低溫對富士蘋果花抗性生理指標的影響
2020-08-04 12:30:35姜琳琳王靜段曉鳳
江蘇農業科學 2020年11期
姜琳琳 王靜 段曉鳳

摘要:以富士蘋果花為研究對象,采用霜凍模擬試驗箱模擬自然霜凍降溫過程,分別測定-2、-2.5、-3、-3.5、-4、-4.5 ℃時蘋果花的丙二醛(MDA)、脯氨酸、可溶性蛋白質和可溶性糖含量。結果表明,隨溫度的降低,蘋果花可溶性糖含量在-2.5 ℃時顯著增加,達到峰值后其含量開始下降并趨于穩定;可溶性蛋白含量在-2.5 ℃時顯著增加,在-3.5~-4 ℃時達到峰值后顯著降低;脯氨酸含量在-3~-3.5 ℃時顯著增加;MDA含量在-3~3.5 ℃時達到峰值后顯著降低。總體來說,上述抗性生理指標對低溫響應表現出較好的一致性,初步確定蘋果花對低溫響應的臨界溫度為-2.5~-3.5 ℃。
關鍵詞:低溫;蘋果花;可溶性糖;丙二醛(MDA);脯氨酸;可溶性蛋白
中圖分類號: S661.101 ?文獻標志碼: A ?文章編號:1002-1302(2020)11-0111-04
收稿日期:2019-07-06
基金項目:國家自然科學基金地區科學基金(編號:41565006)。
作者簡介:姜琳琳(1992—),女,河南許昌人,碩士,主要從事農業氣象研究。E-mail:15261822625@163.com。
通信作者:李紅英,博士,主要從事氣候資源利用與農業氣象災害研究。E-mail:hongyinglhy@126.com。 ?蘋果產業是寧夏果品的傳統產業,是果農種植時間最早、比較效益相對較高的優勢產業。近年來,隨全國蘋果產業發展由東部向西部的轉移,寧夏成為蘋果生產的重要產區,種植面積已達4.46萬hm2,蘋果產業已成為寧夏地方經濟發展和農民增收致富的重要特色優勢產業[1]。寧夏深處內陸,大陸性氣候特點突出,在每年4月冷暖空氣交匯時期,晚霜凍害頻繁發生,嚴重影響蘋果產量和品質,對蘋果產業的發展造成很大威脅。據寧夏氣象科學研究所調查,2018年4月4—8日出現的霜凍天氣使寧夏地區正值花期的蘋果嚴重受凍。其中,銀川河東生態園藝試驗中心、靈武園藝場、吳忠園藝場和中寧轎子山林場蘋果受凍率分別高達90%、70%、90%和85%。
在蘋果低溫抗寒性方面,前人已經做了大量研究。研究表明,蘋果花器官不同發育階段抗寒力存在差異,即花蕾期>盛花期>幼果期,其中花蕾和花朵的低溫半致死溫度分別為-4.53、-4.01 ℃[2]。許多學者通過測定低溫脅迫下植物細胞內可溶性蛋白、脯氨酸、丙二醛(MDA)和可溶性糖等物質的含量對蘋果的抗寒性進行評價[3-5]。但前人研究多集中于蘋果枝條和果實,對低溫下蘋果花的生理響應研究較少。因此,本研究通過模擬自然霜凍降溫過程,對蘋果花中MDA、脯氨酸、可溶性蛋白質以及可溶性糖含量進行測定,分析蘋果花在低溫下的生理生化響應規律,以期為蘋果霜凍指標的確定和防霜技術的研發提供參考依據。
1 材料與方法
1.1 試驗材料
試驗于2017年4月22—23日在寧夏河東生態園藝試驗中心進行,為了確保試驗結果具有可比性,選取樹齡(13~15年)、樹勢、營養狀況一致的富士蘋果樹作為采樣樹體,采取長度至少30 cm、花量均勻的蘋果枝條,插入濕度適宜、覆膜的營養缽培養待用。
1.2 試驗材料的低溫處理
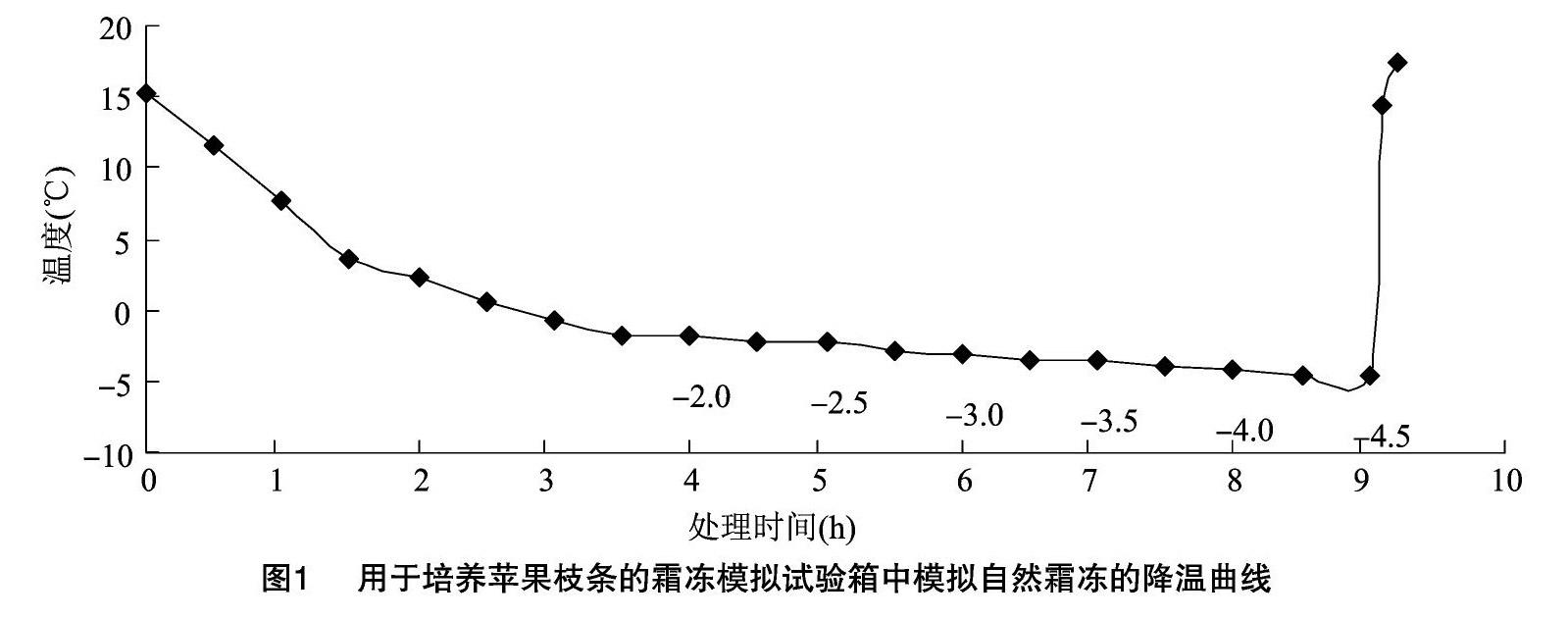
試驗儀器采用中國農業科學院開發的、以PID調節方式控溫的MSX-2F型模擬霜箱系統。將培養的蘋果枝條連同營養缽置于霜凍模擬試驗箱,按照圖1所示設置降溫曲線,試驗處理溫度為-2、-2.5、-3、-3.5、-4、-4.5 ℃,降溫過程中在達到每個處理溫度并持續0.5 h之后取出一批樣品,每批蘋果花樣品為10~15朵,并且為了盡量減少取樣對降溫過程的影響,每次取樣時間嚴格控制在 10 s 之內。以營養缽中室溫培養的樣本作為對照(CK)。低溫處理的每一批樣品要及時用液氮處理保存其生理活性狀態,用于測定各項生理指標,每批樣品重復測定3次。
1.3 測定項目與方法
MDA含量采用硫代巴比妥酸法[6]測定;游離脯氨酸采用磺基水楊酸浸提-酸性茚三酮顯色法[7]測定;可溶性蛋白質含量采用考馬斯亮藍法[7]測定;可溶性糖含量采用蒽酮比色法[7]測定。
1.4 數據處理
采用Excel 2010和SPSS 21.0軟件進行數據處理。
2 結果與分析
2.1 低溫對蘋果花可溶性糖含量的影響
低溫條件下可溶性糖含量越高,抗凍力越強。由圖2可知,蘋果花的可溶性糖含量隨著溫度降低呈現先升高后下降的趨勢。當溫度下降到-2.5 ℃時,蘋果花可溶性糖含量顯著高于-2 ℃時的可溶性糖含量。-3~-2.5 ℃可溶性糖含量維持在較高的水平。在整個研究溫度范圍(-2~-4.5 ℃)內,當可溶性糖含量達到峰值之后開始略微下降并趨于穩定。
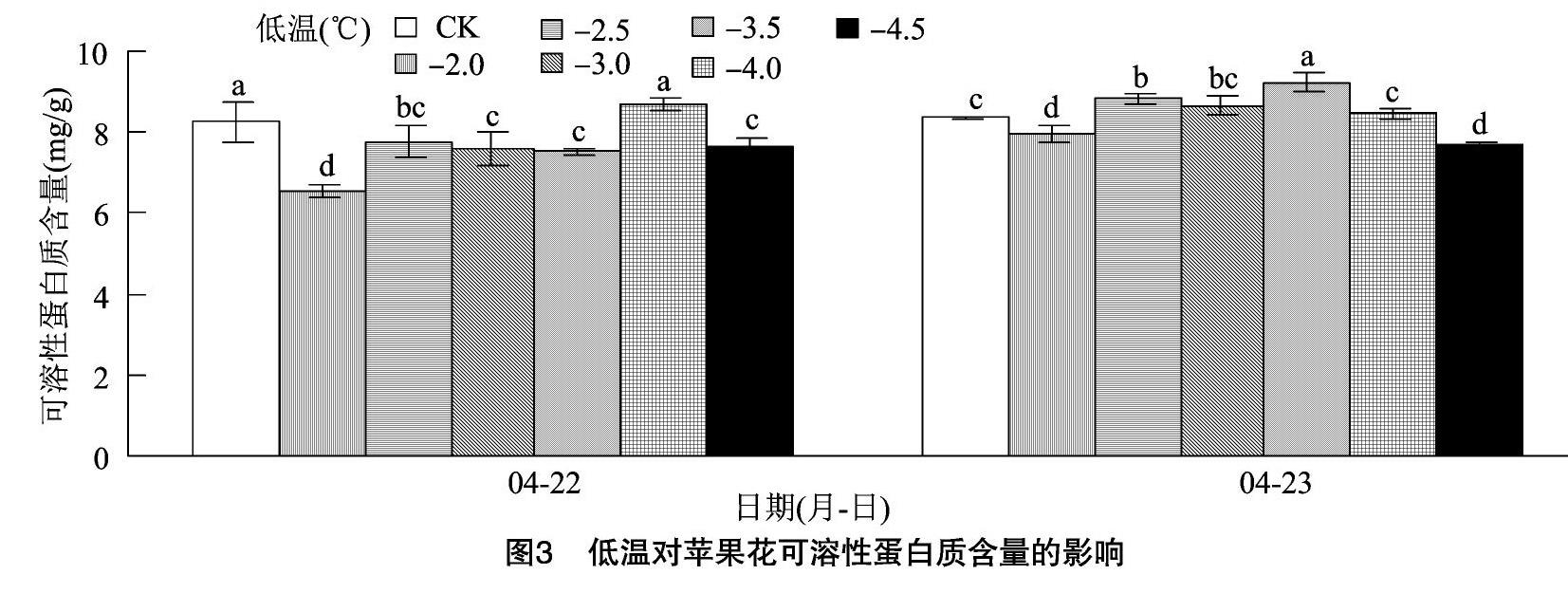
2.2 低溫對蘋果花可溶性蛋白質含量的影響
由圖3可知,隨溫度的降低,蘋果花的可溶性蛋白質含量呈先增加后下降的趨勢。與-2 ℃ 相比,
-2.5 ℃ 時蘋果花可溶性蛋白質含量顯著升高,之后隨著溫度下降其含量繼續上升,在-3.5~-4.0 ℃達到峰值。-4~-2 ℃蘋果花蛋白質含量的增加可能是由于其抗冷性增強導致的,之后其含量的降低可能是由于長期低溫致使蘋果花細胞遭受了不可逆傷害。
2.3 低溫對蘋果花脯氨酸含量的影響
脯氨酸是植物在逆境脅迫下的產物。從圖4可以看出,降溫導致蘋果花脯氨酸含量呈現增加趨勢,4月22、23日,蘋果花的脯氨酸含量分別在-3、-3.5 ℃下顯著高于前一溫度處理,之后其含量也一直顯著高于CK。
2.4 低溫對蘋果花MDA含量的影響
MDA是細胞膜脂過氧化的產物。由圖5可知,隨著溫度的下降,MDA含量呈先上升后下降最后趨于平穩的趨勢。4月22、23日,蘋果花的MDA含量分別在-3.5、-3 ℃達到峰值,分別為6.2、5.8 μmol/g。之后隨溫度繼續降低,MDA含量開始下降,但與隨后一溫度處理相比,其含量無顯著變化。降溫后期蘋果花MDA含量趨于穩定,這可能是由溫度達到了蘋果花所能承受的低溫臨界值,細胞受到不可逆傷害導致的。
3 討論與結論
MDA是細胞膜脂過氧化的產物,逆境脅迫會導致其含量增加[8-9]。本研究表明,其含量在低溫下
呈先上升后下降最后趨于平穩的趨勢。這與閆忠業等對蘋果枝條的研究結果[10]一致,表明當溫度低于蘋果花體受凍臨界值時,蘋果花體將會遭受不可逆的傷害。脯氨酸是植物細胞內分布最廣泛的一種有機滲透調節物質[11],逆境下植物體內脯氨酸含量的增加通常被認為是植物的一種適應性反應[12-13]。也有研究認為,逆境下其含量的增加是植株受傷害的表現[14]。還有研究表明,脯氨酸含量與植株抗寒性沒有明顯的相關性[10,15]。本研究表明,隨溫度的降低,蘋果花的脯氨酸含量呈增加趨勢,結合MDA含量的變化特點,認為其含量的增加是植株受傷害的結果。一般認為,可溶性蛋白質含量與植物的抗寒性相關。有研究表明,低溫下可溶性蛋白質含量呈增加趨勢[16]。也有研究表明,低溫會造成可溶性蛋白質,尤其是高分子可溶性蛋白質含量的迅速下降[14]。本研究表明,低溫下蘋果花的可溶性蛋白質含量呈現先增加后降低的趨勢,這與令凡等的研究結果[17]一致。蛋白質含量增加可能是蘋果花對低溫的適應的表現。可溶性糖是冷害和凍害條件下細胞內的保護物質[18],研究普遍認為,低溫脅迫下其含量的增加能夠提高植物抗逆性[19-21]。在本研究中,低溫脅迫下可溶性糖呈現先增高后下降的趨勢,前期其含量的增加是植株啟動自身防御機制的表現,后期的下降過程是蘋果花受到不可逆傷害的表現。本研究表明,脯氨酸、MDA、可溶性糖和可溶性蛋白質對低溫響應的臨界溫度范圍在-2.5~-3.5 ℃,這與李紅英等研究的蘋果花期低溫敏感范圍[22]具有較好的對應關系。
植物的抗寒性機理是一個復雜的過程,本研究只選取了1個品種進行測定,由于蘋果花期相對較短,本試驗沒有細分花苞期、開放期和落花期。4月22日測定結果和23日有所差別,可能是受花期發育進程的影響。另外,由于試驗需要一定的樣本量,因此沒有細分為花瓣、花藥和柱頭等器官,今后可通過增加測定品種和其他抗寒性指標進一步深入研究蘋果花的抗寒性機理。
參考文獻:
[1]李 國. 寧夏蘋果產業發展提升思考與建議[J]. 寧夏林業,2016(3):21-22.
[2]王來平,薛曉敏,路 超,等. 幾種抗逆增強劑對蘋果花與幼果抗寒力的效果[J]. 江蘇農業科學,2017,45(9):109-112.
[3]付 超,周雪玲,朱春林. 應用隸屬函數法綜合評價高酸蘋果抗寒性及果實品質[J]. 北方園藝,2017,41(2):11-15.
[4]向春燕,樊 麗,亢 鍵,等. 低溫脅迫下嘎拉蘋果果實葉綠素熒光參數與其他生理和品質指標的相關性[J]. 西北農業學報,2014,23(8):135-141.
[5]李江闊,劉 暢,張 鵬,等. 低溫條件下不同時期1-MCP處理對金冠蘋果生理和品質的影響[J]. 食品科學,2015,36(18):220-224.
[6]張志良,瞿偉菁. 植株生理學實驗指導[M]. 3版.北京:高等教育出版社,2003:274-276.
[7]王學奎,黃見良. 植物生理生化實驗原理與技術[M]. 北京:高等教育出版社,2015.
[8]馮建燦,張玉潔,楊天柱. 低溫脅迫對喜樹幼苗SOD活性、MDA和脯氨酸含量的影響[J]. 林業科學研究,2002,15(2):197-202.
[9]王 華,王 飛,陳登文,等. 低溫脅迫對杏花SOD活性和膜脂過氧化的影響[J]. 果樹學報,2000,17(3):197-201.
[10]閆忠業,呂天星,王冬梅,等. 低溫下六個蘋果新品種抗性生理指標的比較[J]. 植物生理學報,2015,51(1):93-96.
[11]鄧鳳飛,楊雙龍,龔 明. 細胞信號分子對非生物脅迫下植物脯氨酸代謝的調控[J]. 植物生理學報,2015,51(10):1573-1582.
[12]湯章城. 逆境條件下植物脯氨酸的累積及其可能的意義[J]. 植物生理學報,1984,20(1):15-21.
[13]Kishor P,Sreenivasulu N. Is proline accumulation per se correlated with stress tolerance or is proline homeostasis a more critical issue[J]. Plant,Cell & Environment,2014,37(2):300-311.
[14]龔 明,劉友良,朱培仁. 低溫下稻苗葉片中蛋白質及游離氨基酸的變化[J]. 植物生理學通訊,1989,25(4):18-22.
[15]徐炯達,樸成日,高文浩,等. 延邊地區蘋果梨抗寒生理研究[J]. 延邊大學農學學報,2009,31(1):26-31.
[16]王曉宇,張艷娥,張林生. 4種非生物脅迫下小麥幼苗表型及可溶性蛋白含量的變化[J]. 干旱地區農業研究,2018,36(2):113-117,169.
[17]令 凡,焦 健,李朝周,等. 不同油橄欖品種對低溫脅迫的生理響應及抗寒性綜合評價[J]. 西北植物學報,2015,35(3):508-515.
[18]王孝宣,李樹德,東惠茹,等. 番茄品種耐寒性與ABA和可溶性糖含量的關系[J]. 園藝學報,1998,25(1):57-61.
[19]李自龍,徐雪風,焦 健,等. 不同品種油橄欖離體葉片對滲透脅迫的生理響應及其抗旱機制[J]. 西北植物學報,2014,34(9):1808-1814.
[20]徐衛平,蔣景龍,任緒明,等. 低溫脅迫對3種柑橘幼苗細胞膜及滲透調節的影響[J]. 分子植物育種,2017,15(3):1104-1108.
[21]Couée I,Sulmon C,Gouesbet G,et al. Involvement of soluble sugars in reactive oxygen species balance and responses to oxidative stress in plants[J]. Journal of Experimental Botany,2006,57(3):449-459.
[22]李紅英,段曉鳳,王 靜,等. 寧夏蘋果花期霜凍敏感性指標研究[J]. 干旱地區農業研究,2015,33(6):184-188.張 宇,徐 剛,樊小雪,等. 生物炭施用及氮肥與有機肥配施對大蒜養分積累及品質的影響[J]. 江蘇農業科學,2020,48(11):115-121.
