銅尾礦白羊草各生長階段根際土壤叢枝菌根真菌群落特征
2020-08-19 08:26:30姚玉珊郭婷艷
生態學報 2020年13期
關鍵詞:生長
賈 彤,姚玉珊,郭婷艷
黃土高原生態恢復山西省重點實驗室,山西大學黃土高原研究所,太原 030006
我國金屬礦產資源豐富,隨著金屬礦產資源開發活動的迅猛發展,礦山周邊土壤環境受到污染的現象日趨嚴重,受損生態系統的修復越來越被重視。山西省作為全國能源重化工基地,工礦企業多、土地破壞面積大、土壤修復任務重,礦區受損生態系統恢復是山西省生態文明建設面臨的重大挑戰之一。山西省運城市垣曲縣,中條山集團北方銅業銅礦峪礦作為全國非煤系統最大的井下開采礦山,在浮選銅精礦過程中廢棄尾砂、銅礦石經破碎球磨等工序加工至顆粒度很小的細砂狀[1]。大量的尾礦堆積導致尾礦庫面積迅速增大。尾礦砂有機質含量低、養分貧瘠、重金屬污染嚴重,這使礦區生態系統生物多樣性、群落結構和功能均受到不同程度的影響[2- 5]。近年來,微生物修復技術因其低耗、高效凈化和安全備受人們關注,特別是菌根修復在眾多領域中得到了廣泛的應用。微生物技術可加快生土熟化,加速植被恢復,是培肥礦區土壤和修復礦區生態的一個重要途徑。
叢枝菌根真菌(Arbuscular mycorrhizal fungi,AMF)是陸地生態系統最重要的土壤微生物之一,自然界中80%以上有花植物都能被叢枝菌根真菌侵染。AMF的宿主專一性相對較低,全球 AMF 物種豐富度范圍在341個OTU(分類操作單元,operational taxonomic unit)至1600個OTU,甚至可能多余 1600 個 OTU[6]。這些 AMF 與大約 20 萬種植物形成共生關系[7]。AM 結構分為表面和根內結構兩部分,其中,根系表面菌絲包括從根內長出和生長于土壤中附著于根系表面的菌絲,根外菌絲為根內菌絲長出和土壤中生長分布在土壤中的菌絲。研究表明,蘆葦根系周圍寄主植物根系的理化特性決定了蘆葦根系AMF的種類多樣性和侵染強度[8],且土壤理化因素是影響宿主植物根系和根際AMF多樣性和群落結構的重要因素[9-10]。AMF多樣性對不同生態系統的植物群落結構、多樣性和生產力有重要影響,在植被重建、生態恢復及植物多樣性維持等當面扮演著重要角色[11-12]。已有研究表明,不同生態系統中AMF多樣性表現出明顯差異[13]。有研究發現,自然生態系統AMF物種數在熱帶森林中最多,而在人為干擾的環境中最少,如農田和污染區[14],并且污染生態系統中AM真菌多樣性指數最低[15]。AM真菌的數量及群落組成,間接受到土地利用方式、土壤耕作等人為干擾的影響,例如,北方農牧交錯帶在草地轉變為農田的過程中,由于土壤結構及養分狀況的改變,間接導致AM真菌多樣性顯著下降及AMF群落結構的改變[16]。AMF生態適應性在很大程度上決定了其生態生理功能的發揮。AMF可以促進宿主植物吸收水分和養分,對植物生長具有重要作用,例如,干旱、重金屬以及極端溫度等逆境脅迫下可提高植物抗逆性[11,17]。因此,研究受損生態系統中AMF多樣性及其關鍵影響因子,有助于生態恢復中可利用菌種資源的開發和利用。
本實驗選擇銅尾礦區主要恢復植被白羊草為研究對象,研究重金屬污染環境下白羊草各生長階段中AMF群落結構與多樣性的變化,探討白羊草各個生長階段中AMF與環境因子的關系,闡明AMF在銅尾礦重金屬脅迫環境中的多樣性以及對白羊草不同發育階段的生態因子響應機制,為銅尾礦生態恢復過程中發掘和利用AMF資源提供科學依據,選育具有抗逆性的植物-微生物共生體,進而提高銅尾礦區的生態修復效率。
1 材料與方法
1.1 研究區概況
試驗區位于山西省運城市垣曲縣,北方銅業銅礦峪礦十八河尾礦庫(35°15′—35°17′N,118°38′—111°39′E),該尾礦壩建于1969年,位于選礦廠西南6 km的河谷中,由上游攔洪壩和下游尾礦壩及兩側山梁圍成。選礦產生的廢棄物以礦砂的形式逐年堆積于十八河尾礦壩,每3—5年堆積形成一個新子壩,并在表層覆蓋30 cm客土,尾礦壩初期壩底標高486 m,壩頂標高509 m,現已堆筑16道子壩,堆積高度84 m,總坡比1∶6。該區域屬大陸性季風氣候,四季分明,春季干旱多風,夏季雨量集中,冬季少雪干燥。年均降雨量為780 mm,年均氣溫14℃,無霜期大于200 d[18]。
1.2 樣品采集
以銅尾礦壩的優勢種白羊草為研究對象,2017年6月初、7月末和9月中旬在S536號子壩(恢復20年[1])對當年新生白羊草進行根際土壤采集,選擇采用五點取樣法在1 m×1 m的樣方內對白羊草根際5—20 cm 深度土壤進行采集,去除植物少部分枯落物及老舊根系后,選擇當年新生根系,利用抖落法收集根際土壤樣本,各取樣點之間的距離至少大于50 m,采樣期間佩戴無菌手套,避免不同樣點間土壤樣品污染。采集的樣本分為兩份,一份用于土壤理化性質的測定,另一份帶回實驗室置于-80℃超低溫冰箱保存,用于微生物高通量測序。
1.3 研究方法
1.3.1土壤理化性質及酶活性的測定
用點位法測定土壤pH(土水比為1∶2.5)。土壤總碳和總氮含量使用元素分析儀(vario EL/MACRO cube, Elementar, Hanau, Germany)進行測定。采用苯酚鈉—次氯酸鈉比色法對土壤脲酶活性進行測定,過氧化氫酶的測定采用高錳酸鉀滴定法,蔗糖酶的測定采用3,5-二硝基水楊酸比色法,堿性磷酸酶的測定采用磷酸苯二鈉比色法[9]。
1.3.2白羊草根際土壤AMF群落的測定
用E.Z.N.A.? soil 試劑盒(Omega Bio-tek, Norcross, GA, U.S.)進行基因組提取,DNA濃度和純度利用NanoDrop2000進行檢測,利用1%瓊脂糖凝膠電泳檢測DNA提取質量。PCR所用的引物選用AML1/AML2為第一對引物[19],AMV4.5NF/AMDGR[20]為第二對引物。使用2%瓊脂糖凝膠回收PCR產物,利用Illumina公司的Miseq PE300平臺進行測序,白羊草根際土壤樣品送到上海美吉生物醫藥科技有限公司完成高通量測序。
1.4 數據處理
使用Trimmomatic軟件對原始序列進行數據去雜和質控過濾,得到優化數據。使用的UPARSE軟件(version 7.1 http://drive5.com/uparse/)對得到的高質量序列按照97%的相似度進行操作分類單元(Operational taxonomic units, OTU)的聚類。對OTU代表序列進行物種注釋,用Qiime軟件(Version1.7.0)中的blast方法[21]與Unit 數據庫[22]行物種注釋分析,并分別在各個分類水平進行統計AMF群落組成。采用Microsoft Excel進行數據處理和作圖,利用SPSS 19.0進行單因素方差分析及Canoco 4.5進行RDA分析。
2 結果與分析
2.1 白羊草不同生長階段根際土壤的理化性質
白羊草不同生長階段的根際土壤理化性質表明,土壤養分(總碳和總氮)及土壤碳氮比均表現為幼苗期顯著高于分蘗期和成熟期的根際土壤。土壤pH為堿性,且白羊草分蘗期的根際土壤pH達最大值(8.9),顯著高于幼苗期(8.2)和成熟期(8.1)的土壤pH(表1)。對于土壤酶活性而言,過氧化氫酶和堿性磷酸酶均在白羊草成熟期達到最大值,而脲酶和蔗糖酶在分蘗期顯著高于其他生長階段(表1)。這表明白羊草各個生長階段土壤理化和酶活性都存在顯著差異。
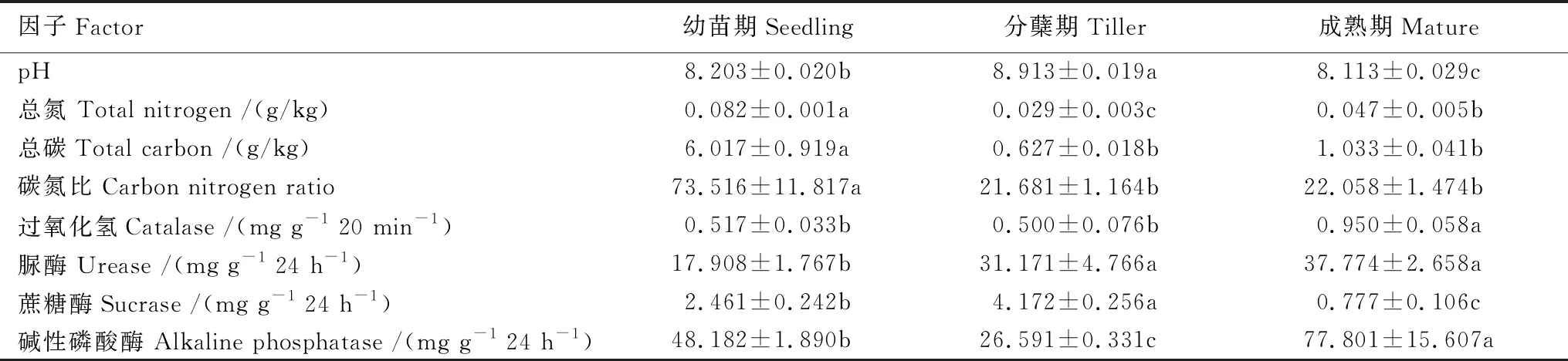
表1 白羊草不同生長階段根際土壤理化特征及酶活性
2.2 白羊草根際土壤AMF分類
不同生長階段白羊草根際土壤樣本共得到6目,白羊草各個生長階段根際土壤中優勢菌均為球囊霉目(Glomerales),其中,白羊草幼苗期根際土壤中球囊菌目(Glomerales)占97.5%,分蘗期為56.82%,成熟期球囊菌目(Glomerales)占75.14%。和球囊霉目類似,白羊草各個生長階段的優勢菌群也具有明顯的差異。分蘗期多樣孢囊霉屬(Diversispora)占優勢,其相對豐度達到31.64%。白羊草成熟期的類球囊霉屬(Paraglomus)相對豐度最高(23%)(表2)。在OTU水平上,白羊草各生長階段根際土壤中屬于AMF的OTU分別為28個(幼苗期)、19個(分蘗期)和15個(成熟期)。
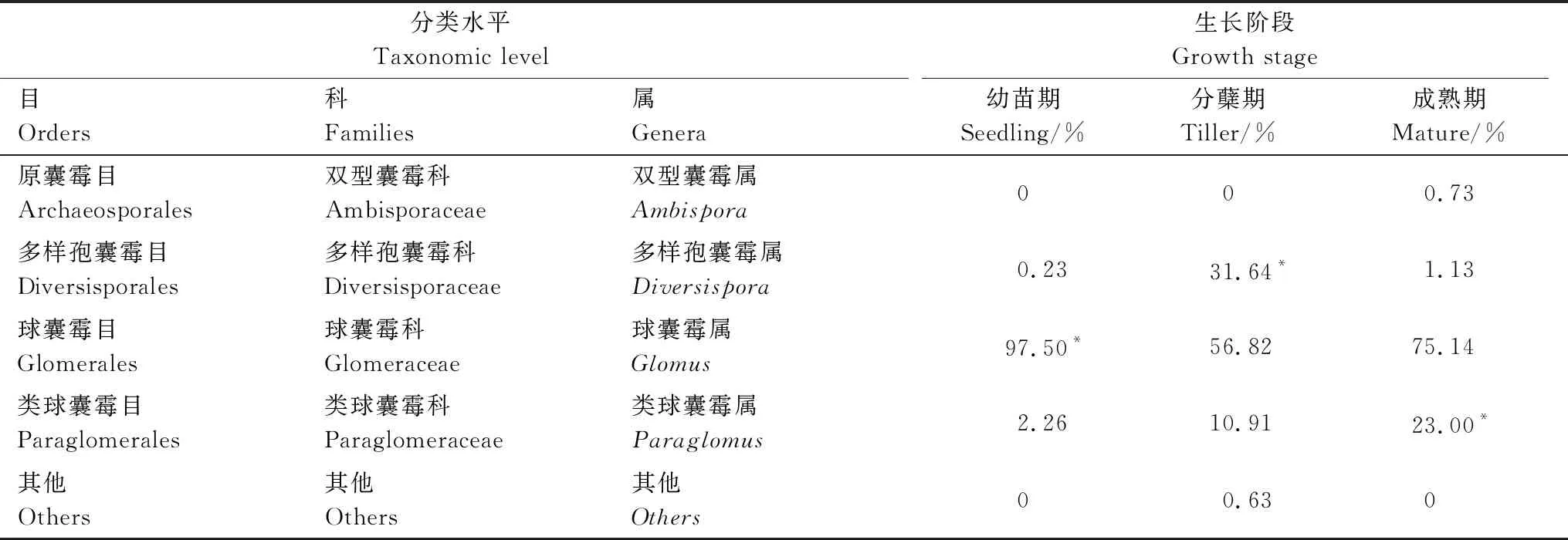
表2 白羊草不同生長階段根際土壤中AMF分類情況
2.3 白羊草不同生長階段根際土壤AMF群落結構與多樣性
由圖1可知,白羊草不同生長階段根際土壤的AMF群落結構具有顯著差異,白羊草3個生長階段之間共有的OTU數為8個,白羊草幼苗期特有的OTU數為9個,分蘗期和成熟期AMF群落中的特有OTU數分別為5個和2個。白羊草幼苗期的GlomusperpusillumVTX00287菌占比最大(29.8%),分蘗期和成熟期相對豐度最多的叢枝菌根真菌均是GlomusMOG18VTX00064,分別達26.8%和27.8%(圖1)
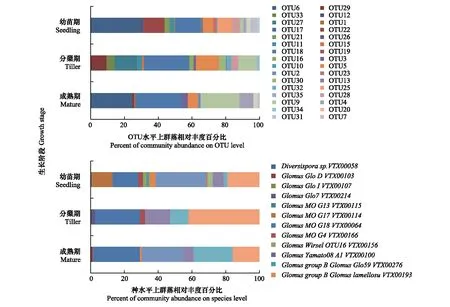
圖1 在OTU(分類操作單元,operational taxonomic unit)水平和種水平上白羊草不同生長階段AMF群落相對豐度Fig.1 Relative abundance of AMF community on OTU and species levels in different growth stages of B. ischaemumOTU:分類操作單元,operational taxonomic unit,不同數字表示各OTU編號;Diversispora:多孢囊霉屬;Glomus:球囊霉屬;Paraglomus:類球囊霉屬;不同屬名后字母及編號:在種水平上各屬叢枝菌根真菌的菌株編號
所有采樣點測序文庫的覆蓋度均達到99%以上,說明絕大部分AMF序列可以被測出,測序結果具有較好的代表性。隨著白羊草的生長,其根際土壤AMF的香農指數逐漸降低,即在幼苗期達到最大值(2.339),而辛普森指數在成熟期達到最大值(0.204)。ACE指數和Chao1指數變化趨勢與香農指數一致,均在幼苗期達到最大值,在成熟期豐富度指數最低(表3)。
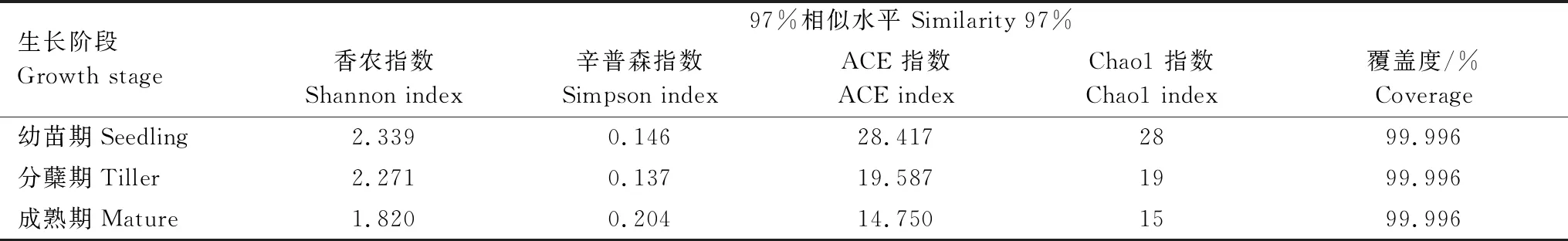
表3 不同生長階段白羊草根際土壤AMF的OTU豐度與多樣性指數
2.4 土壤理化性質及酶活性對白羊草AMF群落的影響
白羊草不同生長階段中,AMF群落受到土壤理化性質和土壤酶活性的影響(圖2),為研究這些生態因子對AMF群落結構的影響大小以及各時期影響AMF的關鍵因素,使用軟件Canoco 4.5對實驗數據進行了RDA約束排序分析,得到在科水平上AMF的群落組成與6種土壤理化性質及4種土壤酶活性之間的關系。結果表明土壤理化性質和土壤酶活性分別能夠解釋81.0%和83.0%的變異信息,白羊草各生長階段AMF群落組成具有差異性,其中,第一排序軸解釋了68.2%,第二排序軸解釋了12.8%的信息,主要影響幼苗期白羊草根際土壤球囊霉科(Glomeraceae)的是土壤養分(總碳和總氮)和碳氮比。分蘗期白羊草根際土壤中的多樣包囊霉科(Diversisporaceae)與土壤pH顯著相關。分蘗期白羊草根際土壤中的多樣孢囊霉科(Diversisporaceae)的叢枝菌根真菌主要受土壤pH的影響(圖2)。白羊草幼苗期,球囊霉科(Glomeraceae)與堿性磷酸酶顯著正相關;分蘗期多樣孢囊霉科(Diversisporaceae)與蔗糖酶顯著相關,成熟期的雙型囊霉科(Ambisporaceae)與土壤過氧化氫酶顯著相關,類球囊霉科(Paraglomeraceae)與土壤脲酶也具有顯著正相關關系(圖2)。這表明不同生長階段白羊草根際土壤的叢枝菌根真菌群落受土壤理化性質和酶活性的影響大小也各有差異。
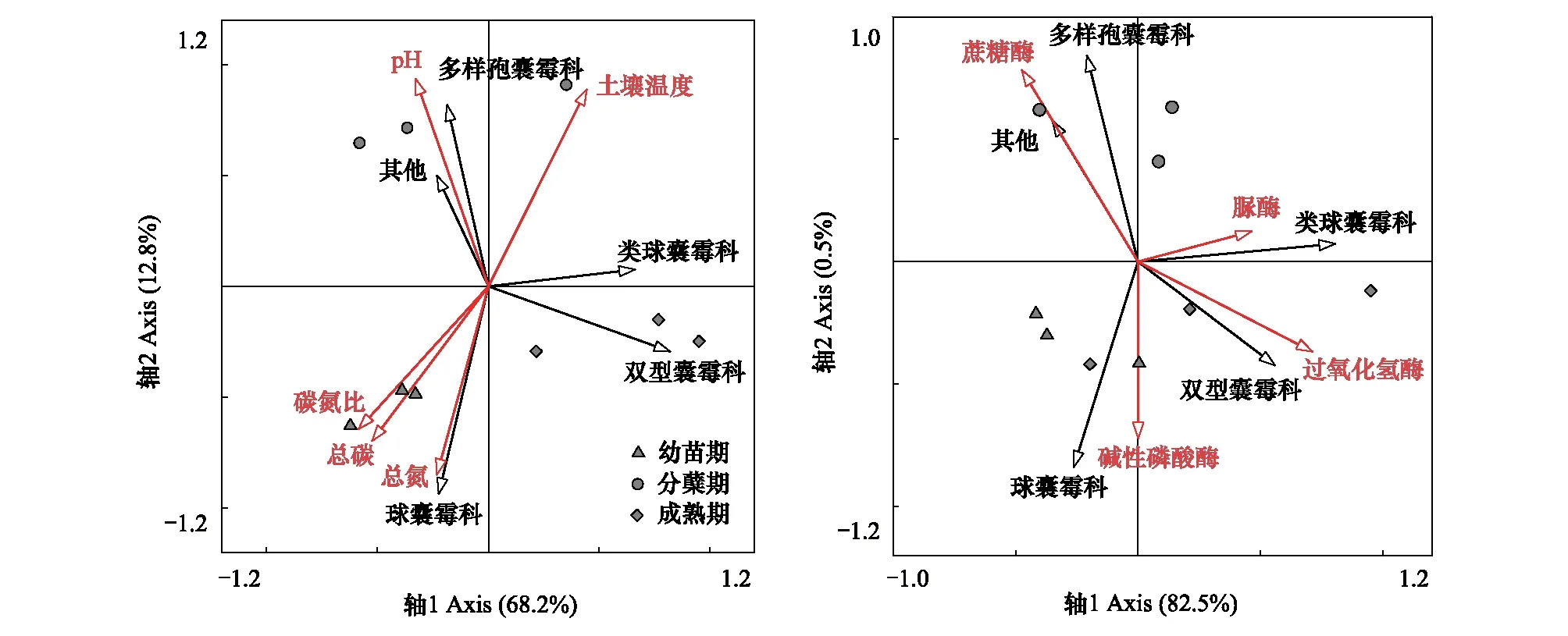
圖2 不同生長階段白羊草AMF群落與土壤理化性質(A)及土壤酶活性(B)的RDA分析Fig.2 Redundancy analysis (RDA) bi-plot of AMF community at family level, soil properties (A) and enzyme activity (B)
3 討論
AMF群落主要分布在根系周圍的土壤中,與土壤環境緊密相關。土壤養分可直接或間接影響AMF侵染、菌根共生體系發育、產孢和分布[23]。Wang[24]等對毛烏素沙漠AMF群落結構和多樣性的研究發現,不同土壤深度的植物AMF組成中,球囊霉屬(Glomus)和無梗囊霉屬(Acaulospora)占總數的70%以上,多樣孢囊霉屬(Diversispora)和巨孢囊霉屬(Gigaspora)僅在淺層土壤中發現。本研究中,處于相同生長階段,白羊草根際土壤的AMF群落組成也具有明顯差異,分蘗期多樣孢囊霉屬(Diversispora)占優勢,成熟期類球囊霉屬(Paraglomus)相對豐度最高,這可能由于分蘗期植物根系所處土層較淺,而隨著植物生長,根系不斷伸長,所處土層深度增加,優勢菌逐漸變為球囊霉屬(Glomus)。有研究發現, AMF的豐度和密度隨著土壤肥力的提升均呈增加趨勢[25],并且在一定范圍內AMF孢子數量隨有機質含量的升高而增多[26]。本研究中白羊草幼苗期,根際土壤中球囊霉科(Glomeraceae)的主要因子是土壤總碳、總氮和碳氮比,這與前人的研究結果類似。土壤pH直接影響叢枝菌根的形成、AMF的產孢和種屬分布。有研究表明,不同AM真菌種的最適pH不同,球囊霉屬(Glomus)能適應較廣泛的土壤pH范圍,但喜在堿性和中性土壤中生長[27-28]。相一致地,本研究發現白羊草各生長階段的優勢AMF均為球囊霉屬(Glomus),這可能與銅尾礦土壤pH呈堿性(8.1—8.9)有關。此外,白羊草分蘗期,根際土壤中的多樣孢囊霉科(Diversisporaceae)與土壤pH顯著相關,這表明相對于AMF物種而言,環境對各AMF的篩選差異更加直接和顯著[29]。
AMF群落的環境影響因子除非生物因子外,生物因子如宿主植物及人為干擾也會對AM真菌的分布產生明顯的影響。AMF與植物形成的共生體系,二者通常具有緊密聯系,地上植被可對AMF群落產生關鍵影響[30-32]。有研究表明,AMF的物種數與植物群落的物種數存在顯著正相關關系[31]。本研究中,不同生長發育階段的白羊草AMF的物種組成和多樣性表現出明顯差異,這可能由于不同生長階段宿主植物所需的光合產物不同,因此,輸送給對使自身生長更為有利的AMF具有一定差異,進而影響各個生長階段AMF的群落組成和多樣性[33],而且植被恢復過程中,白羊草周圍的非菌根植物或弱菌根植物的存在也會對AMF的群落組成及多樣性產生影響[34]。此外,AMF與土壤中其他微生物之間關系復雜,可相互促進、抑制或互不影響[35]。研究表明,逆境條件下AMF有助于提高土壤中微生物的量,同時也提高了土壤中相關酶的活性[36]。本研究中,成熟期白羊草根際土壤中類球囊霉科與土壤脲酶顯著正相關,這與前人的研究結果一致。
綜上所述,各生物與非生物因子并不是獨立地對AMF群落產生作用,而是作為一個整體綜合發揮作用[33]。同時,由于銅尾礦區重金屬污染嚴重[3- 4],因而,重金屬可能對AMF群落結構與多樣性也有重要的影響,而白羊草根際土壤中AMF的優勢物種在脅迫環境中表現出良好的適應性,為進一步研究重金屬脅迫下AMF群落生態功能提供科學基礎,同時,白羊草各個生長階段的優勢AMF可為礦區生態修復提供可利用的菌種資源,從而加速銅尾礦區受損生態系統的恢復。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
