仿刺參—中國明對蝦—斑節對蝦混養下水體微生物組成和多樣性
2020-11-19 10:09:04關曉燕蔣經偉田甲申周遵春王旭達
水產科學 2020年6期
關曉燕,王 擺,蔣經偉,田甲申,董 穎,周遵春,王旭達
( 1.大連海事大學,環境科學與工程學院,遼寧 大連 116023;2.遼寧省海洋水產科學研究院,遼寧省海洋水產分子生物學重點實驗室,遼寧 大連 116023 )
水產養殖環境具有極高細菌豐度,這些細菌在水產養殖過程中發揮著重要作用[1],其中有的能夠加速殘留餌料和糞便的分解[2],有的能夠將非離子氨、亞硝態氮、硫化氫等有毒的物質轉化為低毒性或無毒性的物質[3],有的會導致養殖動物死亡[4]等。因此,研究水產養殖環境細菌群落的多樣性和組成結構,在養殖過程中提高養殖效率、減少病害發生、控制環境污染等方面均有十分重要的意義。近年來,隨著分子生物學技術的不斷發展,越來越多的研究者開始使用高通量測序技術研究水產養殖環境中的不可培養微生物以及環境微生物的整體組成結構,如凡納濱對蝦(Litopenaeusvannamei)生長過程中腸道菌群的變化規律、養殖池塘原核和真核微生物的組成及分類學差異和疾病發生與其腸道菌群結構的關系等研究[5-7]。截至目前,針對水產養殖環境細菌群落的研究依然大多集中在單一養殖物種模式,對多物種混合養殖系統關注較少。
遼東灣位于渤海東北部,作為遼河和大凌河的入海口,適宜的氣候條件以及河流帶來的豐富營養物質使其成為了發展海水養殖產業的絕佳區域。由于具有環境友好、營養物質可循環利用、經濟效益高等特點,多品種的混合養殖目前已經成為了遼東灣地區主要的養殖模式[8]。其中,仿刺參(Apostichopusjaponicus)—中國明對蝦(Fenneropenaeuschinensis)—斑節對蝦(Penaeusmonodon)(A.F.P)的混合養殖作為新興的一種養殖模式,在遼東灣地區得到了大力的發展和推廣,雖然該養殖模式在水產品產出和經濟效益方面取得了成功,但是對于這種養殖模式的基礎理論研究,尤其是養殖環境微生物群落罕有研究。鑒于此,筆者通過高通量測序手段,分析仿刺參—中國明對蝦—斑節對蝦混合養殖過程中池塘水體細菌群落組成及其變化規律,旨在為更好地理解和改進該養殖模式提供理論指導。
1 材料與方法
1.1 養殖池塘情況
試驗池塘位于遼寧省遼東灣沿岸某水產養殖場,池塘養殖面積約3.33 hm2,養殖期間水深1.5 m。仿刺參采取輪放輪捕方式,每年5月投苗,放苗量525 kg/hm2;每年春季或秋季收獲1次,捕大留小,年產量約750 kg/hm2;中國明對蝦放苗時間為5月中旬,放苗密度為30 000尾/hm2;斑節對蝦放苗時間為5月底,放苗密度為7500尾/hm2。養殖期間不投餌。中國明對蝦和斑節對蝦養殖進行到9月底捕撈收獲,仿刺參養殖繼續進行。
1.2 樣品采集和DNA提取
2018年6—9月,每月15日前后參照水生微生物采集方法[9]采集養殖池塘水體樣本1次,每次固定在池塘的中心采集表面下方10 cm處的水樣5 L,樣品采集后,立即置于冰盒中并于12 h內運送至實驗室。
1.3 樣品處理與DNA提取
水體樣品運送到實驗室之后,立即使用0.22 μm微孔濾膜真空過濾收集其中的細菌細胞。將過濾后的濾膜保存在5 mL離心管中,液氮速凍3~5 min,轉移至冰箱中-80 ℃保存備用,接著將濾膜充分剪碎(使樣品表面菌群更好地與裂解液接觸)置于1.5 mL滅菌離心管中,加入裂解液,使其釋放DNA,隨后使用PowerWater?DNA Isolation Kit (MOBIO,USA)按照說明書的步驟提取濾膜上富集細菌的DNA,提取得到的DNA用1%的瓊脂糖凝膠電泳檢測其完整性,同時使用NanoPhotometer?Classic Launched (IMPLEN,GER)測定其含量和純度,所有提取成功的DNA樣品-20 ℃保存備用。
1.4 16S rRNA擴增子測序
應用含有Illumina接頭和barcode序列的引物341F-806R[10]擴增所有提取DNA樣品中的16S rRNA基因的V3~V4可變區,所有的PCR反應體系均為30 μL,其中包括15 μL的Phusion?High-Fidelity PCR Master Mix (New England Biolabs)、0.2 μmol/L的引物和1 μL的模板DNA。PCR反應過程如下:98 ℃預變性1 min;之后98 ℃變性10 s,50 ℃退火30 s,72 ℃延伸30 s,30個循環。用2%的瓊脂糖凝膠電泳檢測PCR產物,使用QIAquick Gel Extraction Kit (QIAGEN, GER)通過切膠回收的方式純化400~450 bp規格的PCR產物主帶,純化后的PCR產物應用NanoPhotometer?Classic Launched (IMPLEN, GER)測定其含量和純度,之后每一個樣品對應的PCR產物根據其含量進行等量混合構建測序文庫。使用Qubit?2.0 Fluorometer (Thermo Scientific)和Agilent Bioanalyzer 2100 system檢測文庫的質量,最后應用IlluminaHiseq測序平臺以PE250 bp策略對文庫進行測序。
1.5 數據處理
首先對測序得到的原始序列進行質量控制,去除質量得分低于20、含有模糊堿基、引物出現錯配或者長度少于150 bp的序列,剩余的序列根據barcode識別其所屬的樣品隨后去除barcode和引物序列。應用FLASH軟件[11]將重疊區域大于10 bp并且沒有任何堿基錯配的paired-end序列拼接為tags,使用QIIME軟件[12]對tags進行聚類,相似度大于97%的tags歸屬于同一個運算分類單元(OTU),應用默認的參數從每個運算分類單元中挑選其代表序列,之后使用Ribosomal Database Project (RDP) classifier根據Greengenes數據庫進行分類學注釋,最后根據每一個運算分類單元所包含的序列數目及其分類學注釋結果計算各樣品細菌群落中從門至屬水平的組成及其相對豐度。
1.6 生物信息學分析
應用PAST v3.0軟件[13]計算各個樣品細菌群落的α-多樣性指數,包括Chao1、ACE、Shannon和Simpson指數。使用QIIME計算各樣品細菌群落結構的加權Unifrac距離并進行非加權配對平均法聚類(UPGMA)以評估各樣品細菌群落的整體差異。使用R語言平臺的“ggplot2”軟件包繪制各樣品從門至屬水平主要物種的相對豐度柱形圖。
2 結 果
2.1 仿刺參—中國明對蝦—斑節對蝦養殖池塘水體中細菌群落的分類學注釋和α-多樣性分析
應用高通量測序技術研究了仿刺參—中國明對蝦—斑節對蝦混養系統完整養殖周期的細菌群落變化,結果見圖1。在配對序列拼接和質量控制之后,共得到了191 956條tags用于細菌群落組成的分析,各樣品tags的數目為43 228~52 975條,平均每個水體樣品tags 47 989條。這些測序得到的tags按照97%的相似性被聚類為運算分類單元(每個運算分類單元代表1個不同的菌屬),共聚類得到1332個運算分類單元,各樣品運算分類單元的數目為753~1003,平均每個水體樣品含有882個運算分類單元。根據每個運算分類單元的代表序列,對其進行了從門至屬水平的分類學注釋,這些檢測到的細菌分別屬于35個門、70個綱、110個目、187個科和336個屬。
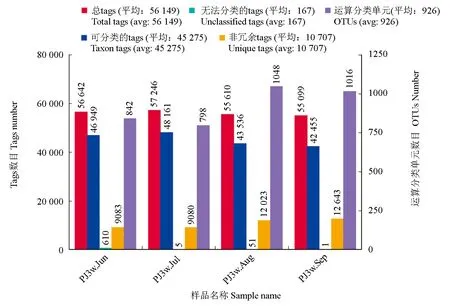
圖1 仿刺參—中國明對蝦—斑節對蝦養殖池塘水體微生物群落高通量測序得到的序列數目和運算分類單元數目Fig.1 Statistics of sequences and OTUs of different samples in the polyculture of sea cucumber with Chinese shrimp and tiger shrimp, A.F.P. systemPJ3w. Jun, PJ3w. July, PJ3w. Aug, PJ3w. Sep分別指6、7、8、9月采集的仿刺參—中國明對蝦—斑節對蝦養殖池塘水樣,下同.PJ3w refers to the water samples from the A.F.P. polyculture system, and Jun, Jul, Aug, and Sep refer to the sampling time, respectively.
仿刺參—中國明對蝦—斑節對蝦養殖池塘水體樣品細菌群落的α-多樣性指數結果見表1。在計算得到的α-多樣性指數中,Chao1和ACE用于評估樣品細菌群落的物種豐富度,而Shannon和Simpson用于評價細菌群落的多樣性。物種豐富度是指細菌群落中含有多少種細菌,其并不計算各個細菌在群落中所占的比例[14],多樣性指數在計算的過程中同時考慮了物種的豐富度和各細菌在群落中的豐度[15]。無論是豐富度還是多樣性指數,6月和7月的水體樣本均低于8月和9月,即養殖后半段(8—9月)的池塘水體具有更高的細菌種類和更均一的細菌豐度分布。
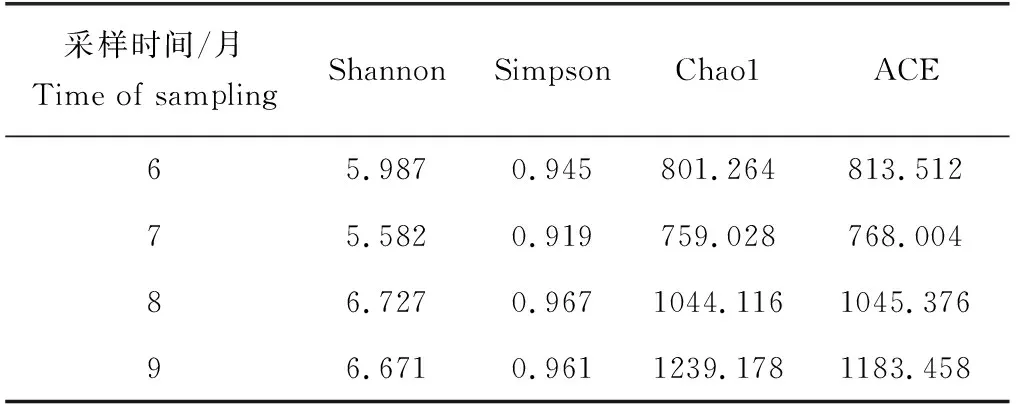
表1 仿刺參—中國明對蝦—斑節對蝦混養池塘水體微生物群落α-多樣性指數表Tab.1 α-diversity indices of microbial communities in water in the A.F.P. polyculture system
2.2 仿刺參—中國明對蝦—斑節對蝦養殖池塘水體中細菌群落結構隨時間的變化
2.2.1 仿刺參—中國明對蝦—斑節對蝦養殖水體中細菌群落結構聚類和主要門水平微生物相對豐度
通過基于加權Unifrac距離的UPGMA聚類評估了不同時間樣品細菌群落結構的整體差異,結果見圖2。試驗結果表明,養殖前半段(6—7月)樣品的細菌群落聚為一類,而養殖后半段(8—9月)樣品的細菌群落聚為另外一類,這與α-多樣性指數的結果相同,說明養殖前半段和養殖后半段池塘水體細菌群落具有明顯的差異。
此外,仿刺參—中國明對蝦—斑節對蝦養殖水體中主要細菌為變形菌門 (51.5%),其次為放線菌門(23.6%)、擬桿菌門(11.3%)和厚壁菌門(9.1%)。養殖前半段水環境中變形菌門的豐度要明顯高于養殖后半段,在6—7月樣品中變形菌門的豐度分別為57.7%和61.9%,而在8—9月樣品中則降至47.3%和39.2%。放線菌門在仿刺參—中國明對蝦—斑節對蝦池塘水體中的豐度僅次于變形菌門,其在6月至9月的樣品中,豐度依次為21.9%、15.1%、26.6%和30.8%,呈現出先降低后升高的趨勢。擬桿菌門是豐度排名第3的細菌門,在6月至9月的樣品中,其豐度分別為12.0%、14.9%、8.9%和9.5%,與放線菌門相反,擬桿菌門的豐度呈先升高后降低的趨勢。此外,在養殖后半段的水體中,厚壁菌門的豐度明顯高于養殖前半段,其豐度由6月至9月依次為5.4%、5.3%、11.1%和14.6%。
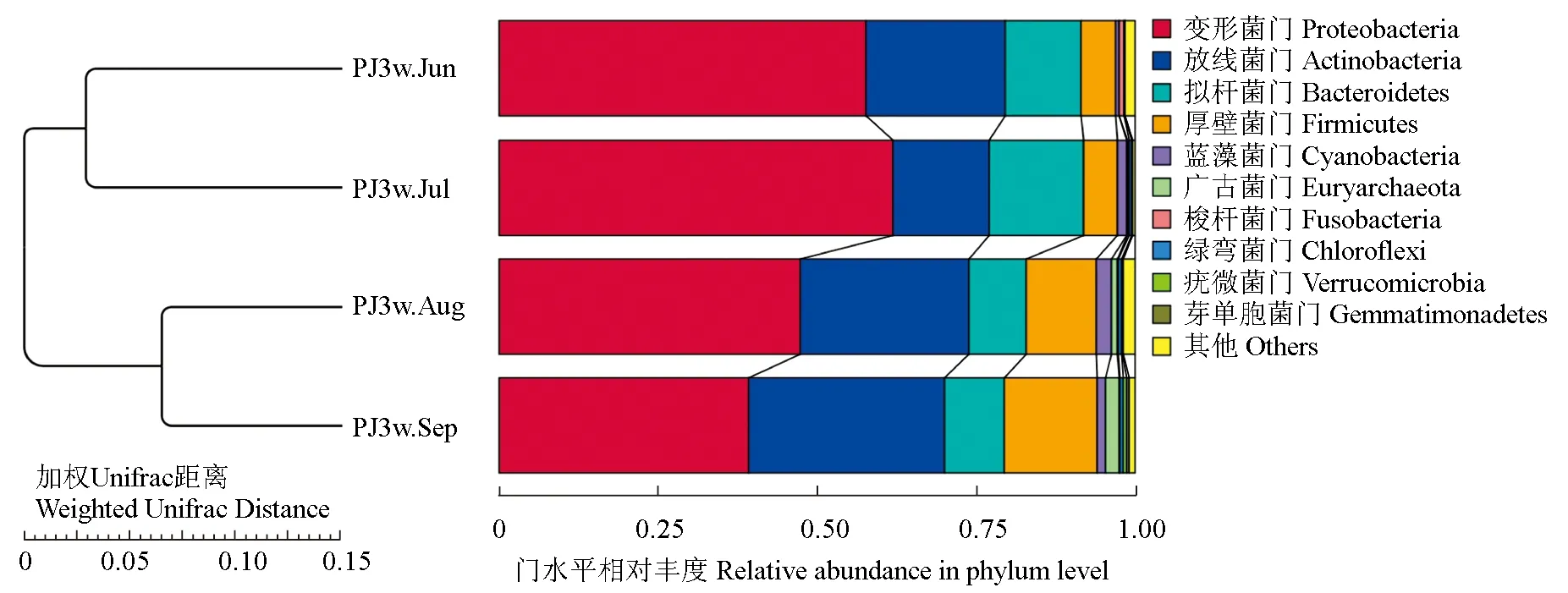
圖2 仿刺參—中國明對蝦—斑節對蝦養殖水體中細菌群落結構UPGMA聚類(左)和主要門水平微生物相對豐度(右)Fig.2 UPGMA cluster (left) and relative abundance of bacteria at phylum level (right) in water in A.F.P. polyculture
2.2.2 仿刺參—中國明對蝦—斑節對蝦養殖水體中細菌群落主要綱水平微生物相對豐度
對仿刺參—中國明對蝦—斑節對蝦池塘水體綱水平微生物相對豐度進行分析,試驗結果見圖3。試驗結果表明,α-變形菌綱和γ-變形菌綱是變形菌門的主要組成成分,養殖前半段α-變形菌綱的豐度較高,其在6月和7月樣品中的豐度分別為30.0%和40.5%,而在8月和9月樣品中則只有23.9%和24.7%,這也是變形菌門養殖前半段豐度較高的原因。γ-變形菌綱與α-變形菌綱不同,其在9月的水體中豐度最低,只有10.0%,而在其余時間的樣品中豐度為18.4%~22.3%。放線菌門中酸微菌綱在養殖后半段豐度突然上升,從6月和7月的1.9%與2.1%上升至8月和9月的13.4%與18.2%。擬桿菌門中黃桿菌綱的豐度從6月樣品中的8.1%降至其他時間樣品中的約3%,而7月樣品中1種未被鑒定的擬桿菌綱富集(9.9%,其他時間只有1.2%~2.9%),二者共同造成了擬桿菌門豐度前高后低的變化規律。
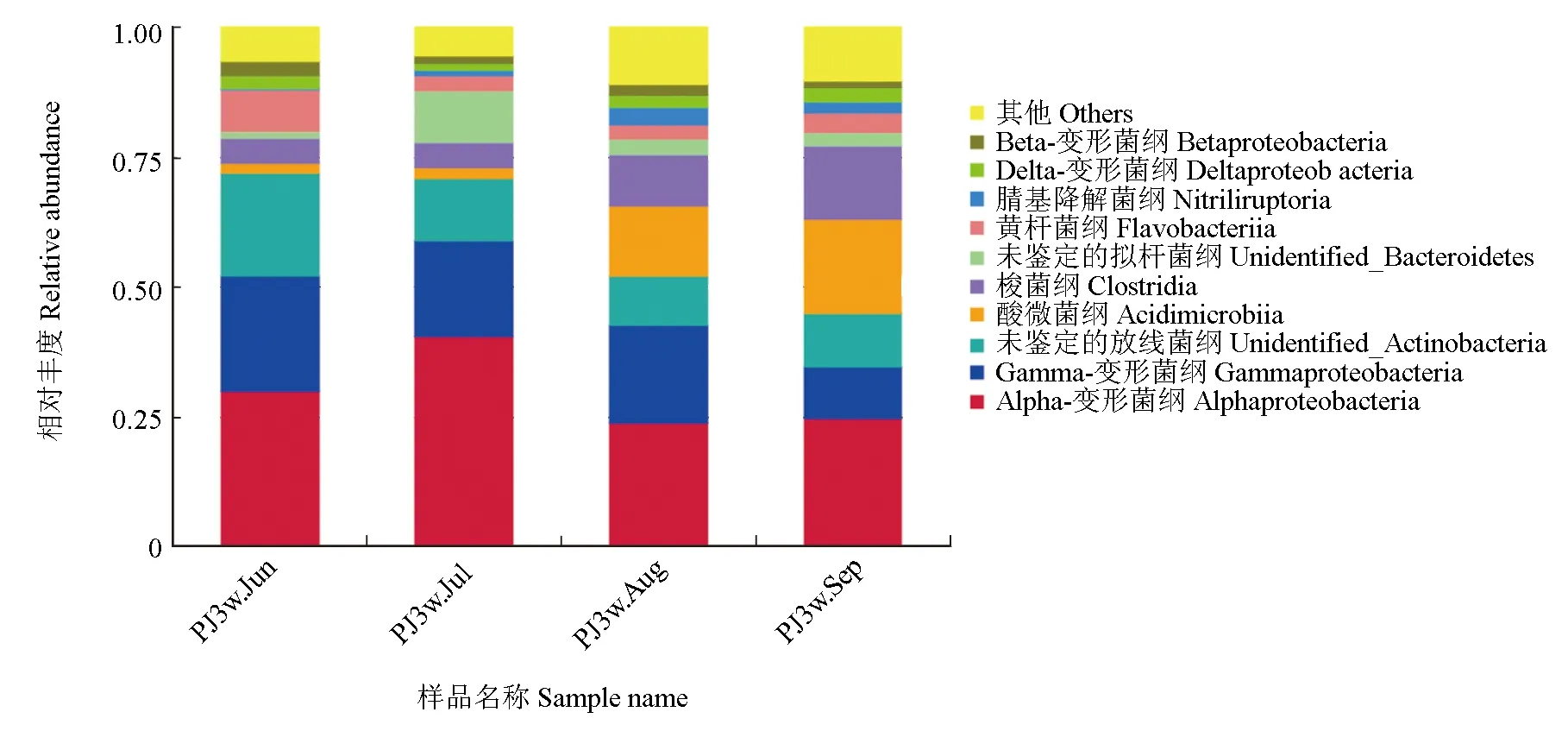
圖3 仿刺參—中國明對蝦—斑節對蝦養殖水體中細菌群落主要綱水平微生物相對豐度Fig.3 The relative abundance of bacteria at class level in water in A.F.P. polyculture
2.2.3 仿刺參—中國明對蝦—斑節對蝦養殖水體中細菌群落主要屬水平微生物相對豐度
對仿刺參—中國明對蝦—斑節對蝦養殖水體中細菌群落主要屬水平微生物相對豐度進行分析,試驗結果見圖4。試驗結果表明,α-變形菌綱中豐度最高的屬為遠洋桿菌屬(Pelagibacter),其在7月樣品中所占的豐度明顯高于其他時間(7月樣品為23.0%,其余時間僅約為10%)。養殖前3個月γ-變形菌綱的豐度較高,但不同時間水體中γ-變形菌綱的組成并不相同,6月樣品中的γ-變形菌綱主要是Halioqlobus,其豐度為7.7%,而在其余時間只有不到1%;7月樣品中的γ-變形菌綱主要為假交替單胞菌(Pseudoalteromonas),其豐度為7.8%,在其余時間只占0.1%~1.3%;8月樣品中的γ-變形菌綱主要為假單胞菌(Pseudomonas)和不動桿菌(Acinetobacter),豐度分別為4.5%和4.0%,而這2種細菌在其余時間樣品中均未檢出。
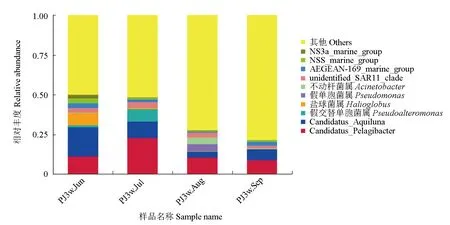
圖4 仿刺參—中國明對蝦—斑節對蝦養殖水體中細菌群落主要屬水平微生物相對豐度Fig.4 The relative abundance of bacteria at genus level in water in A.F.P. polyculture
3 討 論
3.1 不同養殖品種混養水體中微生物群落組成和多樣性差異
近年來,不同養殖品種混養水體中微生物群落的組成成分的研究已有較多報道。Li等[16]測定了江蘇省連云港市沿海地區對蝦、螃蟹和貝類混養池塘水體中微生物群落組成,證實該養殖模式中主要微生物依次為變形菌門、綠彎菌門、放線菌門、厚壁菌門和酸桿菌門;林玉榮等[17]測定了福建省莆田市海水蝦蛤混養池塘水體中微生物群落組成,結果顯示,該養殖模式中以放線菌門、變形菌門和擬桿菌門為主;Zhao等[18]分析了遼東灣沿海斑節對蝦、海蜇(Rhopilemaesculenta)和菲律賓蛤仔(Ruditapesphilippinarum)養殖池塘水體微生物群落組成,發現變形菌門、放線菌門、酸桿菌門和嗜熱菌門是占優勢地位的微生物;Gao等[19]在遼東灣另一種中國明對蝦、海蜇、縊蟶(Sinonovaculaconstricta)混養池塘的水體微生物群落中測定發現,豐度排名前三的微生物分別為變形菌門、放線菌門和擬桿菌門。本試驗結果表明,在仿刺參—中國明對蝦—斑節對蝦混養水體的微生物群落組成中,主要的微生物依次為變形菌門、放線菌門、擬桿菌門和厚壁菌門。上述研究表明,在不同的水產養殖系統中,變形菌門均為豐度最高的微生物,但其他微生物的種類和豐度會受到養殖物種、養殖操作方式以及養殖地域等因素的影響,從而導致不同養殖系統中微生物群落的組成結構各不相同。
此外,在仿刺參—中國明對蝦—斑節對蝦混養水體中,微生物群落聚為兩類,養殖前半段和養殖后半段的微生物群落組成結構和物種豐富度及多樣性具有明顯差異。本試驗前期針對遼東灣地區斑節對蝦—海蜇—菲律賓蛤仔混養進行了研究[18],該養殖模式與仿刺參—中國明對蝦—斑節對蝦類似,均是在每年5月底投放蝦苗直至9月收獲,該養殖模式水體微生物群落的多樣性和組成結構的變化規律與本研究不同,在斑節對蝦—海蜇—菲律賓蛤仔養殖模式中,不同時間水體微生物群落的組成結構各不相同,同時,其水體微生物群落的豐富度和多樣性指數沒有明顯的養殖時間特征,其豐富度指數在養殖過程中沒有明顯變化,多樣性指數也只是在8月有所提升。以上研究結果表明,養殖池塘內的微生物群落組成及其變化規律,具有養殖品種特異性。
3.2 仿刺參—中國明對蝦—斑節對蝦養殖模式微生物群落組成參與控制水體富營養化
在海水池塘養殖過程中,養殖水體的富營養化以及藻類的大量繁殖可能對養殖動物的生存造成威脅。在仿刺參—中國明對蝦—斑節對蝦混養水體中,遠洋桿菌屬是豐度最高的微生物屬。據報道,遠洋桿菌是海洋環境中一種常見的典型貧營養微生物,主要生活在低營養水平的環境中,以溶解性的有機碳和有機氮為底物進行生長和繁殖[20-21]。本試驗結果顯示,在養殖進行到7月時,仿刺參—中國明對蝦—斑節對蝦混養水體中遠洋桿菌的豐度達到了峰值,預示著此時養殖水體中營養物質含量可能較低。此外,假交替單胞菌在仿刺參—中國明對蝦—斑節對蝦養殖進行到7月時也出現了明顯的富集。眾所周知,假交替單胞菌主要附著于高等真核生物,尤其是藻類等海洋植物表面[22],能夠分泌胞外酶、毒素、抗生素、多糖等多種胞外生物活性物質,抑制藻類孢子萌發、降解藻類細胞、分解植物多糖[23],因此,假交替單胞菌在仿刺參—中國明對蝦—斑節對蝦養殖池塘內的富集同樣有助于過量藻類的降解。Aquiluna是一種含有肌動蛋白的光能異養型微生物,有研究檢測到在藻類大量暴發的水產養殖池塘中,通常也伴隨著Aquiluna豐度的明顯上升[24]。在仿刺參—中國明對蝦—斑節對蝦養殖過程中,其水體微生物群落中Aquiluna的豐度呈現持續下降的趨勢,這一結果也間接表明了仿刺參—中國明對蝦—斑節對蝦養殖池塘中的微生物群落組成能夠有效地抑制藻類的大量繁殖。然而,需要說明的是,雖然從仿刺參—中國明對蝦—斑節對蝦養殖過程中水體微生物群落的變化能夠推測到養殖過程中富營養化相關的信息,但是由于仿刺參—中國明對蝦—斑節對蝦混養是一種新興的養殖模式,目前關于該養殖模式的相關研究非常有限,尚未見有關于養殖過程中池塘水環境營養物質及藻類水平的相關報道,有待于后續針對仿刺參—中國明對蝦—斑節對蝦養殖模式進行更為深入和全面的研究。
3.3 仿刺參—中國明對蝦—斑節對蝦養殖模式微生物群落參與水質凈化過程
假單胞菌具有很強的有機物分解和轉化能力,因此其在環境污染生物修復中的應用已成為近年來的研究熱點問題[25-26]。例如,銅綠假單胞菌(P.aeruginosa)YY24可以提高凡納濱對蝦養殖過程中的存活率和質量增加率,同時還能夠凈化水質,提高水體pH,降低氨氮、硝態氮、亞硝態氮的含量[27]。銅綠假單胞菌OP2能夠分解不溶于水的含磷物質,使其成為可溶性磷,更加便于養殖動物吸收,還能有效去除水體中殘留的氨氮和亞硝態氮[28]。本試驗結果顯示,在仿刺參—中國明對蝦—斑節對蝦養殖后期,水體中假單胞菌屬豐度的上升,可能有利于養殖動物的存活和生長及凈化水質。
此外,梭狀芽孢桿菌(Clostridium)的豐度在仿刺參—中國明對蝦—斑節對蝦養殖后半段出現了明顯富集。據報道,丁酸梭狀芽孢桿菌(C.butyricum)可以有效地幫助日本沼蝦(Macrobrachiumnipponense)抵抗亞硝態氮所造成的壓力,同時能夠明顯提升其超氧化物歧化酶和過氧化氫酶的活性[29],這預示著仿刺參—中國明對蝦—斑節對蝦養殖水體中的梭狀芽孢桿菌可能有助于提升養殖對蝦抵抗亞硝態氮的能力,保證養殖后半段對蝦健康快速生長。
除了梭狀芽孢桿菌,在仿刺參—中國明對蝦—斑節對蝦模式養殖后半段微酸菌綱豐度出現了明顯的上升,水環境中微酸菌綱的分布及其豐度要明顯高于其他環境介質[30]。有研究發現,部分微酸菌綱的微生物在重金屬污染的酸性環境中大量繁殖,并且參與利用鐵離子還原介導的氨氧化過程[31]。這說明在仿刺參—中國明對蝦—斑節對蝦養殖后半段富集的微酸菌綱可能與假單胞菌共同參與水環境中氨氮的轉化過程,對于去除養殖池塘內過量的氮、凈化水體、提高養殖物種存活率具有十分重要的意義。
4 結 論
本試驗通過16S rRNA擴增子的高通量測序考察了仿刺參—中國明對蝦—斑節對蝦混合養殖池塘水體細菌群落的多樣性及豐度變化,發現其中主要的細菌為變形菌門、放線菌門、擬桿菌門、和厚壁菌門。養殖前半段和后半段池塘水體細菌群落具有明顯的差異。后半段,池塘水體微生物群落的豐富度和多樣性均出現了明顯的上升;變形菌門的豐度和擬桿菌門的豐度在養殖前半段要明顯高于養殖后半段,放線菌門的豐度呈先降后升的趨勢,而厚壁菌門在養殖后半段的豐度要明顯高于養殖前半段。
