貴州不同酒曲在刺梨酵素發(fā)酵菌劑制備中的應(yīng)用
2020-10-29 06:24:44王瑜李立郎解春芝楊禮壽楊娟萬麗英楊小生
食品與發(fā)酵工業(yè) 2020年20期
王瑜,李立郎,解春芝,楊禮壽,楊娟,萬麗英,楊小生*
1(貴州醫(yī)科大學(xué),藥用植物功效與利用國家重點(diǎn)實(shí)驗(yàn)室,貴州 貴陽,550014)2(貴州省中國科學(xué)院天然產(chǎn)物化學(xué)重點(diǎn)實(shí)驗(yàn)室,貴州 貴陽,550014)3(徐州工程學(xué)院 食品與生物工程學(xué)院,江蘇 徐州,221018)4(六盤水職業(yè)技術(shù)學(xué)院,貴州 六盤水,553001)
酵素是利用微生物的新陳代謝產(chǎn)物的作用,將發(fā)酵原料中的糖類、蛋白質(zhì)等大分子物質(zhì)轉(zhuǎn)化為小分子物質(zhì),富含多種生物活性物質(zhì),如醇、酸、醛、酯、維生素、酶、有機(jī)酸等的功能性產(chǎn)品[1]。酵素的發(fā)酵方式有自然發(fā)酵和純種發(fā)酵2種,其中常用的純種發(fā)酵劑優(yōu)勢微生物有乳酸菌、酵母菌及醋酸菌等,采用復(fù)合菌劑強(qiáng)化發(fā)酵,利用優(yōu)勢菌群在酵素食品發(fā)酵體系中快速構(gòu)建出有益的微生物菌群,抑制腐敗菌、致病菌等雜菌的生長,形成新的代謝產(chǎn)物和風(fēng)味物質(zhì)[2]。目前對于酵素中微生物菌群組成研究較少,MA等[3]采用單分子實(shí)時測序技術(shù)分析了日本和中國臺灣市場上銷售的酵素細(xì)菌組成及多樣性,鑒定了43個菌屬,優(yōu)勢菌群主要為嗜鉻桿菌屬、乳酸菌屬、分枝桿菌屬。凌空等[4]分離鑒定了果蔬酵素不同發(fā)酵周期中微生物,從3種不同發(fā)酵周期的果蔬酵素中分離篩選得到15株菌種。高慶超等[5]采用Illumina MiSeq高通量測序技術(shù)分析了不同發(fā)酵階段的黑果枸杞酵素微生物的群落組成及多樣性,發(fā)現(xiàn)黑果枸杞酵素在發(fā)酵期0~20 d細(xì)菌群落多樣性和豐富度相對較高,在發(fā)酵期30~50 d真菌群落多樣性和豐富度相對較高。通過研究鑒定了果蔬發(fā)酵中的主要微生物菌群,以獲得安全的發(fā)酵劑及為產(chǎn)品的穩(wěn)定性和風(fēng)味提供指導(dǎo)作用[6]。但采用酒曲提供微生物發(fā)酵制備酵素的研究相對較少,更無對微生物群落結(jié)構(gòu)和功能因子之間的相關(guān)性研究。
因此,本研究通過采集貴州省7個不同行政區(qū)域當(dāng)?shù)貍鹘y(tǒng)酒曲發(fā)酵刺梨酵素,運(yùn)用高通量測序方法對刺梨酵素細(xì)菌群落組成和多樣性進(jìn)行分析,比較微生物群落結(jié)構(gòu)差異,同時測定刺梨酵素中的主要功能成分與主要細(xì)菌種群做相關(guān)性分析,為刺梨酵素發(fā)酵劑的制備以及刺梨酵素中微生物群落結(jié)構(gòu)和功能成分之間的相關(guān)性提供依據(jù),為刺梨酵素的標(biāo)準(zhǔn)化開發(fā)提供一定的理論支撐。
1 材料與方法
1.1 材料與試劑
刺梨,貴州貴定敏子食品有限公司;酒曲,來源見1.3;纖維素酶,南寧龐博生物工程有限公司;DNA抽提試劑盒,F(xiàn)astDNA?Spin Kit for Soil;齊墩果酸(純度99%),中國藥品生物制品檢定所;蘆丁(純度94.47%)、VC(純度98%),貴州迪大生物科技有限責(zé)任公司。
1.2 儀器與設(shè)備
SKG-PB-936打漿機(jī),上海達(dá)瑞寶公司;BS-210S電子天平,北京賽多利斯儀器有限公司;EYELA旋轉(zhuǎn)蒸發(fā)儀,上海愛朗儀器有限公司;AG285微量分析天平,梅特勒-托利多儀器上海有限公司;GZX-9420MBE鼓風(fēng)烘箱,上海博迅實(shí)業(yè)有限公司;PAL-1 ATAGO糖度計,日本愛拓(愛拓)中國分公司;TCL-168離心機(jī),上海安亭科技儀器廠;UV-1800紫外分光光度計,上海精密科學(xué)儀器有限公司;Millipore-0026高效液相色譜儀,美國Thermo Fisher公司; ABSON MiFly-6微型離心機(jī),合肥艾本森科學(xué)儀器有限公司;Eppendorf 5430 R小型離心機(jī)、Eppendorf;NanoDrop2000超微量分光光度計,Thermo Fisher Scientific;DYY-6C電泳儀,北京市六一儀器廠; Illumina Miseq MISEQ測序儀,Illumina;QL-901旋渦混合器,海門其林貝爾儀器制造有限公司;TL-48R粉碎研磨儀,上海萬柏生物科技有限公司。
1.3 實(shí)驗(yàn)方法
1.3.1 酒曲的采集與刺梨酵素的制備
(1)酒曲的采集:根據(jù)貴州省行政區(qū)域的劃分,按照區(qū)域具有代表性的原則,以貴陽為中心,選擇貴州的東、西、南、北4個區(qū)域地點(diǎn),于2018年9~10月,分別在貴州省貴陽市花溪區(qū)(GY)、遵義赤水市(ZY)、銅仁地區(qū)德江縣(TR)、黔西南州興仁縣(QXN)、黔東南州凱里市(QDN)、黔南州龍里縣(QN)和畢節(jié)大方縣(BJ)7個行政區(qū)域采集傳統(tǒng)釀酒用高溫大曲用于制備刺梨酵素。
(2)刺梨酵素的制備:取新鮮刺梨洗凈去蒂,按照m(刺梨鮮果)∶m(純凈水)=1∶2打漿混合后,按照其總質(zhì)量的10%白糖,攪拌使白糖完全溶化,添加0.02%(質(zhì)量分?jǐn)?shù))纖維素酶,0.03%(質(zhì)量分?jǐn)?shù))貴州不同區(qū)域采集的酒曲粉末,酒曲使用前采用溫水活化,混合均勻后將其轉(zhuǎn)至陶瓷壇內(nèi),陶瓷壇采用100目紗布封口,在18~25 ℃下有氧發(fā)酵。待發(fā)酵3個月后,pH值降至3~4,糖度降至4%~5%,產(chǎn)品澄清透明,呈淡黃色,取表面上清液冷藏備用。
1.3.2 不同區(qū)域酒曲發(fā)酵刺梨酵素的功能成分檢測方法
刺梨酵素中功能成分總?cè)坪涂傸S酮采用紫外分光光度計,參考南瑩等[7]的方法測定,其中總?cè)埔札R墩果酸為標(biāo)準(zhǔn)品,總黃酮以蘆丁為標(biāo)準(zhǔn)品,VC采用高效液相色譜儀測定,色譜柱為Topsil C18色譜柱(4.6 mm×250 mm,5 μm);糖度采用糖度計測定。
1.3.3 樣品DNA提取與高通量測序分析
(1)樣本收集:取不同酒曲發(fā)酵的刺梨酵素液體樣品,離心后倒掉上清液,將菌體沉淀在-80 ℃下冷凍后,干冰保存狀態(tài)下送至深圳微科服科技有限公司進(jìn)行DNA提取與細(xì)菌菌群檢測。
(2)DNA抽提和聚合酶鏈?zhǔn)椒磻?yīng)(polymerase chain reaction,PCR)擴(kuò)增:根據(jù)FastDNA?Spin Kit for Soil試劑盒使用說明書進(jìn)行總DNA抽提,DNA濃度和純度利用NanoDrop 2000進(jìn)行檢測,利用2%(質(zhì)量分?jǐn)?shù))瓊脂糖凝膠電泳檢測DNA提取質(zhì)量;采用338F(ACTCCTACGGGAGGCAGCAG)和806R(GGACTACHVGGGTWTCTAAT)引物對V3-V4可變區(qū)進(jìn)行PCR擴(kuò)增,PCR試驗(yàn)采用TransGen AP221-02:TransStart Fastpfu DNA Polymerase;20 μL反應(yīng)體系:擴(kuò)增程序?yàn)?5 ℃預(yù)變性3 min,27個循環(huán)(95 ℃變性30 s,55 ℃退火30 s, 72 ℃延伸45 s),最后72 ℃延伸10 min。擴(kuò)增體系為20 μL,4 μL 5×FastPfu緩沖液,2 μL 2.5 mmol/L dNTPs,0.8 μmol/L引物(5 μmol/L),0.4 μL FastPfu聚合酶; 0.2 μL BSA; 10 ng DNA模板[8]。
(3)Illumina Miseq測序:使用2%(質(zhì)量分?jǐn)?shù))瓊脂糖凝膠回收PCR產(chǎn)物,利用AxyPrep DNA Gel Extraction Kit (Axygen Biosciences, Union City, CA, USA) 進(jìn)行純化,Tris-HCl洗脫,2%瓊脂糖電泳檢測。利用QuantiFluor-ST (Promega, USA) 進(jìn)行檢測定量。根據(jù)Illumina MiSeq平臺 (Illumina, San Diego, USA)標(biāo)準(zhǔn)操作規(guī)程將純化后的擴(kuò)增片段構(gòu)建PE 2×300的文庫。利用Illumina公司的Miseq PE300平臺進(jìn)行測序。
1.4 數(shù)據(jù)處理
利用 QIIME 2.0 軟件包,對測序片段進(jìn)行分類操作單元(operational taxonomic units,OUT)聚類,OUT代表序列與Greengene數(shù)據(jù)庫進(jìn)行比對分析,獲取 OUT 對應(yīng)的分類單元及其相應(yīng)的豐度信息;計算 Shannon index和Observed-OTU index度量指數(shù),評估樣品中細(xì)菌菌群的豐富度和多樣性;綜合應(yīng)用 PCA 分析、LEfSe 分析、ANOVA 方差分析等方法分析樣品中的微生物菌群;應(yīng)用冗余分析(redundancy analysis,RDA) 分析關(guān)聯(lián)菌群結(jié)構(gòu)和刺梨酵素功能成分總?cè)啤⒖傸S酮、VC及糖度的相關(guān)性,利用Excel 2010和SPSS Statistics 20.0等統(tǒng)計軟件進(jìn)行數(shù)據(jù)分析和作圖。
2 結(jié)果與分析
2.1 瓊脂糖凝膠電泳檢測DNA提取結(jié)果
采用338F(ACTCCTACGGGAGGCAGCAG)和806R(GGACTACHVGGGTWTCTAAT)引物對V3-V4可變區(qū)進(jìn)行PCR擴(kuò)增,PCR擴(kuò)增產(chǎn)物電泳條帶均在750 bp 左右,條帶長度和亮度較好,表明DNA含量較高,可上機(jī)檢測進(jìn)行后續(xù)實(shí)驗(yàn)。
2.2 不同酒曲發(fā)酵的刺梨酵素微生物OTU數(shù)目分析
通過檢測不同區(qū)域酒曲發(fā)酵的刺梨酵素共22個樣品,得到有效序列1 139 057條,平均每個樣品51 775條。圖1是展現(xiàn)不同區(qū)域間共有和特有OTU數(shù)目的花瓣圖,由圖1可知,7個區(qū)域共有的OTU數(shù)目為46,貴陽特有的OTU數(shù)目為500,畢節(jié)特有的OTU數(shù)目為840,遵義特有的OTU數(shù)目為277,銅仁特有的OTU數(shù)目為1 125,黔西南特有的OTU數(shù)目為618,黔南特有的OTU數(shù)目為559,黔東南特有的OTU數(shù)目為433。
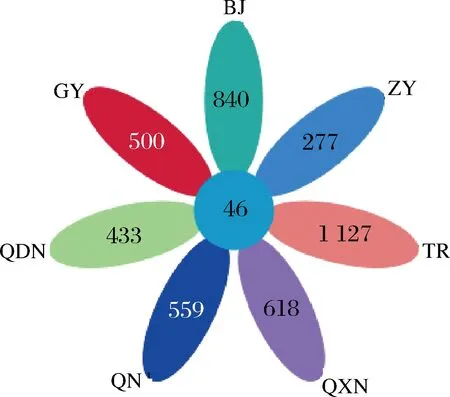
圖1 不同區(qū)域間共有和特有OTU數(shù)目花瓣圖Fig.1 Common and unique OTU number of petals among different regions
2.3 不同酒曲發(fā)酵的刺梨酵素微生物群落的α-多樣性分析
圖2是畢節(jié)、貴陽、黔東南、黔南、黔西南、銅仁、遵義(BJ、GY、QDN、QN、QXN、TR、ZY)7個地區(qū)間α-多樣性盒形圖,更直觀顯示不同區(qū)域間α-多樣性差異。α-多樣性指數(shù)是對樣品中物種多樣性的分析,包含樣品中的物種組成豐富度和均勻度2個因素,通常用Shannon指數(shù)來評估某個樣本的物種多樣性,Shannon值越大,表明樣本的多樣性越復(fù)雜;而用Observed-OTU指數(shù)來評估樣本的物種豐富度呈正相關(guān),Observed OTU指數(shù)是指樣本中實(shí)際測定得到的OTU數(shù)量,衡量樣品中OTU豐富度的指數(shù)。

A-Shannon指數(shù)統(tǒng)計分析;B-Obsrved-OTU指數(shù)統(tǒng)計分析;C-Shannon指數(shù)組間差異檢驗(yàn);D-Obsrved-OTU指數(shù)組間差異檢驗(yàn)圖2 不同區(qū)域酒曲發(fā)酵刺梨酵素微生物α-多樣性盒形圖Fig.2 Microbial life of Roxburgh rose Jiaosu in different regions - diversity box-like diagram
圖2-A反映的是物種的多樣性,圖2-B反映的是物種的豐富度。由圖2-A可知,不同區(qū)域的微生物多樣性差異較大,Shannon結(jié)果表明,銅仁(TR)地區(qū)的酒曲發(fā)酵制備的刺梨酵素具有最高的微生物多樣性,而遵義(ZY)地區(qū)酒曲發(fā)酵制備的刺梨酵素微生物多樣性相對較低;從豐富度方面分析,不同區(qū)域酒曲發(fā)酵的刺梨酵素都具有較高的豐富度,通過比較Observed OTU指數(shù),銅仁(TR)地區(qū)的酒曲發(fā)酵制備的刺梨酵素具有最高的微生物豐富度,而黔東南(QDN)地區(qū)酒曲發(fā)酵制備的刺梨酵素微生物豐富度相對較低。由圖2-C和圖2-D可知,不同區(qū)域細(xì)菌多樣性和豐富度組間存在一定的差異性,但無顯著性差異(P>0.05),然而,有研究發(fā)現(xiàn)不同地區(qū)的酒曲微生物群落構(gòu)成及多樣性存在明顯差異,且酒曲的基本理化指標(biāo)與微生物菌群結(jié)構(gòu)有一定的相關(guān)性[9],可能是地域特有的環(huán)境和溫度導(dǎo)致的差異性。任海偉等[10]研究發(fā)現(xiàn)溫度會直接影響微生物菌群的動態(tài)變化和多樣性,室溫更有利于提高乳酸發(fā)酵強(qiáng)度,抑制腐敗菌生長,使有機(jī)酸和有機(jī)組分得到優(yōu)化重組,實(shí)現(xiàn)較高貯存品質(zhì)。溫度對微生物的多樣性影響顯著,銅仁地區(qū)常年溫度較高,四季溫差變化較小,適宜微生物生長,造成該地區(qū)較高的微生物多樣性和豐富度,因此選擇適宜穩(wěn)定的發(fā)酵溫度是提高酵素品質(zhì)的關(guān)鍵。
2.4 不同酒曲發(fā)酵的刺梨酵素細(xì)菌微生物群落組成分析
2.4.1 不同區(qū)域酒曲發(fā)酵刺梨酵素在門水平上優(yōu)勢菌群結(jié)構(gòu)組成分析
分析不同區(qū)域酒曲發(fā)酵的刺梨酵素菌群在門水平上分布前10的菌群有變形菌門(Proteobacteria)、厚壁菌門(Firmicutes)、擬桿菌門(Bacteroidetes)、放線菌門(Actinobacteria)、疣微菌門(Verruco-microbia)、酸桿菌門(Acidobacteria)、梭桿菌門(Fusobacteria)、綠彎菌門(Chloroflexi)、螺旋體門(Spirochaetes)、未知細(xì)菌(unspecified_Bacteria),其中相對豐度>1%的菌群有3種,即變形菌門(Proteobacteria)、厚壁菌門(Firmicutes)、擬桿菌門(Bacteroidetes)。母應(yīng)春等[11]研究了貴州花溪、貴州和安順3個地區(qū)酒曲的微生物多樣性,發(fā)現(xiàn)厚壁菌門、變形菌門、放線菌門的相對豐度在3個地區(qū)酒曲中均較高;另外陳玲等[12]以濃香型白酒大曲為研究對象,基于16S rRNA基因?yàn)槟康钠危捎梦⑸镏讣y圖譜分析技術(shù)16S rDNA克隆文庫法和高通量測序法分析了大曲中細(xì)菌微生物群落的組成,發(fā)現(xiàn)大曲中的微生物主要分布于厚壁菌門(Firmicutes)、變形菌門(Proteobacteria)、放線菌門(Actinobacteria)和藍(lán)藻菌門(Cyanobacteria/Chloroplast)4個菌門,由此可知刺梨酵素中微生物菌群的組成與發(fā)酵酒曲密切相關(guān)。進(jìn)一步分析不同區(qū)域中各優(yōu)勢菌群所占的比例,其中畢節(jié)(BJ)地區(qū)酒曲發(fā)酵的刺梨酵素中變形菌門(Proteobacteria)占52.3%,厚壁菌門(Firmicutes)占37.9%,擬桿菌門(Bacteroidetes)占7.2%;貴陽(GY)地區(qū)酒曲發(fā)酵的刺梨酵素中變形菌門(Proteobacteria)占69.2%,厚壁菌門(Firmicutes)占26.7%,擬桿菌門(Bacteroidetes)占2.2%;黔東南(QDN)地區(qū)酒曲發(fā)酵的刺梨酵素中變形菌門(Proteobacteria)占71.8%,厚壁菌門(Firmicutes)占19.5%,擬桿菌門(Bacteroidetes)占7.0%;黔南(QN)地區(qū)酒曲發(fā)酵的刺梨酵素中變形菌門(Proteobacteria)占67.0%,厚壁菌門(Firmicutes)占28.2%,擬桿菌門(Bacteroidetes)占2.0%;黔西南(QXN)地區(qū)酒曲發(fā)酵的刺梨酵素中變形菌門(Proteobacteria)占62.9%,厚壁菌門(Firmicutes)占28.9%,擬桿菌門(Bacteroidetes)占2.7%;銅仁(TR)地區(qū)酒曲發(fā)酵的刺梨酵素中變形菌門(Proteobacteria)占58.4%,厚壁菌門(Firmicutes)占27.7%,擬桿菌門(Bacteroidetes)占7.4%;遵義(ZY)地區(qū)酒曲發(fā)酵的刺梨酵素中變形菌門(Proteobacteria)占69.2%,厚壁菌門(Firmicutes)占27.6%,擬桿菌門(Bacteroidetes)占2.0%。綜上可知,采用酒曲發(fā)酵的刺梨酵素中優(yōu)勢菌群為變形菌門(Proteobacteria)、厚壁菌門(Firmicutes)、擬桿菌門(Bacteroidetes)、放線菌門(Actinobacteria),其中以變形菌門(Proteobacteria)的相對豐度最高,每個區(qū)域都占到了50%以上,不同區(qū)域各個菌群有一定差異性,但在門水平上差異不顯著。
2.4.2 不同區(qū)域酒曲發(fā)酵刺梨酵素在屬水平上優(yōu)勢菌群結(jié)構(gòu)組成分析
在屬水平上,7個區(qū)域酒曲發(fā)酵的刺梨酵素中分布前10的細(xì)菌屬為假單胞菌屬(Pseudomonas)、醋酸桿菌屬(Acetobacter)、乳酸菌屬(Lactobacillus)、未知的胞菌科屬(unspecified_Aeromonadaceae)、葡糖醋桿菌屬(Gluconacetobacter)、腸球菌屬(Enterococcus)、未知的乳桿菌屬(unspecified_Lactobacillaceae)、未知的腸桿菌屬(unspecified_Enterobacteriaceae)、未知的梭菌屬(unspecified_Clostridiales)、擬桿菌屬(Bacteroides)、普氏菌屬(Prevotella),其相對豐度如圖3所示。其中相對豐度>10%的菌屬有畢節(jié)(BJ)的假單胞菌屬(Pseudomonas)占28.6%,醋酸桿菌屬(Acetobacter)占17.9%,乳酸菌屬(Lactobacillus)占25.2%,貴陽(GY)的假單胞菌(Pseudomonas)占40.7%,醋酸桿菌屬(Acetobacter)占17.4%,乳酸菌屬(Lactobacillus)占14.8%,黔東南(QDN)地區(qū)的假單胞菌屬(Pseudomonas)占13.0%,醋酸桿菌屬(Acetobacter)占53.8%,乳酸菌屬(Lactobacillus)占8.9%,黔南(QN)地區(qū)的假單胞菌屬(Pseudomonas)占36.2%,醋酸桿菌屬(Acetobacter)占21.7%,乳酸菌屬(Lactobacillus)占20.9%,黔西南(QXN)地區(qū)的假單胞菌(Pseudomonas)占38.3%,醋酸桿菌屬(Acetobacter)占8.4%,乳酸菌屬(Lactobacillus)占11.9%,銅仁(TR)地區(qū)的假單胞菌(Pseudomonas)占43.3%,醋酸桿菌屬(Acetobacter)占6.6%,乳酸菌屬(Lactobacillus)占14.1%,遵義地區(qū)(ZY)的假單胞菌(Pseudomonas)占24.8%,醋酸桿菌屬(Acetobacter)占34.7%,乳酸菌屬(Lactobacillus)占22.2%。
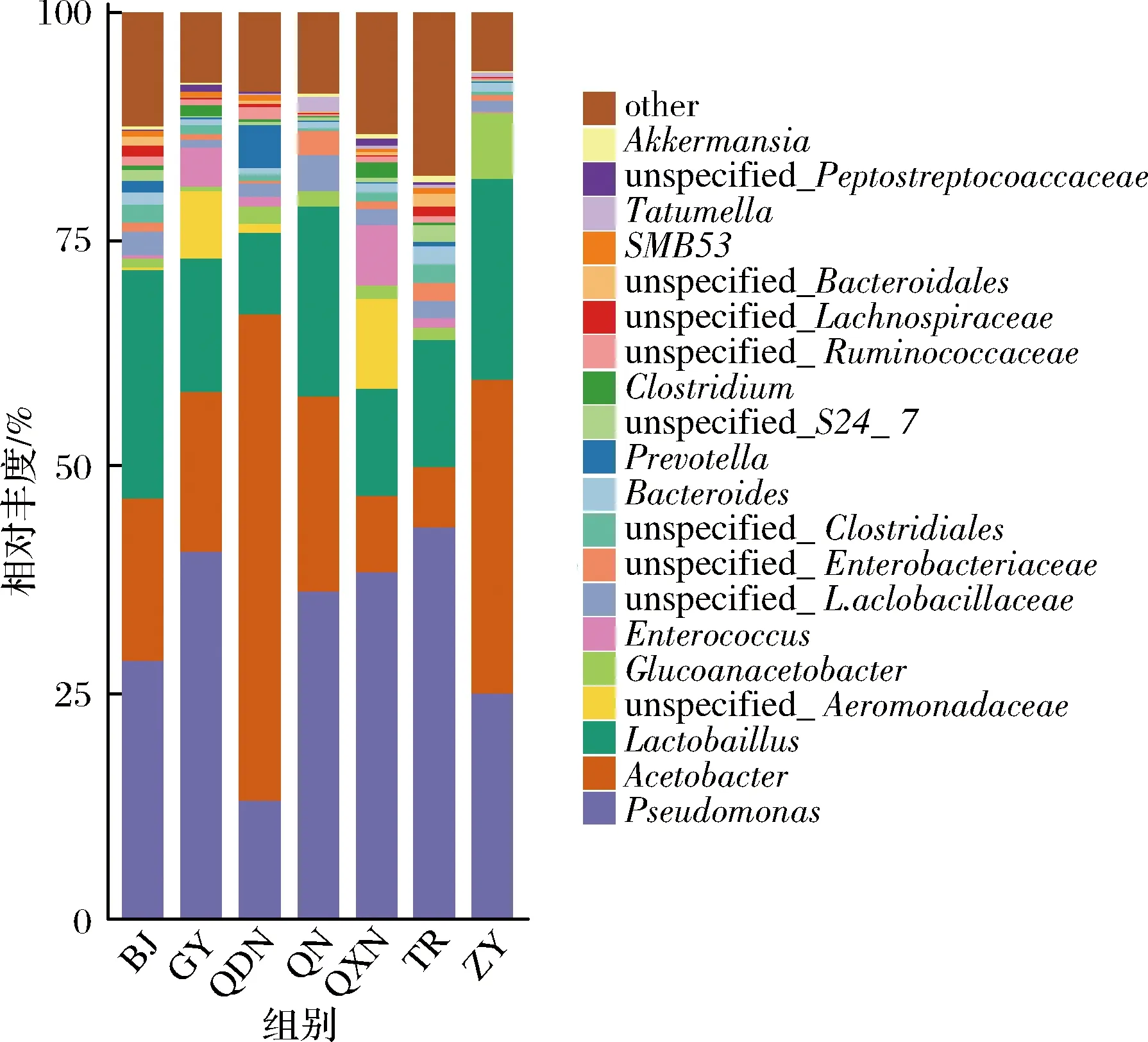
圖3 不同區(qū)域酒曲發(fā)酵刺梨酵素菌群屬水平分布圖Fig.3 Horizontal distribution of flora genus of Roxburgh rose Jiaosu in different regions
由以上分析可知,在屬水平上不同地區(qū)酒曲發(fā)酵的刺梨酵素中已鑒定到的優(yōu)勢菌群為假單胞菌屬(Pseudomonas)、醋酸桿菌屬(Acetobacter)、乳酸菌屬(Lactobacillus)。酵素發(fā)酵過程是由醇和酸共同參與轉(zhuǎn)化的過程,在酶和微生物的作用下將糖類或蛋白質(zhì)直接轉(zhuǎn)化為醇或酸類[13],厚壁菌門中的乳酸菌屬能夠產(chǎn)酸且在酸度較高的環(huán)境下生長[14],主要參與了酵素發(fā)酵的生物轉(zhuǎn)化,與酵素中有機(jī)酸的形成密切相關(guān),乳酸菌是一類具有提高機(jī)體免疫力,促進(jìn)營養(yǎng)物質(zhì)吸收,維持胃腸道菌群平衡的益生菌[15]。研究表明不同乳酸菌種發(fā)酵對桂圓肉中游離態(tài)、結(jié)合態(tài)酚類物質(zhì)含量、單體酚組成及其抗氧化活性都有影響,乳酸菌能提高桂圓肉中游離態(tài)酚類物質(zhì)含量,降低結(jié)合態(tài)酚類物質(zhì)含量,同時提高其抗氧化能力[16],因此,乳酸菌代謝對酵素功能的表達(dá)至關(guān)重要。醋酸桿菌屬是酵素發(fā)酵過程中的一大類產(chǎn)酸細(xì)菌,了解醋酸菌屬的組成及發(fā)酵特性有助于合理控制酵素發(fā)酵進(jìn)程,有效提高酵素的風(fēng)味品質(zhì)[17]。另外醋酸桿菌屬(Acetobacter)與葡糖醋桿菌屬(Gluconacetobacter)也是酵素發(fā)酵過程中產(chǎn)生菌蓋的關(guān)鍵菌群[28],醋酸和乳酸是酵素風(fēng)味的主要成分,二者存在的酸性條件可影響有機(jī)酸、功能成分的穩(wěn)定性[19]。
2.5 不同酒曲發(fā)酵的刺梨酵素功能成分與菌群屬水平物種RDA分析
分析不同區(qū)域酒曲發(fā)酵刺梨酵素的功能成分,結(jié)果如表1所示。
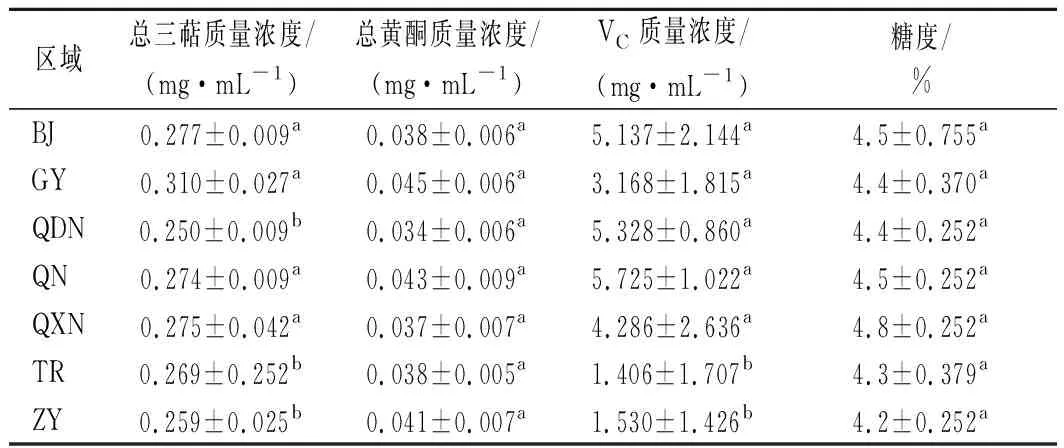
表1 不同區(qū)域酒曲發(fā)酵刺梨酵素功能成分含量表Table 1 Contents of functional components of Roxburgh rose Jiaosu in different regions
由表1可知,功能成分總?cè)坪蚔C含量存在顯著性差異(P<0.05),而不同酒曲發(fā)酵的刺梨酵素中總黃酮和糖度的含量差異不顯著。在相同的發(fā)酵條件下,發(fā)酵底物相同,而發(fā)酵劑不同,導(dǎo)致發(fā)酵產(chǎn)物中功能成分含量的改變,因此發(fā)酵劑中微生物與發(fā)酵產(chǎn)物之間可能存在相關(guān)性, 典型相關(guān)分析(canonical correlation analysis,CCA)/RDA排序如圖4所示。
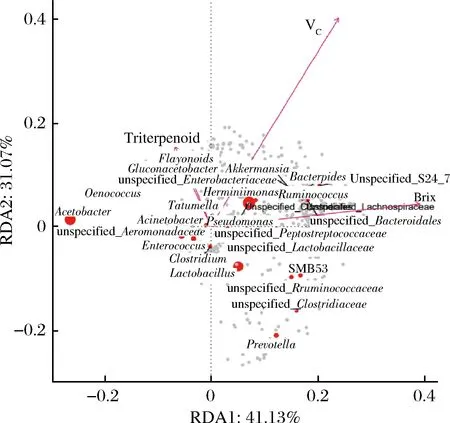
圖4 不同酒曲發(fā)酵的刺梨酵素菌群屬水平物種CCA/RDA排序圖Fig.4 CCA/RDA sequence of flora genus of Roxburgh rose Jiaosu in different regions
RDA物種排序圖內(nèi),環(huán)境因子用箭頭表示,箭頭連線的長度代表某個環(huán)境因子與群落分布和種類分布間相關(guān)程度的大小(解釋方差的大小),箭頭越長,說明相關(guān)性越大,反之越小[20]。箭頭連線和排序軸的夾角代表某個環(huán)境因子與排序軸的相關(guān)性大小,夾角越小,相關(guān)性越高,反之越低。環(huán)境因子之間的夾角為銳角時表示2個環(huán)境因子之間呈正相關(guān)關(guān)系,鈍角時呈負(fù)相關(guān)關(guān)系。每個點(diǎn)代表一個物種,點(diǎn)越大,對應(yīng)物種豐度越高,灰色點(diǎn)代表豐度較低的物種[21],通過功能因子用箭頭連線的長度代表某個環(huán)境因子與群落分布和種類分布間相關(guān)程度的大小[22]。由圖4可知,刺梨酵素中的功能成分與菌群結(jié)構(gòu)之間存在相關(guān)性,通過圖中原點(diǎn)大小判斷醋酸桿菌屬(Acetobacter)、假單胞菌屬(Pseudomonas)、乳酸桿菌屬(Lactobacillus)、葡糖醋桿菌屬(Gluconacetobacter)與刺梨酵素中功能成分的相關(guān)性較大,通過箭頭連線的長度可知VC與菌群的相關(guān)程度最大,功能因子總?cè)坪涂傸S酮之間夾角為銳角,表明2個指標(biāo)之間具有協(xié)同效應(yīng)。而糖度與總?cè)啤⒖傸S酮之間夾角為鈍角,表明糖度與功能因子總?cè)坪涂傸S酮之間呈負(fù)相關(guān)性,由以上結(jié)果可知,刺梨酵素中細(xì)菌菌群組成與其含有的主要功能成分之間存在相關(guān)性。
3 結(jié)論
本試驗(yàn)采用高通量測序方法對貴州不同區(qū)域酒曲發(fā)酵的刺梨酵素細(xì)菌群落組成和多樣性進(jìn)行分析,比較不同地區(qū)酒曲發(fā)酵刺梨酵素的微生物群落結(jié)構(gòu)差異,探究刺梨酵素中微生物群落與功能因子的相關(guān)性。通過對菌群多樣性分析,貴州省7個區(qū)域酒曲發(fā)酵的刺梨酵素共得到有效序列1 139 057條,平均每個樣品51 775條,α-多樣性分析表明,銅仁(TR)地區(qū)酒曲發(fā)酵制備刺梨酵素的微生物多樣性和豐富度相對其他地區(qū)均較高,而遵義(ZY)地區(qū)多樣性相對較低,黔東南(QDN)地區(qū)豐富度相對較低。從菌群結(jié)構(gòu)組成上分析,醋酸桿菌屬(Acetobacter)、乳酸菌屬(Lactobacillus)作為酵素中的主要微生物;通過RDA分析得出刺梨酵素中細(xì)菌菌群組成與其含有的主要功能成分之間存在相關(guān)性,其中與功能因子相關(guān)的菌群按照相關(guān)性依次為醋酸桿菌屬(Acetobacter)、假單胞菌屬(Pseudomonas)、乳酸桿菌屬(Lactobacillus)、葡糖醋桿菌屬(Gluconacetobacter)。以上結(jié)論能夠?yàn)槿蘸髲木魄泻Y選益生菌作為酵素發(fā)酵劑提供借鑒與參考,同時也為酒曲發(fā)酵刺梨酵素的可行性提供了理論依據(jù),后期將進(jìn)一步探究不同酒曲在刺梨酵素發(fā)酵過程中微生物的動態(tài)變化,為刺梨酵素的標(biāo)準(zhǔn)化開發(fā)提供數(shù)據(jù)支撐。
