納米羥基磷灰石通過溶酶體-線粒體通路誘導(dǎo)腎小管上皮細(xì)胞的凋亡機(jī)制
2020-07-18 09:00:00董文敬張海松徐玉曉崔靜陳玉杰王倩葛坤高燕董文英
實(shí)用醫(yī)學(xué)雜志 2020年12期
關(guān)鍵詞:檢測(cè)
董文敬 張海松 徐玉曉 崔靜 陳玉杰 王倩 葛坤 高燕 董文英
1河北大學(xué)附屬醫(yī)院(河北保定071000);2保定市第一中心醫(yī)院(河北保定071000);3河北大學(xué)(河北保定071000);4吉林大學(xué)(吉林吉林132000)
羥基磷灰石(HAP)是骨骼和牙齒中最重要的無機(jī)礦物成分,但并非所有的HAP 都對(duì)人體有益。在泌尿系統(tǒng),由HAP 誘導(dǎo)形成的Randall 斑塊是促進(jìn)腎結(jié)石形成的重要原因[1]。在Randall 斑塊中存在從大小由納米到微米及不同形態(tài)的HAP[2]。草酸鈣結(jié)石是一種最常見的腎結(jié)石,其形成機(jī)制尚未完全闡明[3]。HAP 參與草酸鈣結(jié)石的形成,以及是如何促進(jìn)腎結(jié)石的形成的機(jī)制研究尚未明確。
草酸鈣在腎乳頭Randall 斑塊上的過度生長(zhǎng)是草酸鈣結(jié)石形成的潛在機(jī)制[4]。EVAN 等[5]通過腎臟活檢發(fā)現(xiàn),Randall 斑起源于髓袢基底膜,隨后擴(kuò)散到了腎直小管,最后到尿路上皮表面,促進(jìn)草酸鹽晶體的成核、生長(zhǎng)和聚集,從而增加了腎結(jié)石形成的風(fēng)險(xiǎn)。在含鈣結(jié)石或尿液中發(fā)現(xiàn)了許多磷酸鈣鹽成分,最常見的磷酸鈣鹽是HAP[6]。因此,HAP 與腎結(jié)石之間存在密切的關(guān)系,腎小管上皮細(xì)胞的氧化炎癥損傷與結(jié)石的形成密切相關(guān)[7]。YU 等[8]通過將HK-2 細(xì)胞與不同濃度的HAP 和/或巨噬細(xì)胞共同培養(yǎng),HAP 增加上調(diào)了HK-2 細(xì)胞中骨橋蛋白(OPN)的表達(dá)水平,并引起異質(zhì)的成核、粘附和晶體沉積,從而促進(jìn)了Randall 斑塊和腎結(jié)石的形成。RAO 等[9]通過研究四種不同類型的HAP(球狀、針狀、棒狀、盤狀),結(jié)果表明HAP 的細(xì)胞毒性:球狀>針狀>棒狀>盤狀,HAP 的物理性質(zhì)對(duì)其細(xì)胞毒性起著至關(guān)重要的作用,同時(shí)發(fā)現(xiàn)HAP 可能激活細(xì)胞氧化應(yīng)激反應(yīng),引發(fā)一系列細(xì)胞功能障礙、凋亡和壞死。本實(shí)驗(yàn)設(shè)計(jì)不同濃度的HAP 探究其通過氧化應(yīng)激反應(yīng)對(duì)HK-2 細(xì)胞的凋亡的影響及其機(jī)制,為泌尿系結(jié)石的機(jī)制研究作一定探討。
1 材料與方法
1.1 主要試劑人源近端腎小管上皮細(xì)胞(HK-2細(xì)胞武漢普諾生命科技有限公司中國(guó))、HAP(水熱法合成、分散性好、棒狀、60~100 nm×12 nm)、MEM 培養(yǎng)基和胎牛血清(Gibco 公司 美國(guó))、胰蛋白酶(Sigma 公司 美國(guó))、Annexin V-FITC/PI 細(xì)胞凋亡和壞死雙染料試劑盒、活細(xì)胞/死細(xì)胞染色試劑盒和DCFH-DA(上海貝博生物有限公司中國(guó))、青霉素和鏈霉素、線粒體膜電位檢測(cè)試劑盒(JC-1)、吖啶橙(AO)染料(碧云天生物技術(shù)公司中國(guó))、四甲基偶氮唑藍(lán)(hiazoyl blue tetrazolium dromide,MTT)(Amresco 公司美國(guó))、Lyso-Tracker Red 和Mito-Tracker Green、過氧化氫(H2O2)試劑盒(賽默飛世爾科技有限公司 中國(guó))、Caspase-3 活性試劑盒(南京建成生物工程研究所中國(guó))。
1.2 實(shí)驗(yàn)方法
1.2.1 HAP 的合成和表征根據(jù)文獻(xiàn),利用水熱法合成了HAP[10]。將樣品分散到無水乙醇中、超聲均勻分散,滴于銅網(wǎng),干燥,用透射電子顯微鏡(TEM)進(jìn)行觀察。將樣品分散到無水乙醇中、超聲均勻分散,滴于硅片,干燥,用掃描電子顯微鏡(SEM)進(jìn)行觀察。取適量的樣品,用X-射線衍粉末射儀(XRD)對(duì)其結(jié)構(gòu)進(jìn)行測(cè)定。取適量的樣品,用固態(tài)熒光測(cè)量?jī)x對(duì)樣品的熒光性質(zhì)進(jìn)行。
1.2.2 細(xì)胞培養(yǎng)將HK-2 細(xì)胞接種到含有10%FBS,100 μg/mL 青霉素和100 μg/mL 鏈霉素的MEM培養(yǎng)基上在37 ℃、5%CO2培養(yǎng)箱中培養(yǎng),胰蛋白酶消化用于細(xì)胞繁殖,細(xì)胞同步后,實(shí)驗(yàn)?zāi)P头纸M:(1)對(duì)照組:僅添加等體積PBS 溶液(0 μg/mL);(2)HAP 處理組:將HK-2 細(xì)胞暴露于5、10、20、40、80 μg/mL 的HAP。
含HAP 的PBS 懸液的制備:將一定量的HAP粉末鋪板,用紫外線滅菌,過夜,然后分散在PBS溶液中,制備濃度為4 mg/mL 的懸浮液,并超聲處理5 min。
1.2.3 HK-2 細(xì)胞存活率和活細(xì)胞/死細(xì)胞染色檢測(cè)通過用MTT 的方法測(cè)定HK-2 細(xì)胞存活率[11]。將HK-2 細(xì)胞(2×104)接種到96 孔板上,按照上述分組分別孵育24、48、72 h,分別加入10 μL MTT,在37 ℃的條件下孵育4 h,棄去上清并加入100 μL DMSO,搖床震蕩10 min,使用酶標(biāo)儀測(cè)定570 nm 的吸光度(OD)值。細(xì)胞活性=OD實(shí)驗(yàn)組/OD對(duì)照組×100%。
將HK-2 細(xì)胞(1×105)接種到6 孔板上,實(shí)驗(yàn)?zāi)P头跤?4h,嚴(yán)格按照活死染試劑盒說明書操作,通過熒光顯微鏡觀察,綠色為活細(xì)胞,紅色為死細(xì)胞。
1.2.4 HK-2 細(xì)胞凋亡測(cè)定Annexin V-FITC/PI 細(xì)胞凋亡和壞死雙染料檢測(cè)細(xì)胞凋亡[12]。將HK-2細(xì)胞(1×105)接種到6 孔板上,實(shí)驗(yàn)?zāi)P头譃閮山M:(1)對(duì)照組:僅添加PBS 溶液(0 μg/mL);(2)HAP處理組:將HK-2 細(xì)胞暴露于10、20、40 μg/mL 的HAP,孵育6、24 h,嚴(yán)格按照試劑盒的說明書步驟操作,通過流式細(xì)胞儀檢測(cè)細(xì)胞凋亡情況。
1.2.5 HAP 在HK-2 細(xì)胞內(nèi)分布分別用Lyso-Tracker Red 或Mito-Tracker Green 追蹤HAP 在HK-2細(xì)胞中的細(xì)胞內(nèi)分布[13]。將HK-2 細(xì)胞(2×104)接種到培養(yǎng)皿上,用HAP(40 μg/mL)處理HK-2 細(xì)胞6 h。用Lyso-Tracker Red 和Mito-Tracker Green分別給溶酶體和線粒體染色,用共聚焦顯微鏡觀察HAP 在細(xì)胞中的分布。
1.2.6 ROS 的檢測(cè)用DCFH-DA 檢測(cè)到細(xì)胞內(nèi)ROS 水平[14]。孵育6 h,將細(xì)胞用200 μL DCFHDA 染色,在37 ℃暗處孵育20 min,PBS 洗滌3 次后,PBS 重懸,通過流式細(xì)胞儀檢測(cè)氧化DCFH-DA的熒光。
嚴(yán)格按H2O2試劑盒說明書步驟操作,使用酶標(biāo)儀在415 nm 下測(cè)量吸光度。實(shí)驗(yàn)?zāi)P头纸M:(1)對(duì)照組:僅添加等體積PBS 溶液(0 μg/mL);(2)HAP 處理組:將HK-2 細(xì)胞暴露于5、10、20、40 μg/mL 的HAP。用5 mmol/L N-乙酰半胱氨酸(NAC)預(yù)處理1 h,使用酶標(biāo)儀在570 nm 下測(cè)量吸光度。細(xì)胞活性=OD實(shí)驗(yàn)組/OD對(duì)照組×100%。
1.2.7 溶酶體完整性檢測(cè)吖啶橙(AO)檢測(cè)溶酶體完整性,孵育6 h,用PBS 制備的5 μg/mL 吖啶橙染料(AO)溶液染色15 min,通過流式細(xì)胞儀檢測(cè)通透性。
用Lyso-Tracker Red 檢測(cè)溶酶體通透性的改變。加入Lyso-Tracker Red 和Hochest,分別孵育30 min和15 min。用熒光顯微鏡觀察溶酶體的堿化。
1.2.8 線粒體膜電位(ΔΨm)和Caspase-3 檢測(cè)將HK-2 細(xì)胞(每孔1×105個(gè)細(xì)胞)接種在6 孔板中,同2.2.4 實(shí)驗(yàn)?zāi)P头纸M,孵育6 h,收集細(xì)胞并以1 000 rpm/min 離心5 min,吸出上清液,用PBS洗滌,再次離心獲得細(xì)胞沉淀,加入500 μL JC-1,在黑暗中于37 ℃孵育20 min,PBS 洗滌三次除去未反應(yīng)的JC-1,并通過流式細(xì)胞儀檢測(cè)分析細(xì)胞的線粒體膜電位的變化。
將HK-2 細(xì)胞(每孔1×105個(gè)細(xì)胞)接種在6 孔板中,孵育6 h,嚴(yán)格按照說明書裂解細(xì)胞,取少量細(xì)胞樣品用BCA 蛋白濃度定量試劑盒定量蛋白濃度以保證每組的蛋白濃度在100~200 μg。Caspase-3 活性測(cè)定嚴(yán)格按說明書操作。用酶標(biāo)儀測(cè)定波長(zhǎng)為405 nm 的吸光度(OD)。Caspase-3 活化程度=OD樣品組/OD對(duì)照組×100%。
1.3 統(tǒng)計(jì)學(xué)方法所有結(jié)果至少作三個(gè)獨(dú)立平行實(shí)驗(yàn)。數(shù)據(jù)表示為均數(shù)±標(biāo)準(zhǔn)差。單因素方差分析用于多重比較,兩組間比較用獨(dú)立樣本t檢驗(yàn)。P<0.05 為差異有統(tǒng)計(jì)學(xué)意義。
2 結(jié)果
2.1 HAP 表征通過TEM 和SEM 對(duì)HAP 的形貌和大小進(jìn)行測(cè)定。HAP 為分散性良好的棒狀,長(zhǎng)度×高度:(60~100)nm×12 nm(圖1a、b)。利用XRD 對(duì)HAP 的結(jié)構(gòu)進(jìn)行測(cè)定,通過與標(biāo)準(zhǔn)衍射圖(JCPDS No.09-0432 標(biāo)準(zhǔn)衍射圖)對(duì)比,HAP 與標(biāo)準(zhǔn)衍射圖一致,表明合成了純的HAP(圖1c)。利用固態(tài)熒光測(cè)量?jī)x檢測(cè)HAP 發(fā)藍(lán)光(圖1d)。

圖1 HAP 表征結(jié)果Fig.1 HAP characterization results
2.2 HAP 對(duì)的HK-2 細(xì)胞存活率影響MTT 法觀察不同劑量HAP 處理24、48 和72 h 對(duì)HK-2 細(xì)胞生存率的影響,見圖2a。0、5、10、20、40和80 μg/mL的HAP 處理的HK-2 細(xì)胞在24、48 和72 h 對(duì)HK-2細(xì)胞增殖均有明顯的抑制作用,并呈明顯的劑量依賴性。在24 h,HAP 在5、10 μg/mL 細(xì)胞存活率無明顯差異,約降低17%~19%,80 μg/mL 細(xì)胞存活率約降低60%。本結(jié)果顯示,HAP 抑制HK-2 細(xì)胞增殖,降低細(xì)胞生存率且引起細(xì)胞產(chǎn)生細(xì)胞毒性且呈濃度-時(shí)間依賴性。
活細(xì)胞/死細(xì)胞染色結(jié)果表明HK-2 細(xì)胞增殖能力隨濃度的增加而降低(圖2b)HAP 在20 μg/mL后釋放紅色熒光的死細(xì)胞逐漸增多,釋放綠色熒光的活細(xì)胞逐漸減少。HAP 在5、10 μg/mL 活/死細(xì)胞未觀察到明顯差異,80 μg/mL 時(shí)僅觀察到了少量的活細(xì)胞。活/死細(xì)胞染色結(jié)果與MTT 結(jié)果基本一致。
2.3 HAP 引起的HK-2 細(xì)胞調(diào)亡由于80 μg/mL的HAP 明顯的抑制HK-2 細(xì)胞的增殖并導(dǎo)致壞死脫落。結(jié)合細(xì)胞活性和活死染結(jié)果,選擇0、10、20、40 μg/mL 的HAP 分別作用于HK-2 細(xì)胞共孵育6、24 h,凋亡結(jié)果表明6 h 細(xì)胞的存活率沒有明顯差異(圖2c),24 h 凋亡結(jié)果表明細(xì)胞的凋亡和壞死率隨著濃度的增加而升高(圖2c、d)。
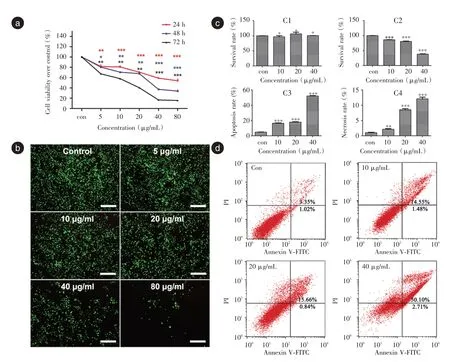
圖2 HK-2 細(xì)胞凋亡結(jié)果Fig.2 Results HK-2 cell apoptosis
2.4 HAP 在HK-2 細(xì)胞中的分布見圖3,HAP的藍(lán)色熒光主要與Lyso-Tracker 標(biāo)記的溶酶體的紅色熒光重疊,與Mito-Tracker 標(biāo)記的綠色熒光僅有部分局限,結(jié)果表明HAP 主要分布在溶酶體。
2.5 HAP 誘導(dǎo)HK-2 細(xì)胞ROS 的釋放DCFHDA 熒光探針檢測(cè)了HAP 誘導(dǎo)的HK-2 細(xì)胞中ROS的變化。熒光強(qiáng)度的定量結(jié)果表明,HK-2 細(xì)胞中的ROS 水平隨濃度增加增加(圖4a)。HK-2 細(xì)胞中的H2O2水平隨濃度增加增加(圖4b)。正常組的細(xì)胞具有暗熒光,表明細(xì)胞內(nèi)ROS 水平低。HAP治療組的綠色熒光均顯著增強(qiáng)(圖4d),表明HAP導(dǎo)致細(xì)胞內(nèi)ROS 水平升高。乙酰半胱氨酸(NAC)是一種抗氧化劑,加入NAC,HAP 誘導(dǎo)的細(xì)胞活性較未加入NAC 的細(xì)胞生存率升高(圖4c),表明細(xì)胞的生存率與ROS 釋放有關(guān),HAP 在5、10 μg/mL細(xì)胞存活率無明顯差異。

圖3 在暴露于40 μg/mL 的HAP 中6 h 后,HK-2 細(xì)胞中HAP 的分布(標(biāo)尺:10 μm)Fig.3 HAP distribution in HK-2 cells after 6 hours of exposure to HAP at 40 μg/mL(Scale:10 μm)
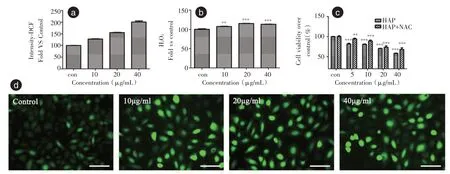
圖4 HK-2 細(xì)胞ROS 的釋放Fig.4 ROS release from HK-2 cells
2.6 HAP 對(duì)HK-2 細(xì)胞溶酶體膜完整性的影響見圖5a。HAP 可使HK-2 細(xì)胞溶酶體通透性增加,溶酶體內(nèi)的大量的水解酶釋放,溶酶體膜受損。Lyso-Tracker Red 是一種堿性染料,溶酶體紅在溶酶體內(nèi)的熒光強(qiáng)弱可間接反應(yīng)溶酶體的完整性,見圖5d。HAP 可使HK-2 細(xì)胞溶酶體完整性受損。結(jié)果表明HAP 可使HK-2 細(xì)胞溶酶體完整性受損,且呈濃度依賴性。
2.7 HAP 對(duì)HK-2 細(xì)胞線粒體功能的影響見圖5b,所示隨HAP 濃度的增加ΔΨm 降低。HAP引起的細(xì)胞內(nèi)ROS 水平升高可能導(dǎo)致ΔΨm降低。
2.8 caspase-3 的釋放caspase-3 是細(xì)胞凋亡的過程中的一種關(guān)鍵酶。見圖5c,所示隨HAP 濃度的增加caspase-3 增加。由HAP 引起的細(xì)胞ΔΨm 降低可能導(dǎo)致caspase-3 激活。
3 討論
腎結(jié)石是一種日益增長(zhǎng)的泌尿系統(tǒng)疾病,約占世界人口的5%~13%[15]。目前治療腎結(jié)石的主要方法是外科手術(shù)[16],但患者的復(fù)發(fā)率高,5年復(fù)發(fā)率約為50%[17]。因此,了解腎結(jié)石的發(fā)病機(jī)制對(duì)結(jié)石的治療和預(yù)防復(fù)發(fā)具有重要意義。目前國(guó)內(nèi)外主要集中在草酸鈣在腎結(jié)石形成機(jī)制的探究[18],而關(guān)于HAP 作為促進(jìn)腎結(jié)石形成的機(jī)制的研究較少。

圖5 HAP 對(duì)HK-2 細(xì)胞線粒體功能和細(xì)胞溶酶體膜完整性的影響Fig.5 Effect of HAP on mitochondrial function and lysosomal membrane integrity of HK-2 cells
近年來國(guó)內(nèi)外出現(xiàn)了關(guān)于HAP 對(duì)腎上皮細(xì)胞損傷的報(bào)道,余駿川等[19]研究了HAP 對(duì)HK-2 細(xì)胞的損傷作用,發(fā)現(xiàn)細(xì)胞存活率與HAP 的濃度呈負(fù)相關(guān)且通過上調(diào)細(xì)胞內(nèi)的OPN 表達(dá)參與腎結(jié)石的形成。COHENA 等[20]將草酸,草酸鈣和HAP與犬腎細(xì)胞(MDCK)共培養(yǎng),結(jié)果表明它們都引起MDCK 細(xì)胞中基質(zhì)Gla 蛋白(MGP)表達(dá)增加,超氧化物歧化酶釋放,乳酸脫氫酶釋放,并促進(jìn)細(xì)胞凋亡率,從而促進(jìn)腎結(jié)石的形成。研究表明HAP 對(duì)腎上皮細(xì)胞造成損傷,但并未對(duì)其造成損傷的機(jī)制進(jìn)一步研究。有研究表明當(dāng)300 μg/mL的HAP 會(huì)對(duì)HK-2 細(xì)胞造成不可逆的損傷,造成大量細(xì)胞凋亡[21],所以設(shè)計(jì)的最大濃度不高于200 μg/mL,本研究通過MTT 和活細(xì)胞/死細(xì)胞染色實(shí)驗(yàn),發(fā)現(xiàn)HAP 可以降低HK-2 細(xì)胞的存活率,抑制細(xì)胞增殖,表明對(duì)HK-2 細(xì)胞具有一定的損傷作用。
研究表明細(xì)胞內(nèi)的氧化應(yīng)激反應(yīng)能引起細(xì)胞氧化炎癥損傷,過量的ROS 會(huì)引起氧化應(yīng)激,導(dǎo)致正常的腎上皮細(xì)胞功能障礙、細(xì)胞凋亡甚至死亡[22]。目前主要凋亡途徑包括線粒體和溶酶體途徑[23],ROS 是ΔΨm 降低的直接原因[24]。當(dāng)細(xì)胞受損時(shí),造成大量ROS 釋放可引起線粒體功能障礙,包括ΔΨm 下降、bcl-2 的降低以及bcl-2 相關(guān)bax 蛋白的釋放,同時(shí)線粒體通過釋放細(xì)胞色素c進(jìn)一步活化Caspase-9,Caspase-9 又可激活下游的Caspase-3,導(dǎo)致細(xì)胞凋亡[25]。本研究通過流式細(xì)胞儀檢測(cè)HAP 可導(dǎo)致ΔΨm 明顯降低、Caspase-3 水平增加,與ROS 的上升趨勢(shì)一致。
溶酶體膜的完整性是細(xì)胞凋亡過程中的關(guān)鍵因素[26],組織蛋白酶B 是一種存在于溶酶體內(nèi)的凋亡介質(zhì)[27],當(dāng)溶酶體膜完整性受損可導(dǎo)致組織蛋白酶B 釋放,從而導(dǎo)致細(xì)胞調(diào)亡。本研究通過共聚焦顯微鏡觀察HAP 主要分布在溶酶體內(nèi),溶酶體膜的通透性增加,組織蛋白酶B 釋放,導(dǎo)致細(xì)胞凋亡。
綜上,本研究對(duì)不同濃度的HAP 通過溶酶體-線粒體途徑誘導(dǎo)HK-2 細(xì)胞凋亡。其機(jī)制:(1)HK-2 細(xì)胞內(nèi)的ROS 釋放增加導(dǎo)致線粒體膜電位降低,激活caspase-3 導(dǎo)致HK-2 細(xì)胞凋亡;(2)HAP主要定位于HK-2 細(xì)胞的溶酶體中導(dǎo)致溶酶體膜的通透性增加,凋亡因子釋放(組織蛋白酶B)引起細(xì)胞凋亡;(3)值得注意的是HAP 作用于HK-2細(xì)胞6 h 時(shí)沒有引起細(xì)胞凋亡,但有大量ROS 產(chǎn)生、線粒體膜電位降低、caspase-3 水平升高等改變。表明凋亡的因素的出現(xiàn)早于細(xì)胞的凋亡。本研究表明HAP 能明顯的抑制HK-2 細(xì)胞增殖,并通過溶酶體-線粒體通徑介導(dǎo)細(xì)胞的凋亡和壞死,從增加了結(jié)石形成的風(fēng)險(xiǎn)。
猜你喜歡
中國(guó)設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2021年6期)2021-11-22 07:50:58
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2020年12期)2021-01-18 06:57:46
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:36
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年9期)2019-11-25 07:34:34
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:50
中學(xué)生數(shù)理化·七年級(jí)數(shù)學(xué)人教版(2019年12期)2019-05-21 02:53:48
