不同譜系節節麥的雜種優勢分析
2020-08-03 06:04:46趙夢雨劉頡堯宋思宇董浩然李玉閣李鎖平
麥類作物學報 2020年2期
蘇 晴,趙夢雨,劉頡堯,宋思宇,吳 瀟,蔣 政,董浩然,李玉閣,李鎖平
(河南大學生命科學學院,河南開封 475004)
利用雜種優勢提高作物產量、品質、適應性和抗逆性,是作物遺傳改良的重要途徑。雜交小麥是今后大幅度提高小麥產量的首選途徑[1]。小麥的遺傳基礎日益狹窄,限制了其雜種優勢利用和遺傳改良[2-3]。從野生近緣物種中發掘有益基因資源并將其導入普通小麥,是當前通過雜交育種改良普通小麥的主要途徑[4-5]。節節麥(AegilopstauschiiCoss.2n=2x=14,DD)是禾本科(Poaceae)小麥族(Triticeae)山羊草屬(Aegilops)的一年生草本植物,是公認的普通六倍體小麥(TriticuvumaestivumL. 2n=6x=42,AABBDD)D基因組的二倍體供體[6]。根據穗型等形態學特征,Hammer[7]將節節麥劃分為 ssp.tauschii和ssp.strangulala兩個亞種。根據其生態習性,節節麥可分為雜草型和野生型;前者通常以田間雜草的形式分布于小麥主產區的田間地頭,后者則通常以自然群落形式存在于伊朗、阿富汗南部、巴基斯坦和我國新疆伊犁地區[8]。Mizuno等[9]、Sohail等[10]、Wang等[11]和Singh等[12]分別采用AFLP、DArT、SNPs分子標記技術,根據節節麥的種群遺傳結構特征,將世界分布的節節麥清晰地劃分為L1(lineage 1)和L2(lineage 2)兩大譜系。L1譜系主要是土耳其、薩爾維亞、阿富汗和中國黃淮麥區(河南、陜西)等東部分布區的雜草型節節麥,L2譜系則主要是伊朗、阿富汗南部、巴基斯坦、中國新疆伊犁地區等西部分布區的野生型節節麥。研究表明,僅有少數隸屬于L2譜系、2-3亞系、ssp.strangulala亞種的節節麥參與了普通小麥的形成[9-10]。而具有比普通小麥D基因組更廣泛遺傳變異而沒有參與普通小麥起源的L1譜系節節麥,可能是對小麥遺傳改良有重要應用價值的寶貴野生種質資源[13]。目前,已從節節麥中發掘出大量優質、抗病、抗逆等對普通小麥遺傳改良有應用價值的有益基因,部分基因已通過直接或間接途徑導入普通小麥[14-23]。
在地理分布上存在隔離的兩大譜系節節麥,在遺傳組成上也存在明顯的遺傳分化,很少存在中間過度類型的節節麥[9]。說明兩個譜系的節節麥在自然條件下是隔離生殖的[11],對L1和L2譜系節節麥是否已形成遺傳學意義上的生殖隔離、是否存在顯著的雜種優勢,目前尚不明確。
本研究擬對L1和L2譜系節節麥進行人工雜交,結合SSR分子標記、花粉活力測定、染色體減數分裂的配對情況及其16個主要農藝性狀的調查,比較L1和L2譜系節節麥的遺傳差異和雜種優勢,為其在普通小麥育種中的雜種優勢利用提供參考。
1 材料與方法
1.1 試驗材料
本研究采用的國外節節麥由美國國家植物種質中心提供,中國河南、新疆的節節麥則由河南大學植物種質資源與遺傳實驗室采集并保存。通過預試驗,配置成功并種植的14個節節麥雜交組合的材料來源信息見表1。
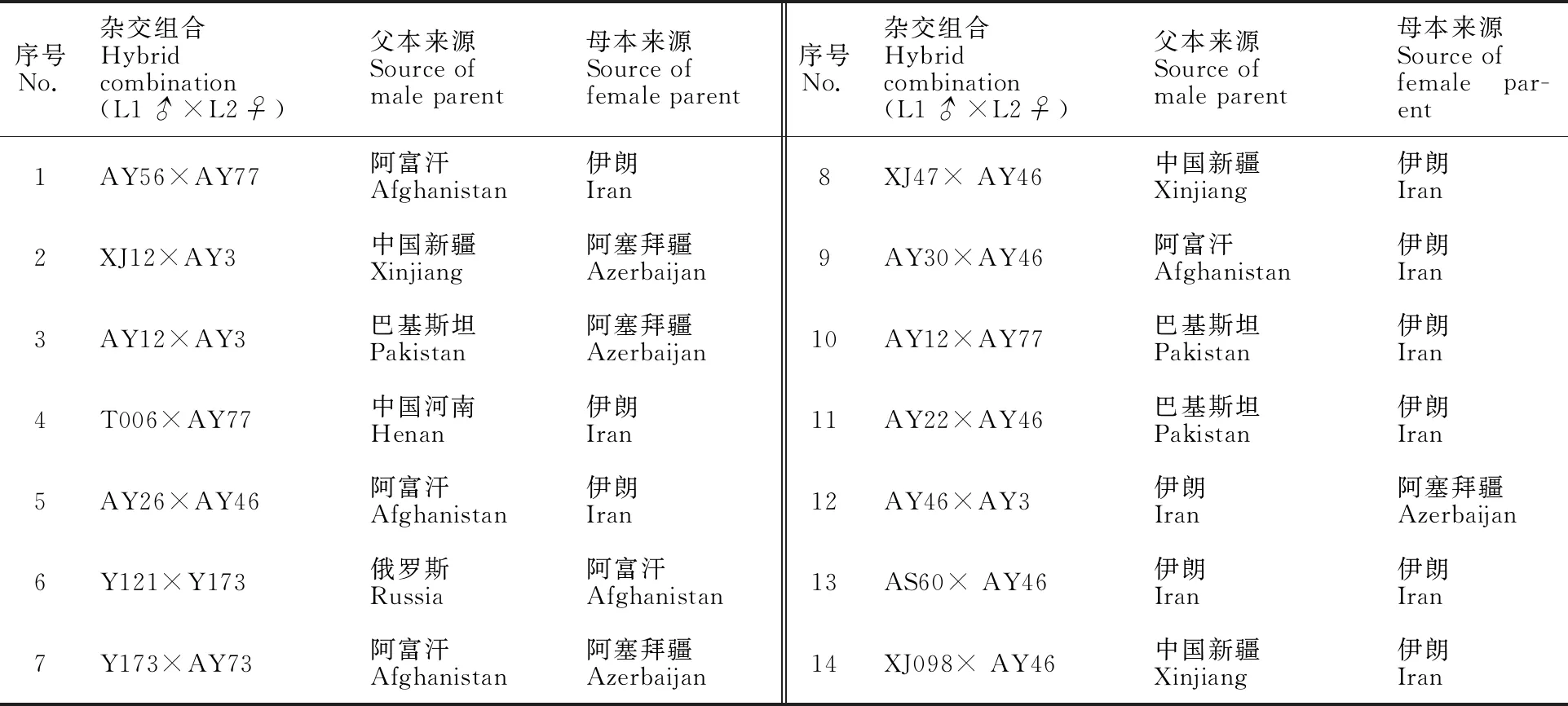
表1 配置雜交組合、親本節節麥編號及來源信息表
1.2 試驗方法
1.2.1 材料種植
采用完全隨機區組設計,每個雜交組合3次重復,每個重復包含父本、雜交種和母本各兩行,每行5株(取材及調查性狀選取中間3株),行距為40 cm,株距為25 cm。人工播種,田間管理按照當地常規生產方式進行。
1.2.2 雜種F1真實性鑒定
取5~8葉齡的節節麥葉片(父本和母本各取一株葉片,所有雜交F1均要取材)用于CTAB法提取親本和子代F1的基因組DNA,選用8對分別位于節節麥不同染色體上的SSR引物(表2),通過PCR擴增、聚丙烯酰胺凝膠電泳[24]、親本和子代F1電泳條帶的組成分析,鑒定雜種F1的真實性。
PCR擴增采用10 μL的反應體系,包含 1 μL 10×PCR緩沖液(Mg2+),1 μL dNTPs (2.5 mmol·L-1),正反向引物(10 μmol·L-1)各0.5 μL,0.1 μLTaqE(5 U· μL-1),2 μL DNA模板(30 ng· μL-1),4.9 μL雙蒸水。PCR反應程序為:95 ℃變性3 min;95 ℃變性30 s,退火45 s(Tm見表2),72 ℃延伸1 min,35個循環;72 ℃保持 15 min。

表2 鑒定雜種的SSR引物序列
根據鑒定結果,拔除假雜種。
1.2.3 花粉育性的觀察和統計
取親本和雜種F1當天開花的3個小穗,分別取上、中、下部花藥,擠出花粉,采用K2-KI溶液染色法進行染色制片和顯微鏡觀察。根據花粉粒染色后的著色深淺、花粉粒的飽滿度等特征辨別花粉的育性[25],取顯微鏡下的3~5視野的花粉進行觀察和計數,求平均值獲得花粉活力的百分比,挑取典型的視野拍照留存。
1.2.4 染色體配對情況的觀察和統計
從14個雜交組合中隨機挑選8個用于同源染色體配對觀察。選取3~5個雜種F1減數分裂時期的小穗,將花藥剝離并用FAA(醋酸∶酒精=1∶3)固定24 h;通過醋酸洋紅染色后制片,顯微鏡觀察,統計并記錄F1減數分裂時同源染色體的配對情況,對典型圖片進行拍照。
1.2.5 主要農藝性狀的調查
通過定性觀察,篩選8個F1生長勢較優的雜交組合進行主要農藝性狀調查。每個重復挑選長勢一致的3株,分別對兩親本(P1,P2)和F1的16個農藝性狀進行田間調查。用中親優勢指數 (index of heterosis,IH) 和超親(高值親本,HP)優勢指數(over-parent heterosis,OPH)衡量L1和L2譜系節節麥的雜種優勢。
IH=[F1-(P1+P2)/2]/[(P1+P2)/2]×100%
OPH=(F1-HP)/HP×100%
1.3 數據處理
采用單因素方差分析對F1與親本的差異進行顯著性檢驗,并通過LSD法進行多重比較。
2 結果與分析
2.1 雜種F1真實性的SSR分子標記分析
通過對14個雜交組合父母親本及其F1進行SSR分子標記檢測,發現所有F1條帶都兼具兩親本的共有帶型,說明F1是真實雜交種。圖1展示了部分雜交組合的親本及其雜交F1的SSR檢測結果。
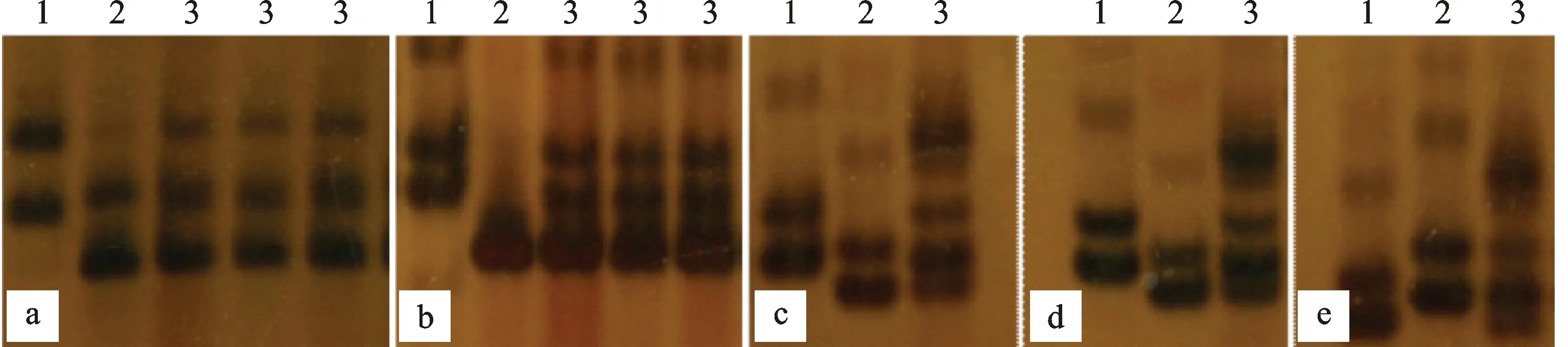
a、b、c、d和e分別代表編號為13、5、1、3和4雜交組合材料。1:父本;2:母本;3:F1。a和b 的引物分別為Xgpw4024和Xcfd132,c~e的引物均為Xgdm61。
2.2 雜種F1代的花粉育性分析
對14個雜交組合的花粉活力進行統計分析,結果(表3)表明,不同節節麥的花粉活力存在較大差異,AS60的花粉活力最高,有活力花粉比例高達99.231%,而AY22的花粉活力較低,有活力花粉占比78.892%。親本花粉活力較高的雜交組合,如AY12×AY77,其F1花粉活力也較高(圖2),說明雜種F1的花粉活力與父母本的花粉活力有關。14個雜交組合F1的花粉活力均與中親值相差不大,育性正常。

A:AY12;B:AY77; C:雜種F1。箭頭指示的是無活力的花粉。
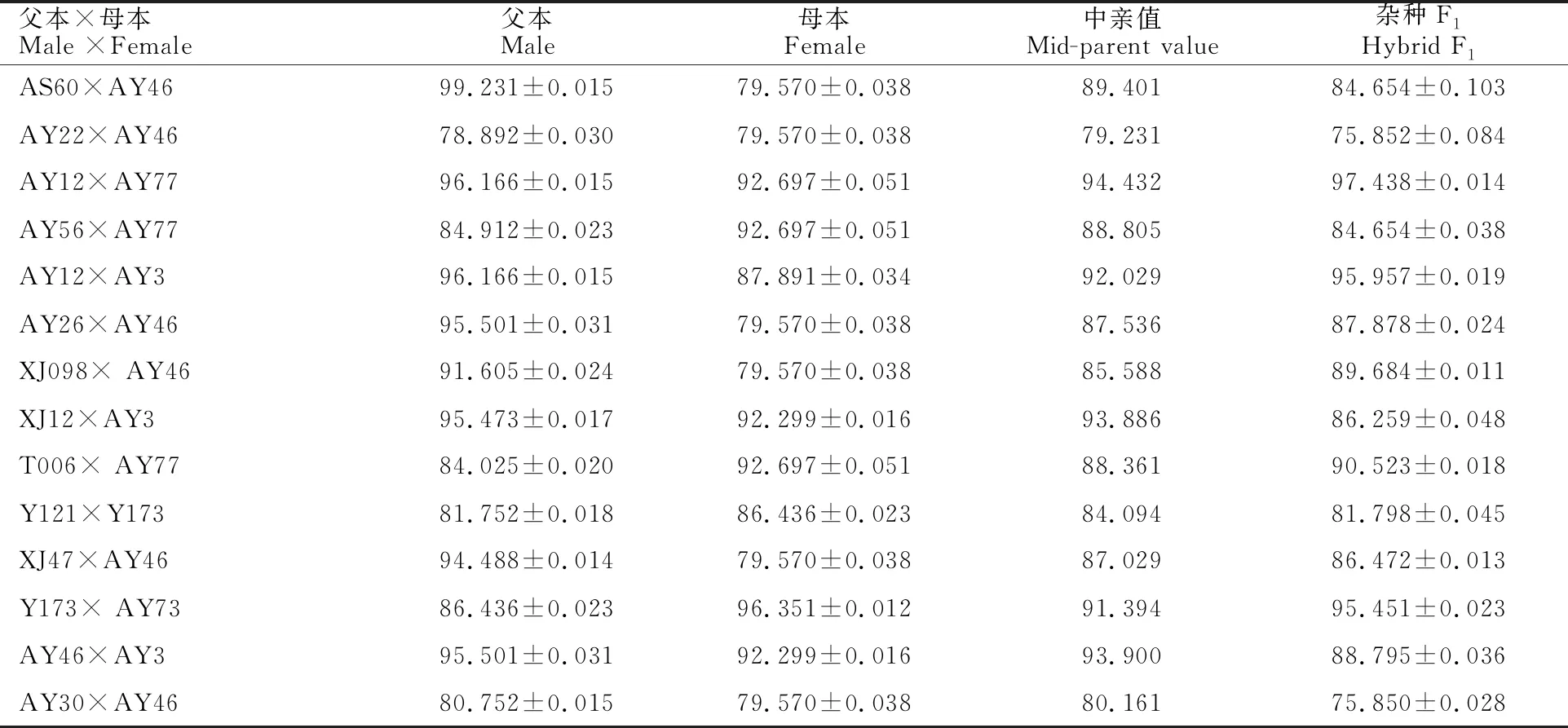
表3 14個雜交組合的花粉活力
2.3 雜種F1代同源染色體的配對統計結果
從14個雜交組合中隨機選取8個雜交組合,對其F1減數分裂時期的同源染色體配對情況進行觀察和統計分析,結果(表4、圖3)表明,雜交組合AY12×AY77、XJ098×AY46的F1減數分裂中期染色體均為7對環狀二價體,其他雜交組合的雜交F1代,減數分離期大多數同源染色體配對形成環狀二價體,個別配對時形成1~2個棒狀二價體。說明L1和L2譜系節節麥在染色體水平上雖有一定的遺傳分化,但同源染色體同源率較高,能正常配對,產生可育的后代。

圖3 雜交組合AY26×AY46的F1同源染色體配對結果圖

表4 不同雜交組合中雜種F1的染色體配對情況統計結果
2.4 主要農藝性狀的雜種優勢分析
在定性觀察的基礎上,對8個F1生長勢較優的雜交組合的產量(分蘗數、穗長、小穗數、粒重、籽粒結實率)、花期(抽穗期、開花期)、株型(株高、旗葉長、旗葉寬)、籽粒(穎殼長、穎殼寬、外稃長、外稃寬、內稃長、內稃寬)等相關農藝性狀進行調查統計發現,L1和L2譜系的節節麥在分蘗數(表5)和粒重(表6)上表現出明顯的雜種優勢,而其 他性狀在父母本和F1間差異不顯著(數據未 列出)。
8個雜交組合的分蘗數均有良好的雜種優勢,中親優勢指數(IH)為120.31%~ 223.73%,超親優勢指數(OPH)3.97%~ 86.76%。有4個組合的百粒重(表6)具有明顯雜種優勢,中親優勢指數(IH)在143.62%~ 147.71%,超高值親本優勢(OPH)在21.71%~28.80%。3個雜交組合(AY56×AY77、AY26×AY46和XJ47×AY46)的百粒重(IH>143%,OPH>21%)和分蘗數(IH>187%,OPH>57%)均存在明顯的雜種優勢。特別是AY26×AY46(圖4)和XJ47×AY46(圖5)在整株生長勢上,雜種F1均明顯優于其父母本。對上述8個雜交組合的百粒重和分蘗數進行多重比較發現(表5,表6),雜種F1的分蘗數均顯著高于親本。

圖5 雜交組合XJ47×AY46的整株表型比較圖

表5 節節麥雜交組合的分蘗數雜種優勢比較
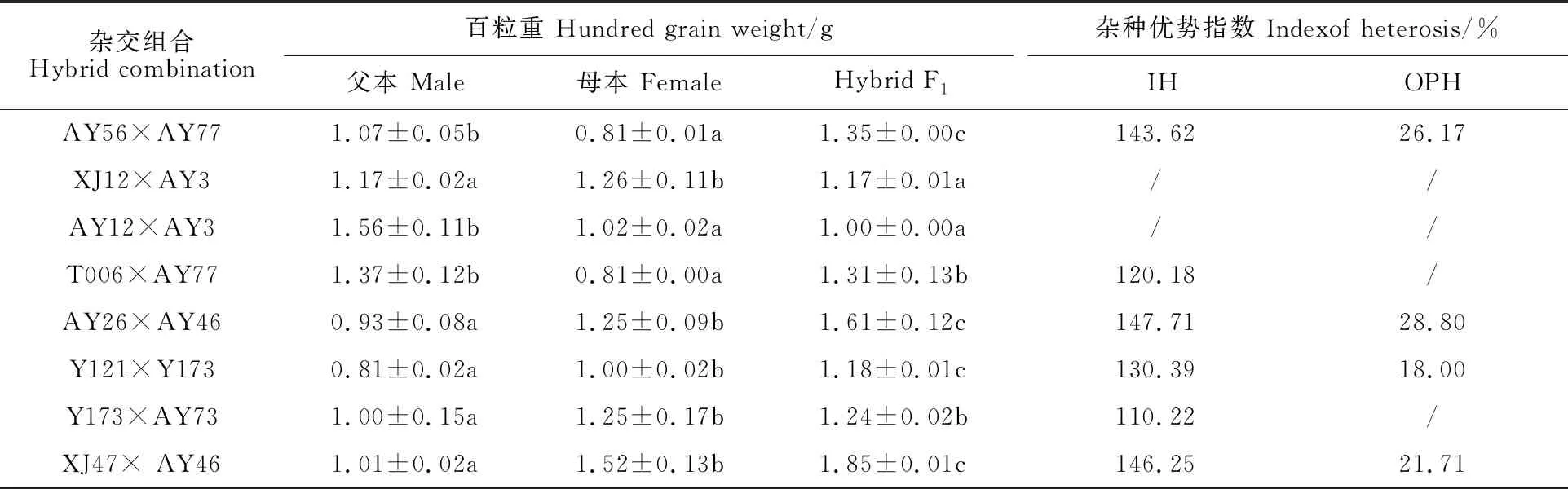
表6 8個節節麥雜交組合的粒重雜種優勢比較

圖4 雜交組合AY26×AY46的整株表型比較圖
3 討 論
作為普通小麥D染色體的供體,節節麥是小麥育種中重要的基因資源庫,節節麥與普通小麥的D基因組的染色體可發生自由交換和重組,染色體重組后幾乎無負遺傳作用,這些為節節麥優勢基因導入普通小麥提供了便利和可行性[26]。目前已在節節麥中篩選并分離出大量抗白粉病[14-15]、小麥癭蚊[16]、葉銹病[17]、稈繡病[18]、條銹病[19]、麥二叉蚜[20]、穗發芽[21]和耐鹽、優質高分子量谷蛋白亞基[22]、耐旱[23]等有益基因,并通過普通小麥與節節麥雜交的直接轉育途徑,或節節麥與四倍體小麥雜交構建橋梁人工親本六倍體小麥,再與普通小麥雜交的間接轉育途徑,已成功對小麥進行了遺傳改良。
節節麥是小麥育種可利用的重要基因資源,世界地理分布清晰的形成了分別代表東、西節節麥分布區的L1和L2兩大譜系[9-11],中間沒有過度類型,說明兩個譜系節節麥在自然狀態下是生殖隔離的。為揭示L1和L2譜系節節麥是否已形成遺傳學意義上的生殖隔離、是否存在顯著的雜交優勢,本研究通過對14個L1和L2譜系節節麥及其雜交組合的花粉活力鑒定、染色體配對情況觀察和16個農藝性狀的統計分析發現,雜交F1的花粉育性正常,花粉母細胞在減數分裂時大多數同源染色體能正常配對形成7對環狀的二價體,少數出現棒狀染色體,說明L1和L2譜系節節麥間雖有明顯的遺傳分化,但并未產生遺傳學意義上的生殖隔離。該結果與孔令讓等[26]對粗山羊草與普通小麥的屬間雜交研究結果類似。
對八個雜交組合的16個農藝性狀調查分析表明,L1和L2譜系節節麥間在粒重和分蘗數上存在顯著雜交優勢,推測節節麥對提高小麥產量有重要的應用潛力。劉 磊等[27]通過對大穗、大粒節節麥AT462和強分蘗節節麥AT18雜交后代性狀的相關性分析結果表明,控制節節麥穗長、小穗數、粒重等產量性狀的基因間加性效應顯著,可應用于小麥的產量育種。
綜上可見,參與小麥進化起源的主要是L2譜系的少部分節節麥[9-10],但存在更廣泛的遺傳變異、沒有參與普通小麥起源的L1譜系節節麥,尤其是與L2譜系有明顯雜種優勢的L1譜系節節麥,是普通小麥遺傳改良可利用的重要種質資源,有望通過直接或間接轉育途徑,用于小麥產量的遺傳改良。
