al. Soil organic carbon and enzymatic activity in aggregates of soils under different plant communities in hilly-gully regions of loess plateau[J]. Acta Pedologica Sinica, 2014, 51(1): 104-113.
[9]葉鈺倩,趙家豪,劉暢,等.間伐對馬尾松人工林根際土壤氮含量及酶活性的影響[J].南京林業大學學報(自然科學版),2018,42(3):193-198.
YE Y Q,ZHAO J H,LIU C,et al.Effects of thinning on nitrogen contents and enzyme activitiesof rhizosphere soil in Pinus massoniana plantations[J].Journal of Nanjing Forestry University(Natural Science Edition),2018,42(3):193-198.
[10]劉先,索沛蘅,杜大俊,等.連栽杉木人工林參與土壤碳氮轉化過程酶活性及其與土壤理化因子相關性[J].生態學報,2020,38(1):1-10.
LIU X, SUO P H, DU D J, et al. Enzyme activity and its correlation with soil physical and chemical factors involved in soil carbon and nitrogen transformation of Chinese fir plantation[J]. Acta Ecologica Sinica, 2020, 38(1): 1-10.
[11]丁令智,滿秀玲,肖瑞晗,等.寒溫帶森林根際土壤微生物量碳氮含量生長季內動態變化[J].林業科學,2019,55(7):178-186.
DING L Z, MAN X L, XIAO R H, et al. Dynamics of soil microbial biomass carbon and nitrogen in the soil of rhizosphere during growing season in the cold temperate forests[J]. Scientia Silvae Sinicae, 2019, 55(7): 178-186.
[12]關松蔭.土壤酶及其研究法[M].北京:農業出版社,1986.
GUAN S Y. Soil enzyme and its research method[M]. Beijing: Agricultural Press, 1986.
[13]魯如坤.土壤農業化學分析方法[M].北京:中國農業科技出版社,2000.
LU R K. Methods for agricultural chemical analysis of soil[M]. Beijing: China Agriculture Scientech Press, 2000.
[14]尚藝婕,張秀,王海波,等.秸稈生物質炭對鎘污染水稻土根際酶活性的影響[J].農業環境科學學報,2016,35(8):1532-1540.
SHANG Y J, ZHANG X, WANG H B, et al. Effects of straw biochar on rhizospheric enzyme activities in Cd contaminated paddy soil[J]. Journal of Agro-Environment Science, 2016, 35(8): 1532-1540.
[15]李龍,辛貴民,杜彥梅,等.春季凍融對2種溫帶森林土壤酶活性和土壤微生物生物量的影響[J].揚州大學學報(農業與生命科學版),2019,40(1):26-32.
LI L, XIN G M, DU Y M, et al. Effects of spring freezing and thawing on soil enzyme activities and soil microbial biomass in 2 temperate forests[J]. Journal of Yangzhou University (Agricultural and Life Science Edition), 2019, 40(1): 26-32.
[16]李聰,孫正峰,曹宇,等.林區不同土地利用模式對土壤理化性質與酶學特性的影響[J].森林工程,2013,29(3):18-24.
LI C, SUN Z F, CAO Y, et al. Effects on physicochemical properties and enzymatic activities of soil by different land utilization patterns in forest region[J]. Forest Engineering, 2013, 29(3): 18-24.
[17]蔡小溪,吳金卓.森林土壤健康評價研究進展[J].森林工程,2015,31(2):37-41.
CAI X X, WU J Z. Research progress on the assessment of forest soil health[J]. Forest Engineering, 2015, 31(2): 37-41.
[18]賈國梅,席穎,許文年,等.三峽庫區消落帶土壤酶活性特征[J].水土保持研究,2015,22(4):24-28.
JIA G M, XI Y, XU W N, et al. Characteristics of the activities of soil enzymes in riparian zone of Three Gorges Reservoir Area[J]. Research of Soil and Water Conservation, 2015, 22(4):24-28.
[19]蘇思慧,王美佳,張文可,等.耕作方式與玉米秸稈條帶還田對土壤水穩性團聚體和有機碳分布的影響[J].土壤通報,2018,49(4):841-847.
SU S H, WANG M J, ZHANG W K, et al. Effects of tillage practices and maize straw incorporation on water-stable aggregates and organic carbon in soils[J]. Chinese Journal of Soil Science, 2018, 49(4): 841-847.
[20]李程程,曾全超,賈培龍,等.黃土高原土壤團聚體穩定性及抗蝕性能力經度變化特征[J].生態學報,2020,35(6):1-10.
LI C C, ZENG Q C, JIA P L, et al. Characteristics of soil aggregate stability and corrosion resistance longitude change in the Loess Plateau[J]. Acta Ecologica Sinica, 2020, 35(6): 1-10.
[21]谷忠元,康黎,羅夢娟,等.湘東地區典型土壤團聚體穩定性的影響因素[J].水土保持通報,2018,38(5):58-63.
GU Z Y, KANG L, LUO M J, et al. Factors affecting stability of soil aggregate in eastern Hunan Province[J]. Bulletin of Soil and Water Conservation, 2018, 38(5):58-63.
[22]孫穎,徐嘉暉,高菲,等.長白山森林土壤有機碳及其在團聚體密度組分中的分布[J].森林工程,2018,34(2):1-5.
SUN Y, XU J H, GAO F, et al. Organic carbon content and its distribution in aggregate-density fractions of forest soils in Changbai mountain[J]. Forest Engineering, 2018, 34(2): 1-5.
[23]蔣臘梅,白桂芬,呂光輝,等.不同管理模式對干旱區草原土壤團聚體穩定性及其理化性質的影響[J].干旱地區農業研究,2018,36(4):15-21.
JIANG L M, BAI G F, LYU G H, et al. Effects of different management modes on soil aggregate stability and physicochemical properties of grassland in arid area[J]. Agricultural Research in the Arid Areas, 2018, 36(4): 15-21.
[24]金萬鵬,范昊明,劉博,等.凍融交替對黑土團聚體穩定性的影響[J].應用生態學報,2019,30(12):4195-4201.
JIN W P, FAN H M, LIU B, et al. Effects of freeze-thaw cycles on aggregate stability of black soil[J]. Chinese Journal of Applied Ecology, 2019, 30(12):4195-4201.
[25]尹寶絲,史常青,賀康寧,等.高寒區華北落葉松林生長季內地表凋落物層碳氮磷化學計量特征[J].應用與環境生物學報,2019,25(2):268-274.
YIN B S, SHI C Q, HE K N, et al. Litter carbon, nitrogen, and phosphorus stoichiometry of Larix principis-rupprechtii in alpine region during growing season[J]. Chinese Journal of Applied and Environmental Biology, 2019, 25(2): 268-274.
[26]邵文山,李國旗.土壤酶功能及測定方法研究進展[J].北方園藝,2016,39(9):188-193.
SHAO W S, LI G Q. Research progress of soil enzymes function and its determination method[J]. Northern Horticulture, 2016, 39(9):188-193.
[27]姬秀云,李玉華.黃土高原植被恢復對不同粒徑土壤團聚體中酶活性的影響[J].水土保持通報,2018,38(1):24-28.
JI X Y, LI Y H. Effects of different revegetation types on soil enzyme activities in different aggregates fractions in Loess Plateau[J]. Bulletin of Soil and Water Conservation, 2018, 38(1):24-28.
[28]NIE M, PENDALL E, BELL C, et al. Soil aggregate size distribution mediates microbial climate change feedbacks[J]. Soil Biology and Biochemistry, 2014, 68(1): 357-365.
[29]ZHANG P J, ZHENG J F, PAN G X, et al. Changes in microbial community structure and function within particle size fractions of a paddy soil under different long-term fertilization treatments from the Tai Lake region, China[J]. Colloids and Surfaces B: Biointerfaces, 2007, 58(2): 264-270.
[30]SOUZA R C, SOLLY E F, DAWES M A, et al. Responses of soil extracellular enzyme activities to experimental warming and CO2 enrichment at the alpine treeline[J]. Plant and Soil, 2017, 416(1): 527-537.
[31]趙海燕,徐福利,王渭玲,等.秦嶺地區華北落葉松人工林地土壤養分和酶活性變化[J].生態學報,2015,35(4):1086-1094.
ZHAO H Y, XU F L, WANG W L, et al. Soil nutrients and enzyme activities in Larix principis-rupprechtii plantations in the Qinling Mountains, China[J]. Acta Ecologica Sinica, 2015, 35(4): 1086-1094.
[32]李瑩飛,耿玉清,周紅娟,等.基于不同方法測定土壤酸性磷酸酶活性的比較[J].中國生態農業學報,2016,24(1):98-104.
LI Y F, GENG Y Q, ZHOU H J, et al. Comparison of soil acid phosphatase activity determined by different methods[J]. Chinese Journal of Eco-Agriculture, 2016, 24(1): 98-104.
[33]王兵爽,李淑君,張舒桓,等.西瓜根系分泌酸性磷酸酶對有機肥營養的響應[J].土壤學報,2019,56(2):454-465.
WANG B S, LI S J, ZHANG S H, et al. Responses of acid phosphatase secreted by watermelon roots to organic manure nutrition[J]. Acta Pedologica Sinica, 2019, 56(2): 454-465.
[34]趙鵬志,陳祥偉,楊小燕,等.低分子有機酸對東北黑土酶活性與養分關系的影響[J].南京林業大學學報(自然科學版),2018,42(1):105-112.
ZHAO P Z,CHEN X W,YANG X Y,et al.Relationship between enzyme activities and nutrients of black soil subjected to low molecular organic acid[J].Journal of Nanjing Forestry University(Natural Science Edition),2018,42(1):105-112.
[35]杜倩,梁素鈺,李琳,等.闊葉紅松林土壤酶活性及微生物群落功能多樣性分析[J].森林工程,2019,35(1):1-7.
DU Q, LIANG S Y, LI L, et al. Soil enzyme activities and microbial community functional diversity of broad-leaved Korean pine forest[J]. Forest Engineering, 2019, 35(1): 1-7.
[36]李龍,尹航,黃世臣,等.春季解凍期3種溫帶森林土壤酶活性動態變化[J].土壤通報,2018,49(3):609-615.
LI L, YIN H, HUANG S C, et al. Dynamic changes of soil enzyme activities in three types of temperate forest during spring thawing period[J]. Chinese Journal of Soil Science, 2018, 49(3): 609-615.
