澳系睡蓮花粉離體萌發及低溫保存研究
2020-09-02 06:54:35唐毓瑋龍凌云黃秋偉蘇群池昭錦盧家仕毛立彥*
熱帶作物學報 2020年7期
唐毓瑋 龍凌云 黃秋偉 蘇群 池昭錦 盧家仕 毛立彥 *

摘 ?要:為研究適宜睡蓮花粉離體萌發的培養基和低溫保存的方法,以睡蓮品種‘默笙和3份原種睡蓮的花粉為試驗材料,采用單因素、正交試驗設計,篩選出最適宜的培養基,觀測睡蓮花粉在不同溫度下保存的萌發率。結果表明,4種澳系睡蓮花粉的最適培養基的各組分濃度不同,花粉的生活力存在差異,澳洲原生淺色睡蓮與澳洲原生深色睡蓮的花粉生活力較高,分別為43.90%和45.63%;‘默笙睡蓮的花粉生活力次之,為31.84%;‘白巨睡蓮的花粉生活力較低,為9.73%;4種澳系睡蓮花粉均不耐貯藏,不同溫度下保存72 h后萌發率均為0。本研究結果可為睡蓮雜交育種的親本選擇、花粉的低溫保存研究提供參考。
關鍵詞:睡蓮;花粉生活力;離體培養;低溫保存中圖分類號:S31??????文獻標識碼:A
Pollen Germination in vitro and Cryopreservation Reserch of Anecphya Waterlily
TANG Yuwei1, LONG Lingyun1, HUANG Qiuwei1, SU Qun2, CHI Zhaojin1, LU Jiashi2, MAO Liyan1*
1. Guangxi Subtropical Crops Research Institute, Nanning, Guangxi 530001, China; 2. Flowers Research Institute, Guangxi Academy of Agricultural Sciences, Nanning, Guangxi 530007, China
Abstract: The pollens ofNymphaea‘Moshen and three orginalNymphaea accessions?were used as?the test material to study the optimal medium components for the pollen germinationin vitroofNymphaea and the method for prolonging pollen viability at low temperature.?The single factor and orthogonal design were used to select the most suitable medium, and observe the germination rate of the pollens preserved at different temperatures. The optimum medium concentration for the pollens of the four accessions?was different. The pollen viability varied among accessions. The pollen viability ofN. immutabilis subsp. Immutabilis blue formandN. immutabilis subsp. immutabilis purple form was 43.90% and 45.63% respectively, and that ofN.‘Moshen andN.‘Gigantea Albertde Lestangwas 31.84% and 9.73%, relatively. The pollen ofNymphaea was not tolerant to storage, and lost the viability after 72 hours of storage at different temperatures. The results of the study could provide references for parents selection and pollen cryopreservation in hybrid breeding ofNymphaea.
Keywords: Nymphaea; pollen viability;in vitro culture; cryopreservation
DOI: 10.3969/j.issn.1000-2561.2020.07.013
睡蓮是睡蓮科(Nymphaeaceae)睡蓮屬(Ny- mphaea)植物的總稱,屬于多年生草本植物[1],花色豐富,花態多樣,群體花期長,極具觀賞價值,在水生園林景觀中應用廣泛。睡蓮屬植物主要以生態學為基礎進行分類,可分為Anecphya、Brachyceras、Hydrocallis、Lotos、Nymphaea(orCastakua)5個亞屬[2-5]。Anecphya亞屬睡蓮俗稱為澳系睡蓮,因原種主要分布于澳大利亞而得名,澳系睡蓮適宜廣西地區種植,群體花期長達8~9個月。澳系睡蓮花態獨特,花瓣形似荷花,與其他亞屬的睡蓮形態有明顯差異,是重要的雜交育種材料[6]。
花粉生活力的強弱是影響雜交育種成敗的重要因素[7],授粉前對植株的花粉生活力進行評價,選擇花粉生活力較高的親本可以提高育種效率。花粉離體萌發法評價花粉生活力是目前較為準確和直觀的方法,在杜鵑、山茶、月季、海棠等植物的育種研究中均有應用[8-11]。關于睡蓮花粉的研究較少,楊夢飛等[12]與毛立彥等[13]的研究均表明,睡蓮花粉離體培養4 h后,萌發率趨于穩定;張慧會等[14]研究證明,30 ℃是睡蓮花粉離體萌發的最佳溫度。不同亞屬的睡蓮廣泛分布在暖溫帶至熱帶地區[15],單朵花期約4~5?d,雜交育種經常受地域、花期等因素制約。延長睡蓮花粉的生活力能有效解決花期不遇、異地授粉等問題,而低溫保存是延長花粉生活力的主要手段[16],因此低溫保存研究對于睡蓮的雜交育種有重要意義。目前,關于睡蓮花粉低溫保存方面的研究還未見報道,本研究以澳系睡蓮為試驗材料,進行花粉離體萌發試驗,篩選較佳的花粉離體萌發培養基,研究適宜的花粉保存溫度,為雜交育種的親本選擇、種質資源的保存提供參考依據[17]。
1??材料與方法
1.1材料
試驗地點位于廣西壯族自治區南寧市興寧區亞熱帶作物研究所睡蓮種質資源圃(108.38° E,22.87°N),屬于亞熱帶季風氣候,年平均氣溫21.6?℃,年平均降水量約1300 mm。試驗地為睡蓮種質資源圃,水體無浮萍、藻類等污染,水位平穩,水深約40 cm。
以4種澳系睡蓮為試驗材料,在花蕾時期做套袋處理,避免昆蟲攜帶花粉干擾試驗。隨機選取長勢良好、無病蟲害的植株,在花藥散粉時,采摘睡蓮花朵帶回實驗室內,將成熟花藥中的花粉均勻抖落在干燥培養皿中混勻(4種睡蓮花粉分別置于4個培養皿中),再將花粉分裝到若干支2?mL離心管中密封保存,用于花粉培養基篩選與花粉保存試驗。
1.2方法
1.2.1 ?花粉離體萌發法 ?取出保存花粉的2?mL離心管,使用移液槍向離心管加入700?μL培養液,置于30?℃培養室,500 lx的光照條件下自然培養,每隔1?h觀察一次,直至花粉萌發率穩定為止。將離心管輕輕搖勻,用移液槍吸取50?μL花粉液滴在載玻片上,使用Leica DM 2500顯微鏡進行鏡檢,使用Image-Pro Plus 6.0軟件統計視野內的花粉總數、萌發花粉數,每個玻片隨機選取5個不重復的視野,每個視野花粉數量不少于40粒,以花粉管長度大于花粉粒直徑視為萌發。培養基用水為雙蒸餾水,試驗重復3次。
花粉萌發率=(視野內萌發花粉數/視野內花粉總數)×100%
1.2.2 ?培養基篩選??以‘默笙睡蓮的新鮮花粉為材料進行單因素試驗設計,探討蔗糖、MgSO4、H3BO3、KH2PO4、CaCl2·2H2O對花粉萌發的影響,其意義在于縮小正交試驗因素水平的篩選范圍。參照張瑞等[18]的方法,以BK培養基[7](100?g/L蔗糖、100?mg/L H3BO3、300?mg/L CaCl2·2H2O、200?mg/L MgSO4、100?mg/L KH2PO4)作為基本培養基,在探討每個影響因素時,設置不同濃度梯度,其余因素均以BK培養基為假設最適值。
蔗糖設6個濃度梯度:50、100、150、200、250、300 g/L;MgSO4設6個濃度梯度:100、200、300、400、500、600 mg/L;H3BO3設6個濃度梯度:50、75、100、125、150、175 mg/L;KH2PO4設6個濃度梯度:100、200、300、400、500、600?mg/L;CaCl2·2H2O設6個濃度梯度:100、200、300、400、500、600?mg/L。以雙蒸餾水為培養基作陰性對照,以‘默笙睡蓮花露[19]作陽性對照(睡蓮花朵第1天盛開時,柱頭上會分泌花露,以接收昆蟲所攜帶的花粉)。
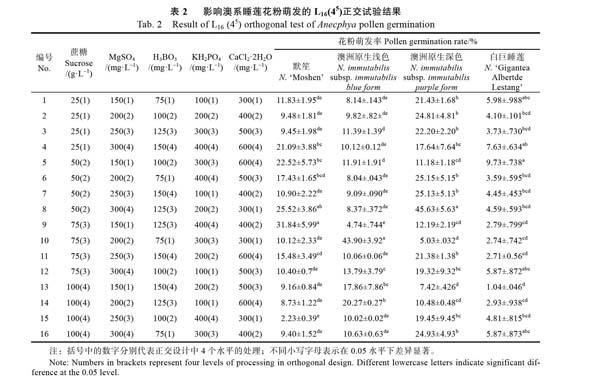
在單因素試驗的基礎上,以蔗糖(25、50、75、100 g/L)、MgSO4(150、200、250、300 mg/L)、H3BO3(75、100、125、150 mg/L)、KH2PO4(100、200、300、400 mg/L)、CaCl2·2H2O(300、400、500、600?mg/L)進行5因素4水平共16個組合L16(45)的正交試驗。參試材料進一步擴增至4種,每種材料進行3次重復試驗,依據花粉萌發率篩選出澳系睡蓮花粉的最佳萌發培養基。
1.2.3 ?花粉保存??以上述干燥的花粉作為試驗材料,每1 g分裝于2 mL離心管內密封保存,分別置于常溫(CK)、4、?20、?40、?70 ℃的密閉黑暗條件下保存。每隔24、48、72 h,取1支離心管的花粉進行離體培養萌發。分別選用4種澳系睡蓮對應的最佳培養基,觀測花粉的離體萌發率。試驗操作步驟、培養時間與條件、統計方法與上述方法相同。每種澳系睡蓮進行3次重復試驗。
1.3數據處理
采用Image-Pro Plus 6.0軟件統計每個視野睡蓮花粉的萌發率,采用Excel 2013 軟件進行數據統計與制圖,采用SPSS 18.0軟件進行方差分析。
2??結果與分析
2.1不同培養基組分對花粉萌發率的影響
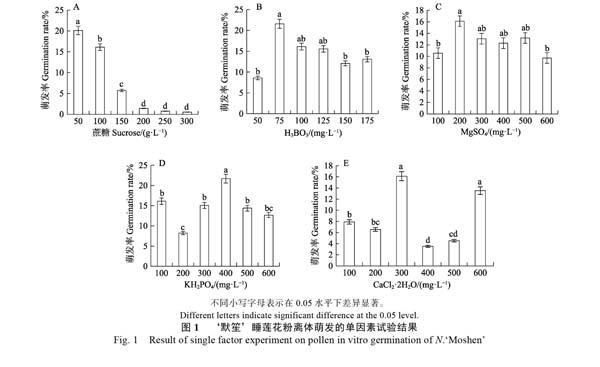
2.1.1 ?蔗糖濃度對花粉萌發的影響??陰性對照(雙蒸餾水處理)的花粉萌發率為0;陽性對照(睡蓮花露處理)的萌發率為14.33%。從圖1A可以看出,低濃度的蔗糖更適宜‘默笙睡蓮花粉的萌發,濃度在50 g/L時,花粉萌發率達到試驗范圍內最高值20.12%,高于陽性對照。隨著蔗糖濃度的提高,花粉萌發率顯著下降。方差分析結果表明,當蔗糖濃度達到200?g/L以上時,花粉萌發率較低,且無顯著差異。依據單因素試驗的結果,可將蔗糖濃度進一步精確調整至25、50、75、100 g/L進行正交試驗。
2.1.2 ?其余因子的濃度對花粉萌發的影響 ?與不同濃度的蔗糖處理相比,H3BO3、MgSO4、KH2PO4、CaCl2·2H2O的濃度對‘默笙花粉萌發的影響規律不明顯,不同濃度范圍表現出不同的結果。
不同小寫字母表示在0.05水平下差異顯著。
Different letters indicate significant difference at the 0.05 level.
由圖1B可知,H3BO3濃度為50 mg/L時,花粉萌發率最低(8.59%),當濃度上升至75?mg/L時,萌發率顯著提升至最高(21.59%),之后隨著H3BO3濃度的提高,萌發率緩慢下降。方差分析結果表明:當H3BO3濃度為75?mg/L時,花粉萌發率顯著高于50、150、175 mg/L的H3BO3處理。
由圖1C可知,MgSO4的濃度從100 mg/L提升至200?mg/L時,花粉萌發率顯著提升,從10.57%提升至最高(16.11%)。當濃度超過200?mg/L以上時,‘默笙的花粉萌發率隨著濃度的提高呈緩慢下降的趨勢。方差分析表明:當MgSO4濃度為200?mg/L時,‘默笙花粉的萌發率顯著高于100、600?mg/L MgSO4的處理。
由圖1D可知,當KH2PO4的濃度在200~600?mg/L時,花粉萌發率呈現中間高、兩頭低的分布趨勢。方差分析結果表明:當KH2PO4濃度為400?mg/L時,‘默笙睡蓮花粉的萌發率顯著高于其他處理,而100?mg/L處理的花粉萌發率與300、500、600 mg/L處理無顯著差異。
由圖1E可知,300、600 mg/L的CaCl2·2H2O適宜‘默笙花粉的萌發,萌發率分別為16.11%、13.52%。方差分析顯示:300、600?mg/L?CaCl2·2H2O的2個處理之間無顯著差異,且花粉萌發率顯著高于其他處理。
2.2正交試驗各處理對澳系睡蓮花粉萌發的影響
正交試驗設計組合及結果如表2所示,各處理對4種澳系睡蓮花粉萌發率的影響不同。其中,‘默笙的9號處理平均花粉萌發率最大,達到31.84%,顯著高于除8號處理外的其他處理的花粉萌發率(P<0.05);澳洲原生淺色睡蓮的10號處理與澳洲原生深色睡蓮的8號處理的花粉萌發率均顯著高于同類別中其他處理的平均花粉萌發率(P<0.05),平均花粉萌發率分別達到43.90%和45.63%;‘白巨睡蓮的5號處理花粉萌發率相對較高,但與1號、4號、8號、16號處理的平均花粉萌發率差異不顯著(圖2)。
注:括號中的數字分別代表正交設計中4個水平的處理;不同小寫字母表示在0.05水平下差異顯著。
Note: Numbers in brackets represent four levels of processing in orthogonal design. Different lowercase letters indicate significant difference at?the?0.05 level.
A:‘默笙花粉;B:澳洲原生淺色睡蓮花粉;C:澳洲原生深色睡蓮花粉;D:‘白巨睡蓮花粉
A: The pollen ofN. ‘Moshen; B:?The pollen ofN. immutabilis subsp. immutabilis blue form; C: The pollen ofN. immutabilis subsp. immutabilis purple form; D: The pollen ofN.‘Gigantea Albertde Lestang.
2.3不同保存條件對花粉離體萌發率的影響
由圖3可知,隨著花粉保存時間的增加,萌發率大幅度下降。?70?℃保存的花粉24 h以后花粉萌發率下降至0。常溫、4、?20、?40?℃保存的花粉72 h以后萌發率下降至0。花粉在4?℃條件下保存效果相對較好,24?h后的萌發率由31.84%下降至18.88%;48 h后,萌發率進一步降至4.32%。花粉在4?℃條件下保存24 h和48 h時的萌發率均高于其他溫度處理,表明‘默笙澳系睡蓮的花粉相對適合在4?℃條件下保存。
如圖4所示,‘白巨睡蓮花粉的初始萌發率較低,花粉隨著保存時間的增加,萌發率呈下降趨勢。?70?℃條件下保存24 h,萌發率為0;常溫條件下保存48?h,花粉無萌發;?4、?20、?40?℃條件下保存72?h,萌發率降至0。花粉在4?℃條件下保存24?h,萌發率小幅度下降,由9.73%降至9.05%;48?h后,萌發率大幅度降至2.61%。花粉在常溫、?20、?40、?70?℃下保存24?h,萌發率均大幅度下降,分別為2.71%、3.66%、2.08%、0。結果表明‘白巨睡蓮在4?℃條件下可保存24 h。
由圖5可知,澳洲原生淺色睡蓮的花粉隨著保存時間的增加,萌發率大幅度下降,不同溫度下保存24 h后,萌發率降至10%以下,48 h后均下降至2%以下,72 h后萌發率為0。花粉在4 ℃下處理效果相對略好,24 h后萌發率由43.90%下降至7.02%,略高于其他的溫度處理;48 h后,萌發率降至1.65%。花粉在?20?℃條件下保存24?h,萌發率由44.90%下降至7.02%,僅次于4℃的處理;48?h后,萌發率降至1.81%,略高于其他的溫度處理。
如圖6所示,澳洲原生深色睡蓮花粉隨著保存時間的增加,萌發率大幅下降。在常溫、?20、?40、?70?℃條件下保存48 h,萌發率下降至0。花粉在4?℃下保存效果相對略好,萌發率略高于其他溫度處理,但花粉生活力仍然大幅下降,保存至24 h,萌發率由45.63%下降至15.60%,48 h后萌發率進一步下降至1.52%,72?h后萌發率為0。
3討論
蔗糖是影響植物花粉萌發的重要因素,為花粉萌發和花粉管生長提供能源物質,同時維持著花粉細胞外界環境的滲透壓[20-22],H3BO3能增加花粉對糖的吸收、運轉和代謝,形成糖硼酸復合體[23-24],Ca2+的動態平衡影響花粉的萌發與花粉管的頂端生長[25]。本研究中,蔗糖的單因素和正交試驗結果表明,50、75?g/L的蔗糖配適宜的B3+、Ca2+、K+、Mg2+分別對4種澳系睡蓮花粉萌發有較好的促進作用。但是B3+、Ca2+、K+、Mg2+的單因素試驗與正交試驗結果不完全吻合。例如,在單因素試驗的BK培養基中,75 mg/L?H3BO3處理的‘默笙花粉萌發率最高,而在正交試驗的9號培養基中,125?mg/L H3BO3處理的‘默笙花粉萌發率更高,其原因可能是各因素存在交互作用,H3BO3與不同濃度的蔗糖、Ca2+、K+、Mg2+互作產生了不一樣的效應。毛立彥等[13]對‘保羅蘭睡蓮花粉的研究發現,蔗糖、CaCl2、H3BO3對睡蓮花粉萌發存在極顯著的交互作用;常海龍等[26]研究表明,在硼酸、蔗糖、硝酸鈣、硫酸鎂4個因素中,蔗糖對甘蔗花粉萌發的影響最大,硼酸次之,它們對花粉管的生長最為關鍵。4種澳系睡蓮花粉的最適培養基的各組分濃度不同,合理配比可以增加花粉的萌發率。通過花粉離體萌發法,可以直觀、有效地評價花粉生活力,在睡蓮的雜交育種過程中,對于父本材料的篩選有指導意義。在本研究中,‘白巨睡蓮的花粉生活力較低,不適宜作為父本育種材料。
低溫保存可以降低花粉呼吸作用、水分代謝、延長花粉生活力[27],是解決花期不育、異地授粉的主要手段。聶超仁等[28]研究表明,鐘花櫻花的花粉可在?20、?80?℃條件下保存150 d仍保持20%以上的萌發率;吳月亮等[29]研究證明,?80?℃超低溫條件保存文冠果花粉1年,花粉萌發率接近50%,而常溫條件下僅能保存15?d。本試驗中,4種澳系睡蓮的花粉在4?℃下保存相對較好,但72 h后花粉均喪失生活力,仍然無法解決睡蓮雜交育種中異地授粉、跨花期授粉等問題。
花粉含水量是影響花粉低溫保存時間長短的因素,在低溫冷凍過程中,含水量過高,在低溫下細胞內結冰過多,造成細胞膜受損,影響保存后的生活力[30]。趙嬋璞等[31]研究表明,未干燥的有斑百合花粉?80?℃保存480?d后萌發率下降至2.3%,經過8 h干燥處理的花粉(含水量21.3%)?80?℃貯藏720?d后,萌發率為20.7%。時婷婷等[32]發現,經干燥處理的‘索邦和‘西伯利亞百合新鮮花粉超低溫保存420?d后的花粉萌發率分別為60.12%和58.02%,而CK(未干燥的花粉)萌發率僅為3.20%和2.21%。本試驗沒有考慮澳系睡蓮花粉的含水量對花粉低溫保存的影響,有待后續進一步研究。
參考文獻
[1]?關克儉, 肖培根, 潘開玉, 等. 中國植物志: 第27卷[M]. 北京: 科學出版社, 1979: 8-11.
[2]?廖衛偉, 楊志娟, 朱天龍, 等. 睡蓮的植物學性狀及其分類[J]. 現代農業科技, 2016(24): 148, 154.
[3]?余翠薇, 陳煜初, 余東北, 等. 睡蓮5個亞屬花、葉、塊莖和基因組大小比較[J]. 浙江農業科學, 2017, 58(8): 1353-1356, 1361.
[4]?李子俊. 我國首批跨亞屬耐寒睡蓮育成[J]. 中國花卉盆景, 2013(10): 4-7.
[5]?劉義滿, Slearmlarp Wasuwat, 柯衛東, 等. 泰國睡蓮考察報告[J]. 中國園藝文摘, 2009, 25(3): 120-124.
[6]?黃國振. 睡蓮[M]. 北京: 中國林業出版社, 2009, 23-25, 45, 134-182.
[7]?Preston R E. The intrafloral phenology ofStreptanthustortuosus (Brassicaceae)[J]. American Journal of Botany, 1991, 78(8): 1044-1053.
[8]?陸 ?琳, 彭綠春, 宋 ?杰, 等. 不同高山杜鵑品種花粉活力測定及貯藏方法研究[J]. 山西農業科學, 2016, 44(2): 175-178.
- 賈文慶, 王少平, 李紀元. 大花紅山茶花粉形態特征和培養條件及其儲藏過程的生理動態分析[J]. 西北植物學報, 2015, 35(4): 754-760.
- 王嵐嵐, 游 ?捷, 俞紅強. 月季花粉離體萌發液體培養基組分的優化[J]. 河北農業大學學報, 2008(3): 42-45.
- 李千惠, 姜文龍, 徐同樂, 等. 日本海棠花粉最佳離體萌發培養基篩選及萌發過程研究[J]. 經濟林研究, 2018, 36(2): 154-160.
- 楊夢飛, 李怡鵬, 樓宇濤, 等. 培養液組分及培養時間對睡蓮花粉萌發和花粉管生長的影響[J]. 浙江農業科學, 2016, 57(10): 1689-1691.
- 毛立彥, 唐毓瑋, 謝振興, 等. ‘保羅蘭睡蓮花粉離體萌發及花粉管生長的研究[J]. 西南農業學報, 2019, 32(5): 1155-1161.
[14]?張慧會, 祝遵凌. 香水蓮花花粉活力測定最適培養條件分析[J]. 分子植物育種, 2020, 18(2): 553-560.
- 李淑娟, 尉 ?倩, 陳 ?塵, 等. 中國睡蓮屬植物育種研究
進展[J]. 植物遺傳資源學報, 2019, 20(4): 829-835.
[16]?王欽麗, 盧龍斗, 吳小琴, 等. 花粉的保存及其生活力測定[J]. 植物學通報, 2002(3): 365-373.
[17]?孫春麗, 潘延云. 擬南芥花粉活力的測定及其在花粉發育研究中的應用[J]. 植物學通報, 2008(3): 268-275.
[18]?張 ?瑞, 李 ?洋, 梁有旺, 等. 薄殼山核桃花粉離體萌發和花粉管生長特性研究[J]. 西北植物學報, 2013, 33(9): 1916-1922.
[19]?程 ?茵, 馬光耀, 趙 ?瑩, 等. 睡蓮花露主要成分及抗氧化作用[J]. 食品研究與開發, 2019, 40(8): 108-114.
[20]?黃 ?靜, 曹秋芬, 玉 ?平, 等. 培養基成分對南瓜離體花粉萌發的影響[J]. 中國西瓜甜瓜, 2004(3): 10-11.
[21]?杜紀紅, 葉正文, 蘇明申, 等. 桃花粉離體萌發和花粉管生長特性研究[J]. 西北植物學報, 2011, 31(1): 64-71.
[22]?鄧衍明, 葉曉青, 梁麗建, 等. 茉莉花粉離體培養萌發及花粉管生長特性研究[J]. 華北農學報, 2014, 29(5): 107-113.
[23]?Fang K F, Du B S, Zhang Q,et al. Boron deficiency alters cytosolic Ca2+concentration and affects the cell wall components of pollen tubes inMalus domestica[J]. Plant Biology, 2019, 21(2): 1-9.
[24]?朱江華, 呂芳德, 王 ?森, 等. 6個藍莓品種花粉特性和花粉育性的比較[J]. 經濟林研究, 2016, 34(2): 101-?108.
[25]?湯紅明, 徐冬青, 徐根娣, 等. 植物花粉萌發的研究進展[J]. 安徽農業科學, 2006(24): 6436-6438, 6440.
[26]?常海龍, 張 ?偉, 陳俊呂, 等. 甘蔗花粉離體萌發研究[J]. 熱帶作物學報, 2019, 40(10): 2068-2075.
- 史鋒厚, 范蓉蓉, 周 ?婷, 等. 垂絲海棠花粉貯藏特性研究[J]. 經濟林研究, 2013, 31(4): 190-194.
- 聶超仁, 況紅玲, 于靜亞, 等. 鐘花櫻花粉離體萌發培養及低溫保存技術研究[J]. 西南林業大學學報(自然科學), 2018, 38(1): 54-58.
- 吳月亮, 湯 ?鑫, 劉 ?迪, 等. 不同貯藏溫度下文冠果花粉離體萌發研究[J]. 沈陽農業大學學報, 2018, 49(5): 600-604.
- 張亞利, 尚曉倩, 劉 ?燕. 花粉超低溫保存研究進展[J]. 北京林業大學學報, 2006(4): 139-147.
- 趙嬋璞, 張泉衛, 史寶勝, 等. 有斑百合花粉低溫貯藏研究[J]. 河北農業大學學報, 2014, 37(1): 54-58.
[32]?時婷婷, 張桂玲, 溫四民, 等. 百合花粉超低溫保存方法研究[J]. 北方園藝, 2013(3): 64-66.
