杉木人工林凋落物添加與去除對土壤碳氮及酶活性的影響*
2020-08-25 03:37:16阮超越劉小飛呂茂奎李曉杰付作琴任寅榜謝錦升
土壤學報 2020年4期
關鍵詞:影響
阮超越,劉小飛,呂茂奎,李曉杰,付作琴,任寅榜,黃 俊,謝錦升?
杉木人工林凋落物添加與去除對土壤碳氮及酶活性的影響*
阮超越1,劉小飛1,2,3,呂茂奎1,李曉杰1,付作琴1,任寅榜1,黃 俊1,謝錦升1,2?
(1. 福建師范大學地理科學學院,福州 350007;2. 濕潤亞熱帶山地生態國家重點實驗室培育基地,福州 350007;3. 福建三明森林生態系統與全球變化研究站,福建三明 365000)
為了解未來氣候變化過程中森林生產力增加的背景下,凋落物增加如何影響土壤碳氮過程,在杉木人工林中通過模擬實驗研究凋落物添加(一倍)與去除對土壤中碳氮、碳氮同位素(δ13C、δ15N)、微生物生物量碳氮(MBC、MBN)及酶活性的影響。結果表明:凋落物添加后土壤中氮獲得酶(β-N-乙酰氨基葡萄糖苷酶)活性顯著上升,加速對土壤中有機質的分解獲取氮素;凋落物添加與去除處理對土壤碳的影響較小,土壤有機碳(SOC)與可溶性有機碳(DOC)均未發生顯著變化;土壤中δ13C豐度與凋落物處理之間未呈現出相關規律性,而δ15N豐度在凋落物添加處理后顯著上升。這些結果說明,凋落物處理對杉木林土壤中氮的影響較為敏感,對土壤碳的影響較小。因此,未來氣候變化導致森林生產力提高、凋落物輸入增加,可能會導致土壤中氮素的損失,迫使土壤微生物分泌更多的氮獲得酶同植物競爭土壤氮,最終可能會造成土壤碳氮循環的不平衡,對整個生態系統造成嚴重影響。
凋落物處理;土壤碳氮同位素;酶活性
森林生態系統是陸地生態系統的主體部分,其生物量以及生產量占到陸地生態系統的90%和70%[1],儲存著陸地生態系統45%左右的碳,在維持全球碳氮平衡、減緩大氣的溫室效應等方面發揮著重要的作用。凋落物作為森林生態系統的重要組成部分,是土壤有機質的重要物質源也是土壤碳氮的主要來源,凋落物每年通過分解歸還到土壤里的有機碳約為50 Gt、歸還到土壤中的總氮大約是植物所需氮量的70%~80%[2]。凋落物的輸入深刻影響著森林生態系統的碳氮循環過程。
全球氣候變暖已成為不爭的事實。隨著氣溫的升高和大氣CO2濃度增加可能會導致我國熱帶亞熱帶地區森林生產力增加,新陳代謝增強,凋落物增多[3-5]。凋落物的增加會改變對土壤中碳氮的輸入,影響土壤中的碳氮過程。一方面有觀點認為,凋落物輸入量的增加促進了土壤中有機碳的積累[6-7],另一方面有觀點認為凋落物量的增加導致向土壤中輸入更多的新鮮可溶性有機質,加速土壤中有機碳氮的分解,促使土壤有機碳氮產生激發效應[8-9],并最終導致土壤有機碳氮含量的下降。凋落物對土壤有機質的影響取決于凋落物自身的特性和土壤條件。傳統的觀點認為土壤有機質長期積累主要由難分解(高C/N)植物凋落物形成的,因為高質量(低C/N)的凋落物降解速度更快,不易在土壤中積累,而難分解的凋落物通過抵抗降解在土壤中積累[10]。然而越來越多的研究提出了相反的觀點,有研究通過室內培養實驗發現,低質量的凋落物輸入增多可能會導致土壤更加缺氮,加劇了微生物對N素的需求,促進微生物對土壤有機質的分解從而獲取N素,可能不利于土壤有機質的積累[11];高質量(低C/N)凋落物有利于土壤微生物的轉化和吸收,并最終以微生物殘體的形式進入土壤,有利于土壤有機質的積累[12]。
我國亞熱帶地區屬于海洋性季風氣候,分布著世界上面積最大的常綠闊葉林。然而,過去幾十年來由于對木材和其他林產品的需求,大面積天然常綠闊葉林被人工林、特別杉木人工林取代[13]。現有杉木林面積約900萬hm2,占全國人工林面積的30%[14]。那么未來氣候變化背景下,杉木人工林土壤碳氮固持能力對亞熱帶地區具有重要的影響。假定未來氣候變化(如氣溫升高,CO2濃度增加)能夠促進亞熱帶杉木人工林生產力的提高,從而增加地上凋落物輸入,那么杉木凋落物輸入量增加,對杉木林中土壤碳氮過程產生怎樣的影響,土壤中碳氮固持是增多還是減少?針對這個科學問題,本研究以亞熱帶杉木人工林為研究對象,通過對林中土壤設置去除與添加(一倍)凋落物處理為期四年的控制試驗,研究杉木林凋落物輸入量的變化對土壤碳氮循環過程產生的影響,為進一步認識全球變化背景下亞熱帶杉木人工林土壤碳氮過程的響應機制提供依據。
1 材料與方法
1.1 研究區概況
研究樣地位于福建三明全球變化森林生態研究站(26o09′24″N,117o28′03″E),地處武夷山東南、戴云山西北,以低山丘陵為主,平均海拔300 m,屬于中亞熱帶季風氣候。實驗3年期間(2013—2015年)年均降水量1 749 mm,主要集中在3—8月份。年均氣溫18.7℃,蒸發量1 585 mm,相對濕度81%,無霜期達300 d。植被以常綠闊葉林為主,森林覆蓋率可達67.8%[15],土壤以花崗巖發育的紅壤為主。1976年將米櫧天然林皆伐、煉山后營造成杉木人工林。樣地林分密度2 858株·hm–2,平均樹高18.2 m、胸徑15.6 cm,林下植物以毛冬青()、芒萁()、狗骨柴()為主。
1.2 試驗設計與樣品采集
2012年6月,在杉木人工林設置3個20 m×20 m的標準樣地,在每個樣地內設3個1 m×1 m的小區作不同處理:(1)CK,保留凋落物,即不做任何處理;(2)LE,去除凋落物,去除小區地表全部凋落物,并在小區上方0.5 m高處放置1.5 m×1.5 m大小的正方形尼龍網,阻止凋落物掉入;(3)LA,添加凋落物,每月將LE處理中尼龍網上截留的凋落物移至添加凋落物處理的小區內。每種處理設3個重復,共9個小區。
2016年5月,在每個小區中隨機多點采樣,去除凋落物,取0~10 cm的土層樣品并混合。隨后將土壤樣品放置于保溫箱中保存,然后帶回室內處理分析。在去除土壤中的根系以及石塊后,取部分新鮮土樣于–20℃保存待測土壤酶活性,其余土壤過2 mm篩,取少量鮮土自然風干后過0.154 mm鋼篩,測定土壤理化性質。
1.3 測定方法
土壤中全碳、全氮含量采用土壤碳氮元素分析儀(Elementar Vario MAX,Germany)進行測定,可溶性有機碳、氮(DOC、DON)含量采用硫酸鉀提取[16]、TOC-VCPN分析儀測定,微生物生物量碳、氮(MBC、MBN)采用氯仿熏蒸—硫酸鉀提取[17]、TOC-VCPN、流動(QC8000)分析儀測定,土壤中δ13C、δ15N采用穩定同位素質譜儀(IsoPrime100,Isoprime,Manchester,UK)測定。
土壤酶活性測定:參照 Saiya-Cork等[18]的方法測定土壤中6種參與碳、氮、磷循環相關的酶活性。用傘形酮(MUB)作為標示底物來測定4類水解酶活性:包括β-葡萄糖苷酶(βG)(EC3.2.1.21)、纖維素水解酶(CBH)(EC3.2.1.91)、β-N-乙酰氨基葡萄糖苷酶(NAG)(EC3.1.1.14)和酸性磷酸酶(AP)(EC 3.1.3.2)。氧化酶中酚氧化酶(PHO)(EC1.14.18.1)和過氧化物酶(PEO)(EC 1.11.1.7)測定采用L-二羥苯丙氨酸(DOPA)為底物,用多功能酶標儀(Synergy H4,美國)測定熒光度(水解酶)或吸光度(氧化酶),各種酶的縮寫、反應類型及所用底物見表1。
懸浮樣品制備:取1g鮮土置于125 mL、50 mmol·L–1、pH 5.0的醋酸鹽緩沖液中,用磁力攪拌器攪拌使其均質化,后用移液器取200 μL移入96孔微孔板,用酶標儀進行測定。
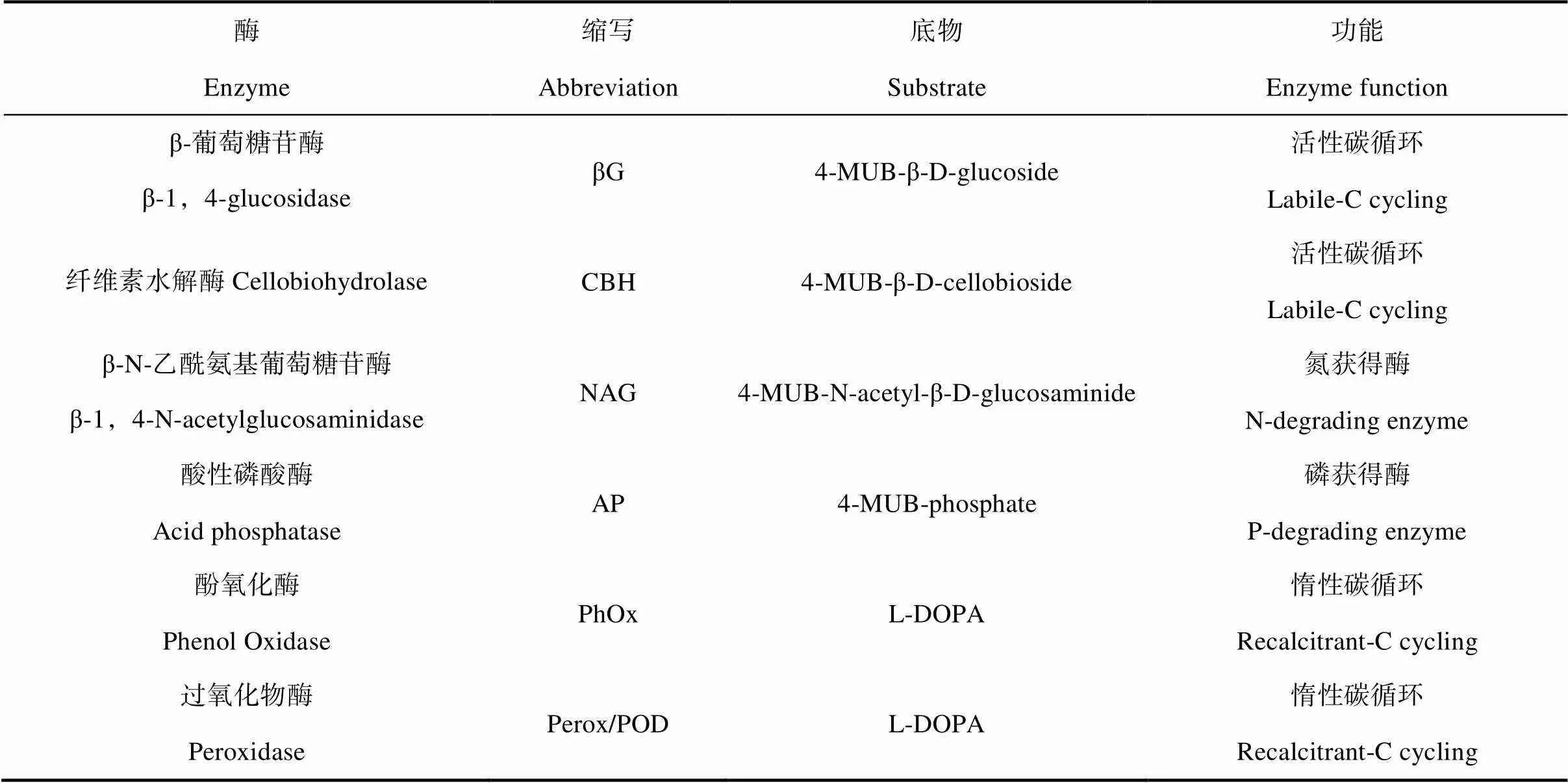
表1 研究選取的胞外酶相應的底物及功能
1.4 數據分析
所有數據運用SPSS20.0軟件進行分析,采用單因素方差分析(One-way ANOVA)進行差異性分析,運用Origin9軟件進行作圖。
2 結 果
2.1 凋落物添加與去除對土壤碳氮的影響
2.2 凋落物添加與去除對土壤碳氮同位素的影響
通過對土壤C、N同位素自然豐度的分析發現,添加凋落物較去除凋落物相比,δ13C豐度則顯著上升(<0.05,圖1),與對照相比,添加與去除凋落物后土壤中δ13C豐度均無顯著變化(>0.05),但δ13C豐度呈現出隨著凋落物下降而減少的趨勢。凋落物添加顯著提高土壤δ15N值,而凋落物去除降低了土壤δ15N值,說明隨著凋落物輸入增加土壤δ15N值也逐漸升高。
2.3 凋落物添加與去除對土壤酶活性的影響
凋落物添加后,土壤β葡萄糖苷酶、酸性磷酸酶活性顯著下降(<0.05),而β-N-乙酰氨基葡萄糖苷酶、酚氧化酶、過氧化物酶活性顯著升高(<0.05),纖維素水解酶呈現下降現象但未達到顯著水平(>0.05)。去除凋落物處理后,β葡萄糖苷酶、纖維素水解酶、酸性磷酸酶、過氧化物酶均顯著下降(<0.05),而β-N-乙酰氨基葡萄糖苷酶與酚氧化酶變化不顯著(>0.05)。
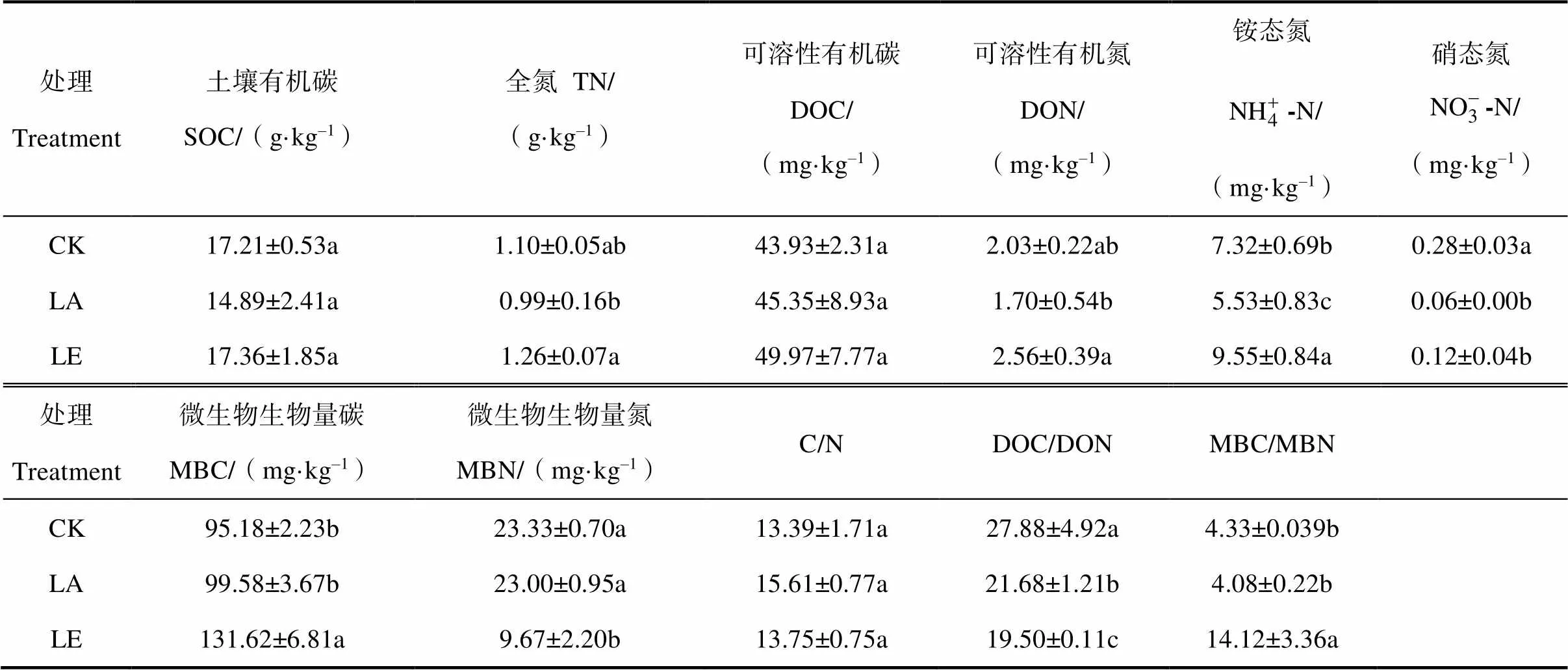
表2 凋落物處理下土壤碳氮含量變化
注:CK,對照;LA,凋落物添加;LE,凋落物去除。下同。同一列中無相同字母表示處理之間差異顯著(<0.05)。Note:CK,Control;LA,Litter added;LE,Litter excluded. The same below. Different letters in the same column indicate significant difference at 0.05 level.
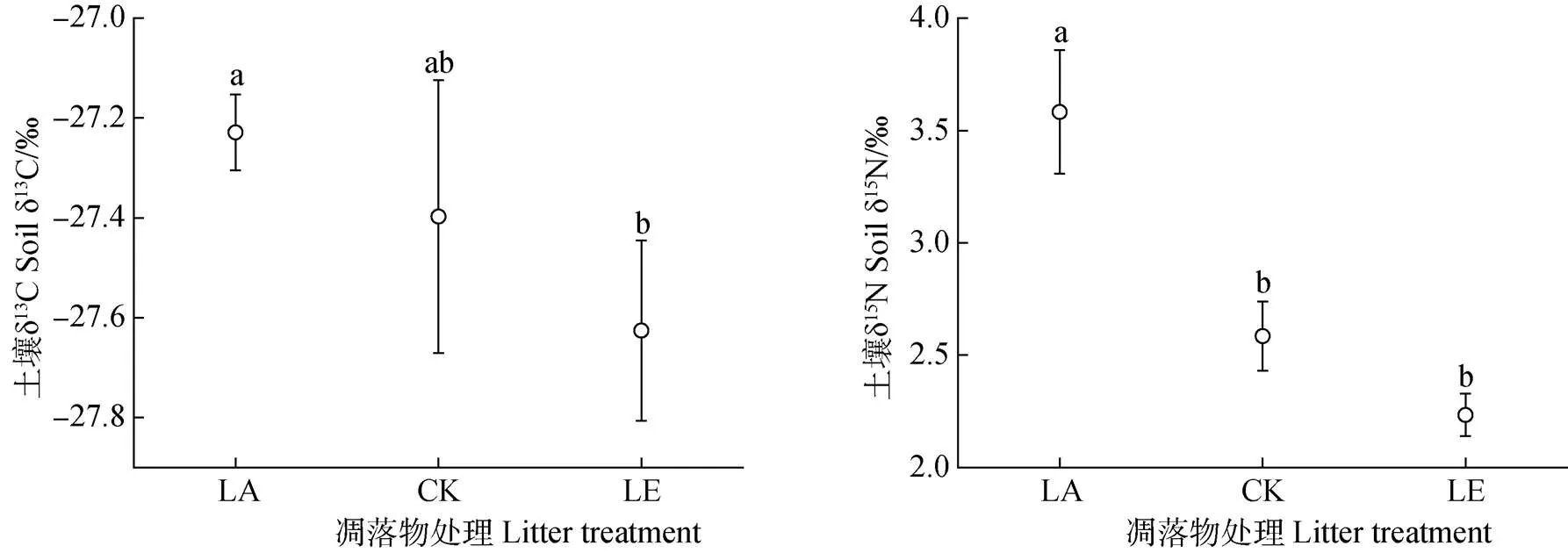
圖1 凋落物處理下土壤碳氮同位素的變化
3 討 論
3.1 土壤碳氮對凋落物處理的響應
凋落物可以通過兩種途徑來影響著土壤養分,一方面是通過淋溶、分解作用直接為土壤提供可溶性養分,N的含量是凋落物分解過程中主要的影響因素;另一方面通過影響土壤微生物來改變土壤的養分[19-20]。本研究中,杉木林凋落物處理四年后,添加凋落物處理土壤DON、銨態氮、硝態氮均發生顯著下降現象,而且凋落物添加處理后,DOC/DON明顯升高,表明杉木凋落物增多時土壤氮含量下降,而土壤微生物生物量碳氮未發生顯著變化,表明微生物量未發生變化,這與Wisconsin的橡樹林中添加凋落物導致土壤氮增加的結論相反[21],可能是因為本實驗中未對杉木根系做出處理。凋落物添加后可能對土壤中杉木細根產生了影響,凋落物作為土壤養分源,添加后會對細根的覓養行為產生影響,部分根系延伸至凋落物層及凋落物層與表層土壤連接處,導致細根生物量[22]和菌根侵染率[23]增加,更好地吸收凋落物層中淋溶出的及土壤中的養分,因此,添加凋落物可能會影響植物與微生物之間的養分利用關系。杉木作為內生菌根樹種,其菌根無法分解有機質,只能通過向腐生微生物提供光合產物,刺激腐生微生物分解土壤有機質,為其提供氮素,從而導致土壤中氮素消耗增加、土壤氮含量下降。數據顯示凋落物添加后土壤中氮獲得酶顯著升高,這進一步印證了由于土壤中氮素的損失導致微生物為滿足自身需求不得不增強β-N-乙酰氨基葡萄糖苷酶的分泌來獲取氮素(圖2)。此外,凋落物添加后導致土壤含水率增加[24],可能增強了土壤中氮的硝化-反硝化作用,提高了土壤氮氣態損失的風險。而無論是凋落物添加還是去除處理,對土壤有機碳的影響均不明顯,可能是因為凋落物分解對于土壤中總有機碳的改變需要較長的時間[25];孫麗娟等[26]在2010年鼎湖山地區針闊葉混交林中通過添加凋落物后發現,添加凋落物對土壤碳的影響不明顯。相比土壤氮而言,土壤有機碳本身的背景值較大,短期的控制實驗很難對土壤有機碳產生顯著的影響。

圖2 凋落物處理下土壤酶活性的變化
3.2 土壤C、N同位素對凋落物處理的響應
目前多數研究利用土壤C、N同位素自然豐度值的變化從微觀的角度來揭示土壤碳氮的變化[27-28]。因為同位素具有分餾效應,所以不同物質之間C、N同位素豐度有差異[29]。土壤中δ13C、δ15N組成能較好的闡明土壤中有機碳與全氮的遷移、轉化并能夠對土壤中有機質的分解程度進行評估,是研究土壤碳氮循環的重要工具[30]。有關研究發現,影響土壤有機碳中δ13C值的主要因素是土壤微生物的分解作用和植物凋落物及其根系的δ13C值[31]。本研究中土壤δ13C豐度與凋落物量并沒有呈現出相關規律性:凋落物添加與去除處理后δ13C相對于對照來說均未產生顯著變化。因此,從土壤δ13C的角度進一步說明了凋落物量的變化對土壤有機碳的影響較小。而且,凋落物添加處理后,土壤β葡萄糖苷酶顯著降低,一定程度上反應了土壤中活性碳組分的分解減少,因為凋落物添加處理后微生物具有豐富碳源,微生物不需要分泌多余的胞外酶來獲取碳源。土壤δ15N豐度在凋落物添加后顯著升高,凋落物去除后顯著降低,說明土壤中氮對凋落物的處理更為敏感,而土壤碳受其影響較小。有研究顯示[32],表層土壤氮同位素與植物體氮同位素具有顯著相關性,地上植被對土壤氮同位素具有重要影響,本研究中,凋落物添加后顯著提高了土壤δ15N豐度值,一般而言,土壤δ15N豐度值越高,說明這個系統越開放,土壤中N較容易損失[33],所以,凋落物添加后δ15N豐度的升高一定程度上反映了土壤中可能發生了氮素的轉移或損失,如土壤中DON、銨態氮和硝態氮顯著降低。杉木凋落物輸入增加導致β-N-乙酰氨基葡萄糖苷酶上升,主要與土壤中氮素的減少有關,凋落物添加后可能導致的植物細根生物量的增加會使得植物進一步吸收土壤氮素,微生物需要同植物共同競爭土壤氮素,從而使微生物分泌更多的氮獲得酶來滿足自身需求。微生物具有自身相對穩定的碳氮比,當土壤中氮素減少時,微生物就會通過調節自身對不同胞外酶的分泌,即分泌更多的氮獲得酶來獲取自身穩定的碳氮比。此外,凋落物添加處理后,土壤微生物C/N無顯著變化;但去除凋落物處理顯著降低了微生物C/N,這可能是因為凋落物去除一方面減少了有機碳的輸入,同時也會去除凋落物層中的微生物,導致了微生物群落結構發生了變化,造成微生物C/N下降[34]。
3.3 土壤酶活性對凋落物處理的響應
從土壤中酚氧化酶和過氧化氫酶的變化也可以證實上述推理,凋落物添加處理,酚氧化酶和過氧化氫酶活性均增加,而凋落物去除后兩種酶活性降低。相比C/N比較高的杉木凋落物,微生物會偏向選擇利用土壤中的有機質,而凋落物添加處理后,微生物碳源充足,氮素不足,微生物會利用新鮮凋落物作為碳源來獲取能量,去分解土壤有機質獲取氮素;由于杉木凋落物C/N比較高,木質素/N較高,那么微生物利用杉木凋落物時則需要分泌更多的酚氧化酶或者過氧化氫酶。因此,凋落物添加后酚氧化酶和過氧化氫酶活性增加是因為微生物需要分解杉木凋落物獲取碳源而滿足自身能量需求,并非反應微生物分解土壤難分解有機質。同理,凋落物去除后酚氧化酶和過氧化氫酶活性降低,則說明微生物對難分解有機物的分解減少,即相比對照而言,凋落物去除等于去除了供給微生物較高C/N比的底物,微生物失去了分解對象,沒有新鮮碳源作為能量來源,那么微生物的分解作用也降低,導致酚氧化酶和過氧化氫酶活性降低。而且,在凋落物去除處理后,土壤δ15N值顯著降低,說明土壤氮循環變得更加封閉和緩慢[25,27];同時土壤β葡萄糖苷酶、纖維素水解酶均出現不同程度下降現象;由于β葡萄糖苷酶、纖維素水解酶都參與土壤中碳的分解與循環[34],所以凋落物去除可能會減緩土壤碳氮循環速率。
4 結 論
土壤氮對杉木人工林中添加、減少凋落物的影響較為敏感,土壤碳受其影響不顯著。凋落物添加增強了植物對土壤中氮素的吸收,導致土壤中氮素的減少,而土壤微生物為了維持自身碳氮比,會分泌更多的氮獲得酶去分解土壤中的有機質來獲取氮素,使得微生物與植物之間競爭土壤氮,導致土壤中氮素的循環加快,最終可能導致土壤碳氮失衡。
[1] Post W M,Emanuel W R,Zinke P J,et al. Soil carbon pools and world life zones[J]. Nature,1982,298(5870):156—159.
[2] Gholz H L,Fisher R F,Prichett W L. Nutrient dynamics in slash pine plantation ecosystems[J]. Ecology,1985,66(3):647—659.
[3] Sombroek W,Nachtergaele F O,Hebel A. Amounts,dynamics and sequestering of carbon in tropical and subtropical soils[J]. Ambio,1993,22(7):417—426.
[4] Finzi A C,Allen A S,DeLucia E H,et al. Forest litter production,chemistry,and decomposition following two years of free-air CO2enrichment[J]. Ecology,2001,82(2):470—484.
[5] Ellsworth D S,Thomas R,Crous K Y,et al. Elevated CO2affects photosynthetic responses in canopy pine and subcanopy deciduous trees over 10 years:A synthesis from Duke FACE[J]. Global Change Biology,2012,18(1):223—242.
[6] Fang X,Zhao L,Zhou G Y,et al. Increased litter input increases litter decomposition and soil respiration but has minor effects on soil organic carbon in subtropical forests[J]. Plant and Soil,2015,392(1/2):139—153.
[7] Leff J W,Wieder W R,Taylor P G,et al. Experimental litterfall manipulation drives large and rapid changes in soil carbon cycling in a wet tropical forest[J]. Global Change Biology,2012,18(9):2969—2979.
[8] Fontaine S,Barot S,Barré P,et al. Stability of organic carbon in deep soil layers controlled by fresh carbon supply[J]. Nature,2007,450(7167):277—280.
[9] Kuzyakov Y. Priming effects:Interactions between living and dead organic matter[J]. Soil Biology & Biochemistry,2010,42(9):1363—1371.
[10] Zhang D Q,Hui D F,Luo Y Q,et al. Rates of litter decomposition in terrestrial ecosystems:Global patterns and controlling factors[J]. Journal of Plant Ecology,2008,1(2):85—93.
[11] Wang Q K,Wang S L,He T X,et al. Response of organic carbon mineralization and microbial community to leaf litter and nutrient additions in subtropical forest soils[J]. Soil Biology & Biochemistry,2014,71:13—20.
[12] Cotrufo M F,Wallenstein M D,Boot C M,et al. The Microbial Efficiency-Matrix Stabilization(MEMS)framework integrates plant litter decomposition with soil organic matter stabilization:Do labile plant inputs form stable soil organic matter?[J]. Global Change Biology,2013,19(4):988—995.
[13] Yang Y S,Xie J S,Sheng H,et al. The impact of land use/cover change on storage and quality of soil organic carbon in midsubtropical mountainous area of Southern China[J]. Journal of Geographical Sciences,2009,19(1):49—57.
[14] Li M X.A study on the restoration of the hydrological functions in the second generation Chinese fir plantation[D]. Changsha:Central South University of Forestry & Technology,2007. [李明學. 第二代杉木人工林水文功能恢復研究[D]. 長沙:中南林業科技大學,2007.]
[15] Liu X F,Lin T C,Yang Z J,et al. Increased litter in subtropical forests boosts soil respiration in natural forests but not plantations of[J]. Plant and Soil,2017,418(1/2):141—151.
[16] Jones D,Willett V. Experimental evaluation of methods to quantify Dissolved organic nitrogen(DON)and Dissolved organic carbon(DOC)in soil[J]. Soil Biology& Biochemistry,2006,38(5):991—999.
[17] Vance E D,Brookes P C,Jenkinson D S. An extraction method for measuring soil microbial biomass C[J]. Soil Biology & Biochemistry,1987,19(6):703—707.
[18] Saiya-Cork K R,Sinsabaugh R L,Zak D R. The effects of long term nitrogen deposition on extracellular enzyme activity in anforest soil[J]. Soil Biology & Biochemistry,2002,34(9):1309—1315.
[19] Liao L P,Lindley D K,Yang Y H. Decomposition of mixed foliar litter I. A microcosm study[J]. Chinese Journal of Applied Ecology,1997,8(5):459—464. [廖利平,D. K. Lindley,楊永輝. 森林葉凋落物混合分解的研究I.縮微(Microcosm)實驗[J]. 應用生態學報,1997,8(5):459—464.]
[20] Tu Y,You Y M,Sun J X. Effects of forest floor litter and nitrogen addition on soil microbial biomass C and N and microbial activity in a mixedandliaotungensis forest stand in Shanxi Province of China[J]. Chinese Journal of Applied Ecology,2012,23(9):2325—2331. [涂玉,尤業明,孫建新. 油松-遼東櫟混交林地表凋落物與氮添加對土壤微生物生物量碳、氮及其活性的影響[J]. 應用生態學報,2012,23(9):2325—2331.]
[21] Nielsen G E,Hole F D. A study of the natural processes of incorporation of organic matter into soil in the University of Wisconsin Arboretum[J]. Arts and Letters,1963,52:213—227.
[22] Wang W,Wu X G,Hu K,et al. Increased litterfall regulates fine root biomass and distribution in three typical forests in subtropical China[J]. Acta Ecologica Sinica,2016,36(11):3391—3401. [王微,伍小剛,胡凱,等. 凋落物輸入對中亞熱帶不同森林細根生物量及分布的影響[J]. 生態學報,2016,36(11):3391—3401.]
[23] Brzostek E R,Dragoni D,Brown Z A,et al. Mycorrhizal type determines the magnitude and direction of root-induced changes in decomposition in a temperate forest[J]. New Phytologist,2015,206(4):1274—1282.
[24] Li X J,Liu X F,Xiong D C,et al. Impact of litterfall addition and exclusion on soil respiration inplantation and secondaryforest in mid-subtropical China[J]. Chinese Journal of Plant Ecology,2016,40(5):447—457. [李曉杰,劉小飛,熊德成,等. 中亞熱帶杉木人工林和米櫧次生林凋落物添加與去除對土壤呼吸的影響[J]. 植物生態學報,2016,40(5):447—457.]
[25] Berg B. Litter decomposition and organic matter turnover in northern forest soils[J]. Forest Ecology & Management,2000,133(1/2):13—22.
[26] Sun L J,Zeng H,Guo D L. Effects of litter and mineral nitrogen input on soil organic carbon decomposition in subtropical mixed forest in Dinghu Mountain,South China[J]. Chinese Journal of Applied Ecology,2011,22(12):3087—3093. [孫麗娟,曾輝,郭大立. 鼎湖山亞熱帶常綠針闊葉混交林凋落物及礦質氮輸入對土壤有機碳分解的影響[J]. 應用生態學報,2011,22(12):3087—3093.]
[27] Duan Z H,Qiao Y M,Quan X L,et al. Analysis of nitrogen and carbon composition and stable isotope characteristics and physicochemical properties of wetland and grassland soil in source region of the Yellow River[J]. Journal of Soil and Water Conservation,2015,29(4):247—252,315. [段中華,喬有明,全小龍,等. 黃河源區濕地、草地土壤理化性質和碳氮組成及其穩定同位素特征分析[J]. 水土保持學報,2015,29(4):247—252,315.]
[28] Wang M L,Lai J P,Hu K T,et al. Compositions of stable organic carbon and nitrogen isotopes in wetland soil of Poyang Lake and its environmental implications[J]. China Environmental Science,2016,36(2):500—505. [王毛蘭,賴建平,胡珂圖,等. 鄱陽湖濕地土壤有機碳氮同位素特征及其環境意義[J]. 中國環境科學,2016,36(2):500—505.]
[29] Natelhoffer K J,Fry B. Controls on natural nitrogen-15 and carbon-13 abundances in forest soil organic matter[J]. Soil Science Society of America Journal,1988,52(6):1633—1640.
[30] Zeng W B. Changes of soil carbon and nitrogen content and isotope values under different land use conditions in Danjangkou Reservior[D]. Beijing:Graduate University of the Chinese Academy of Sciences,2012. [曾偉斌. 丹江口庫區不同土地利用下土壤碳氮含量及其同位素值的變化[D]. 北京:中國科學院研究生院,2012.]
[31] Billings S A,Richter D D. Changes in stable isotopic signatures of soil nitrogen and carbon during 40 years of forest development[J]. Oecologia,2006,148(2):325—333.
[32] Wang Z J,Liang X,He Q F,et al. Differential characteristics of soil δ15N under varying vegetation in Karst areas[J]. Acta Ecologica Sinica,2011,31(17):4970—4976. [汪智軍,梁軒,賀秋芳,等. 巖溶區不同植被類型下的土壤氮同位素分異特征[J]. 生態學報,2011,31(17):4970—4976.]
[33] Hobbie E A,Ouimette A P. Controls of nitrogen isotope patterns in soil profiles[J]. Biogeochemistry,2009,95(2/3):355—371.
[34] Bian X L,Zhao W L,Yue Z H,et al. Research process of soil enzymes effect on carbon and nitrogen cycle in agricultural ecosystem[J]. Chinese Agricultural Science Bulletin,2016,32(4):171—178. [邊雪廉,趙文磊,岳中輝,等. 土壤酶在農業生態系統碳、氮循環中的作用研究進展[J]. 中國農學通報,2016,32(4):171—178.]
Effects of Litter Carbon, Nitrogen and Enzyme Activity in Soil under Chinese Fir
RUAN Chaoyue1, LIU Xiaofei1, 2, 3, Lü Maokui1, LI Xiaojie1, FU Zuoqin1, REN Yinbang1, HUANG Jun1, XIE Jinsheng1, 2?
(1. School of Geographical Sciences, Fujian Normal University, Fuzhou 350007, China; 2. Cultivation Base of State Key Laboratory of Humid Subtropical Mountain Ecology, Fuzhou 350007, China; 3. Sanming Research Station of Forest Ecosystem and Global Change, Sanming, Fujian 365000, China)
【】Climate change triggers variation of soil net primary productivity, which are likely to affect litter input to forest soil. Litter is an important source of nutrients for plants and soil microorganisms in forest ecosystems and may also alter stocks and cycling of soil carbon (C) and nitrogen (N). However, so far little has been found in the literature about responses of soil C and N to litter input in subtropical forests. This study aims to understand how soil C and N respond to increased litter input in a Chinses fir plantation in South China, caused by climate change. 【】In this study, a four-year field experiment, designed to have two plot, Plot A with litter kept on the soil and Plot B with litter removed, was carried out in a Chinese fir plantation in South China. Soil samples were collected from the plots at the end of the experiment and analyzed for soil C and N and their isotopes (δ13C, δ15N), enzyme activity microbial biomass C and N (MBC, MBN), as well as N-acquiring enzymes. 【】Results show that Plot A significantly increased activity of soil N-acquiring enzyme activity (β-N- acetylglucosaminidase) and hence accelerated decomposition of soil organic matter. Therefore it was found that litter triggered decline of soil ammonium, nitrate N and DON, but barely affected soil MBC and MBN, which is probably because litter intensified the competition of microbes with plants for soil N, so soil microbe released more N-acquiring enzyme to meet their demand for N. In Plot A phenol oxidase and peroxidase increased in activity, while in Plot B they decreased in activity, which indicates that soil microbes tend to decompose Chinese fir litter rather than soil organic matter as energy to acquire N. In Plot A soil organic C and dissolvable organic C did not vary much, which indicates that litter does not have much impact on carbon in the soil. Litter did not have much impact on soil δ13C either, but did, by significantly increasing soil δ15N. Soil δ15N is an indicator of openness of soil N system. The higher the soil δ15N, the more open the soil N system, the more likely the occurrence of N loss, which indicates that litter accelerates soil N mineralization and loss. Besides, it was also found that in Plot B β-glucosidase and cellulolytic enzyme declined in activity, which indicates that litter removal may downregulate soil C recycling rate. 【】Based on the findings of the experiment, it could be concluded that 1) Litter significantly affects soil N, but not soil C, which indicates that soil N is more sensitive to changes in litter input than soil C; 2) Litter may alter the soil microbial environment of the Chinese fir plantation and enhance root growth and N uptake of the plant, thus leading to intensified competition between soil microbes and plants for soil N and decline of soil N in content; 3)As Chinese fir litter is high in C/N, soil microbes need to excrete more soil phenol oxidase and peroxidase activity to decompose fresh litter for energy to compete for soil N. Therefore, climate changes cause variation of litter production in Chinese fir forest, which in turn produces different impacts on recycling of soil C and N, thus disturbing balance of soil C and N in subtropical coniferous forests.
Litter treatment; Soil carbon and nitrogen isotopes; Enzyme activity
S714
A
10.11766/trxb201808060408
阮超越,劉小飛,呂茂奎,李曉杰,付作琴,任寅榜,黃俊,謝錦升. 杉木人工林凋落物添加與去除對土壤碳氮及酶活性的影響[J]. 土壤學報,2020,57(4):954–962.
RUAN Chaoyue,LIU Xiaofei,Lü Maokui,LI Xiaojie,FU Zuoqin,REN Yinbang,HUANG Jun,XIE Jinsheng. Effects of Litter Carbon,Nitrogen and Enzyme Activity in Soil under Chinese Fir[J]. Acta Pedologica Sinica,2020,57(4):954–962.
* 國家重點研發計劃項目(2016YFD0600204)和國家自然科學基金項目(U1405231)資助Supported by the National Key Research and Development Project of China(No. 2016YFD0600204)and the National Natural Science Foundation of China(No. U1405231)
,E-mail:jshxie@163.com
阮超越(1993—),男,安徽合肥人,碩士研究生,主要從事森林生態研究。E-mail: 2686309431@qq.com
2018–08–06;
2018–10–06;
2018–12–10
(責任編輯:盧 萍)
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
