不同時間點針刺對穴區HSP27、MAP4的影響*
2020-08-01 07:45:16白艾靈趙征宇蔡定均
世界科學技術-中醫藥現代化 2020年5期
韋 婷,白艾靈,王 宇,劉 祎,趙征宇,蔡定均
(成都中醫藥大學針灸推拿學院 成都 610037)
針灸治療疼痛療效肯定,其鎮痛效果已經得到了世界公認[1-2]。據美國自然神經科學雜志[3]報道,針灸可以促使穴位局部釋放一些天然的止痛物質,最終達到鎮痛的目的。針刺鎮痛具有明顯的時間差異特性,中醫的傳統時間針灸方法,如子午流注針法、靈龜八法用于痛證治療已取得良好療效[4-5]。現代研究認為,針灸鎮痛的時間效應與5-羥色胺和去甲腎上腺素[6]、香草酸瞬時電位受體4[7]、P2X3 受體[8]及穴區嘌呤信號[9]等有關。但目前尚缺乏細胞骨架相關分子對不同時間點針刺鎮痛效應影響的研究。針刺可以使穴區局部微環境發生改變,引起局部神經感受器的興奮、細胞功能的激活、各種相關化學物質的釋放以及其相互作用形成針刺局部反應小網絡,發揮針刺鎮痛等效應,進而調節機體失衡狀態[10-12]。成纖維細胞作為皮下結締組織中主要的感受性和效應性細胞,在針刺力學轉化過程中起著至關重要的作用。此外,成纖維細胞具有獨立振蕩作用的外周生物鐘,該自主生物鐘對肌動蛋白細胞骨架施加節律性調節,驅動自身節律的運行[13-14]。本課題組前期在針刺鎮痛時間效應的研究中發現,不同時間點針刺對大鼠佐劑性關節炎痛閾的影響不同,在ZT12 和ZT16 時間點針刺均具有明顯的鎮痛效應,且該效應具有時間差異[9]。進一步研究發現,這種鎮痛時間效應差異的產生與針刺調整穴區局部成纖維細胞骨架蛋白有關,不同時間點針刺對成纖維細胞骨架重構的作用以及對其蛋白表達量的影響不同,其中在ZT12和ZT16時間點,成纖維細胞骨架形態改變具有顯著差異[15]。那么針刺鎮痛的不同時間效應是如何影響成纖維細胞骨架重構的,仍有待進一步研究。
引起成纖維細胞骨架重構中間環節的力-化學信號轉導過程受多種蛋白的調控。其中熱休克蛋白27(heat shock protein,HSP27),不僅參與細胞的增殖、分化和細胞凋亡等信號轉導的調節,同時具有保護細胞免受各種應激因素損傷,促進蛋白質正確折疊的功能[16-17]。此外,微管相關蛋白4(microtubule associated protein 4,MAP4)是一種廣泛表達于全身組織的微管動力學調節蛋白,其能促進微管聚合,維持細胞骨架和連接結構的完整,在維持微管穩定方面起著重要的作用[18]。基于此,本研究選擇時間針灸效應明確的痛證為載體,擬觀察在ZT12、ZT16 兩個時間點針刺對成纖維細胞骨架調控相關蛋白分子的變化,從而探討不同時間針刺對成纖維細胞骨架調控的相關原理,以期揭示針刺時間效應的始動機制,為擇時針刺鎮痛提供理論依據。
1 材料與方法
1.1 動物與分組
清潔級雄性SD 大鼠,7-8 周齡,體質量200 ± 20 g,四川省醫學科學院實驗動物研究所提供,許可證號:SCXK(川)2013-15。于成都中醫藥大學時間生物醫學節律室的隔離單元內進行大鼠馴化。自動光暗控制,7:00-19:00 為光照期,室溫22℃±1℃,相對濕度50%-60%,每隔1 小時自動通風1 次。經1 周馴化后,選取光暗周期同步的大鼠,采用疼痛甩尾測試儀進行痛閾基礎值的測定和篩選,納入篩選標準:在3-10 s之間的輻射熱可引起甩尾反應者。然后將篩選留用的36 只大鼠,根據痛閾值完全隨機分為造模組24 只,空白組12 只。造模成功的大鼠以痛閾值隨機分為針刺組和模型組,每組12只。根據針刺時間點的不同又分為ZT12(19:00)、ZT16(23:00)(ZT0 開燈時間7:00,ZT12關燈時間19:00)2個時間亞組。每個對應時間點各18只,隨機為空白組、模型組和針刺組,每組6只。
1.2 主要試劑與設備
完全弗氏佐劑(CFA,批號:101M8711,美國Sigma公司);BCA 蛋白濃度測定試劑盒(批號:KGP903,南京凱基生物公司);預染蛋白Marker(批號:00215343,美國NEB公司);ECL發光試劑盒(批號:BL520A,美國Thermo 公司);Anti-HSP27 抗體(批號:ab109376)、Anti-MAP4 抗體(批號:ab232947)、Anti-β-actin 抗體(批號:ab8227)和山羊抗兔IgG(H+L)(批號:ab6721)均由英國abcam 公司生產。疼痛甩尾測試儀(型號:7360 型,UGOBASILES. R. L,意 大 利Biological Research Apparatus);DYY-6C 電 泳 儀、ChemiDoc XRS+Systems 凝膠掃描成像儀以及化學發光凝膠成像儀均由美國BIO-RAD 公司提供;TGL-16G-A 型高速低溫離心機由上海安亭科學儀器廠生產。
1.3 模型復制
在ZT12 和ZT16 兩個時間點進行模型復制,造模組24 只大鼠在右側足墊部皮內注射CFA 0.1 ml,每個時間點各12只;空白組12只大鼠注射生理鹽水0.1 ml,每個時間點各6 只。24 h 后,造模組大鼠右后足爪明顯腫脹,活動受限跛行,輻射熱甩尾測定大鼠痛閾潛伏期縮短,表明造模成功。
1.4 治療方法
采用自制大鼠固定器固定大鼠,在ZT12、ZT16 對針刺組大鼠進行針刺治療,選擇大鼠右側“足三里”穴(ST36),消毒后直刺7 mm,留針30 min,每5 min 采用捻轉平補平瀉法行針1次,每次1 min,頻率120次/min,1 次/d,治療7 d。對兩個時間點的其余組大鼠用相同方法抓取及固定30 min。
1.5 觀察指標及檢測方法
免 疫 印 跡 法(Western blot)測 定 穴 區HSP27、MAP4 蛋白表達:針刺治療結束后,股動脈放血法處死大鼠,提取大鼠患側足三里局部的皮膚和皮下組織,裂解液裂解10 min,12 000 r/min 離心10 min,取上清液,BCA法測定樣品蛋白濃度,經蛋白定量后,煮沸10 min變性蛋白,電泳,轉膜,切膜,TBS 浸濕膜后,封閉1 h,膜上滴加一抗,4℃過夜,滴加二抗,孵育2 h,經ECL化學發光法檢測蛋白條帶。結果以目的蛋白相對表達量表示(目的蛋白相對表達量=目的蛋白積分光密度值(IOD)/內參積分光密度值(IOD)。
1.6 統計學處理
采用統計軟件SPSS 21.0進行數據分析,數據以均數±標準差(xˉ±s)來描述,經過正態分布和方差齊性檢驗后,各組間比較用單因素方差分析,方差齊者采用LSD 法,方差不齊者采用Tamhane's T2 法,以P<0.05表示差異有統計學意義。
2 結果

圖1 ZT12、ZT16兩個時間點各組大鼠穴區皮膚HSP27蛋白表達量
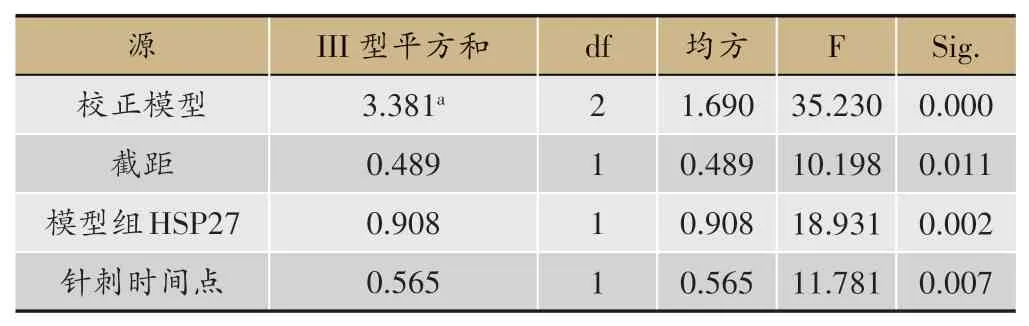
表1 主體間效應的檢驗(因變量:針刺組HSP27)
2.1 不同時間點針刺對大鼠穴區HSP27 蛋白表達量的影響
針刺大鼠足三里7 天后,由圖1 結果所示,相同時間點不同組間比較:在ZT12 時間點,模型組HSP27 蛋白表達量較空白組明顯增高(P<0.05);針刺組HSP27表達量較模型組明顯降低(P<0.05)。在ZT16 時間點,模型組HSP27 表達量較空白組明顯增高(P<0.05)。不同時間點相同組間比較:在空白組與針刺組中,ZT16 時間點HSP27 表達量較ZT12 時間點顯著增高(P<0.01)。為排除不同時間點造模對針刺組HSP27 蛋白表達的混雜影響,故將兩個時間點模型組的HSP27 蛋白表達量納入協變量,運用協方差分析不同時間針刺對HSP27 蛋白的調整力度。如表1 和圖2所示,不同時間點造模對針刺組HSP27 蛋白表達具有顯著的影響(P<0.01),在排除不同時間點造模對針刺調節作用的干擾后,在兩個不同時間點針刺對HSP27 蛋白表達量的影響仍然存在顯著的差異(P<0.01),且ZT16 時間針刺對該蛋白的調整幅度大于ZT12時間點。

圖2 不同時間點針刺對HSP27的調整幅度

圖3 ZT12、ZT16兩個時間點各組大鼠穴區皮膚MAP4蛋白表達量
2.2 不同時間點針刺對大鼠穴區MAP4 蛋白表達量的影響
針刺治療7 天后,由圖3 結果所示,相同時間點不同組間比較:在ZT12 時間點,模型組大鼠穴區MAP4表達量較空白組明顯增高(P<0.05);不同時間點相同組間比較:空白組ZT16 時間點大鼠穴區MAP4 表達量較ZT12 時間點明顯增高(P<0.05);針刺組大鼠穴區MAP4 表達量ZT16 時間點較ZT12 時間點明顯增高(P<0.05)。為排除不同時間點造模對針刺組MAP4蛋白表達的混雜影響,將兩個時間點模型組的MAP4蛋白表達量納入協變量,運用協方差分析不同時間針刺對MAP4蛋白的調整力度。如表2和圖4所示,不同時間點造模對針刺組MAP4蛋白表達具有顯著的影響(P<0.01),在排除不同時間點造模對針刺調節作用的干擾后,在兩個不同時間點針刺對MAP4 蛋白表達量的影響仍然存在明顯的差異(P<0.05),且ZT16 時間點針刺對MAP4 蛋白表達量的調整幅度大于ZT12時間點。

表2 主體間效應的檢驗(因變量:針刺組MAP4)

圖4 不同時間點針刺對MAP4的調整幅度
3 討論
HSP27 蛋白是HSP 亞家族中的重要一員,在生物體內參與多種復雜的功能活動,并反映出不同的生物學效應,能夠調節多種信號轉導通路,具有細胞保護作用。磷酸化的HSP27 可能介導血管緊張素II 與血小板衍生生長因子誘導的血管平滑肌細胞的骨架重建和遷移,并且具有促進微絲形成和穩定黏著斑的作用,這對促進細胞遷移有著重要作用[19-20]。有研究發現,在急性胰腺炎早期,氧化應激損傷降低了細胞骨架的穩定性,同時HSP27 蛋白及其磷酸化蛋白被激活,從而發揮對胰腺細胞骨架穩定性的保護作用[21]。各種對細胞有害的應激環境能夠誘導細胞骨架的重組或破壞,p38 絲裂原活化蛋白激酶(p38MAPK)信號轉導通路使HSP27 發生磷酸化,磷酸化的HSP27 參與調控細胞骨架重組,穩定細胞骨架結構[22]。也有研究認為,HSP27 的磷酸化主要受蛋白激酶D(PKD)的激活,從而發揮對缺血性損傷后的神經保護作用[23]。本課題組研究發現,在ZT12 和ZT16 時間點復制佐劑型關節炎疼痛模型,導致這兩個時間點大鼠穴區皮膚中HSP27 蛋白表達應激性增加,針刺治療后能明顯降低ZT12 時間點HSP27 蛋白表達;在不同時間點針刺對HSP27的蛋白調整作用不同,其中在ZT16時間點針刺對該蛋白表達的調整幅度大于ZT12時間點。
MAP4 是微管相關蛋白的一種,具有調節微管排列、促進微管組裝、交聯微管和肌動蛋白等生理功能。MAP4 包被在微管外部時,微管蛋白亞單位不能脫離微管的末端,從而起到了穩定微管的作用。MAP4 既可以獨立引起微管解聚,又能協同相關蛋白通道共同作用于微管,影響細胞骨架重構。體內外研究均顯示MAP4 能增強微管蛋白聚合和整體微管的穩定性[24]。MAP4主要通過兩種途徑促進微管的穩定性:①通過捆綁微管并促進對外力的抵抗;②通過結合微管和空間阻斷主動分解微管的蛋白質[25]。MAP4 的磷酸化誘導MAP4從微管中脫離,使微管細胞骨架失穩。缺氧導致心肌細胞微管結構遭到破壞并激活p38MAPK 信號傳導通路,使MAP4磷酸化增加,從而誘導微管細胞骨架解聚并降低細胞活性[26-27]。炎性條件下,肺內皮屏障功能障礙使MAP4 磷酸化伴隨p38MAPK 信號途徑的激活而增加,從而誘導微管解聚,使肺血管通透性增高[18]。本課題組的研究結果發現,在ZT12和ZT16時間點復制佐劑型關節炎疼痛模型,導致ZT12時間點大鼠穴區皮膚中MAP4 蛋白表達應激性增加;在不同時間點針刺對MAP4 的蛋白調整作用不同,其中在ZT16 時間點針刺對該蛋白表達的調整幅度大于ZT12時間點。
HSP27 和MAP4 在細胞骨架的調控過程中發揮至關重要的作用。細胞化學信號的產生依賴于細胞骨架將機械信號傳導至細胞核,成纖維細胞作為應力細胞,能感知和傳導機械力刺激,將其轉化為力學信號從而導致細胞產生一系列生物效應[28-29]。成纖維細胞的這種信號轉導作用得力于其細胞骨架的感受器作用,細胞骨架是一組由纖維結構組成的蛋白網狀支架,由微絲、微管及中間絲構成。機械力作用于成纖維細胞引起的細胞骨架重構及形態變化參與信號傳導。針刺作用屬于物理力學刺激的一種,能夠將刺激作用轉換為生物化學信號,對成纖維細胞骨架、形態以及其相關生長因子進行調節[30-31]。已有研究表明,針刺對成纖維細胞骨架重構及其骨架蛋白具有明確的調整作用,且這種效應具有時間差異[15]。本研究在前期不同時間點針刺對鎮痛效應不同的研究基礎上[7-9,15],選擇針灸鎮痛效應具有明顯差異的兩個時間點,即ZT12和ZT16時間點,通過比較這兩個時間點針刺對穴區皮膚中HSP27 和MAP4 蛋白表達量的差異,探討其對成纖維細胞骨架調控的相關原理,以期揭示針刺時間效應的始動機制,為針刺鎮痛的擇時治療提供依據。本研究發現,在ZT12和ZT16兩個時間點,針刺在相同時間點不同組間以及不同時間點相同組間,對HSP27 和MAP4 蛋白表達均具有不同的調節作用,并且在ZT16時間點,針刺對HSP27、MAP4蛋白表達量的調整作用均大于ZT12時間點。因此本課題組推測,針刺通過對穴區HSP27、MAP4 蛋白表達的調整,從而影響成纖維細胞骨架重構的改變,在不同時間點ZT12、ZT16針刺對穴區成纖維細胞骨架重構的不同調控效應,可能與這兩個時間點針刺對骨架蛋白表達相關信號分子HSP27、MAP4 的調整作用不同有關,從而發揮不同的鎮痛效應。
猜你喜歡
核科學與工程(2021年4期)2022-01-12 06:30:26
中國民間療法(2021年1期)2021-04-20 02:30:34
基層中醫藥(2021年7期)2021-03-26 18:00:27
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
中學物理·高中(2016年12期)2017-04-22 11:53:03
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
中醫研究(2014年8期)2014-03-11 20:29:23
中醫研究(2014年6期)2014-03-11 20:29:02
中國中醫藥現代遠程教育(2014年17期)2014-03-01 04:29:28
