神農(nóng)架華山松人工林土壤呼吸特征及其受枯立木分布的影響
2020-11-23 06:05:40黃金蓮崔鴻俠唐萬鵬雷靜品周文昌胡琛馬國飛
生態(tài)環(huán)境學(xué)報 2020年9期
黃金蓮,崔鴻俠,唐萬鵬,雷靜品,周文昌,胡琛,馬國飛
1.中國林業(yè)科學(xué)研究院林業(yè)所,北京 100091;2.湖北省林業(yè)科學(xué)研究院,湖北 武漢 430075;3.神農(nóng)架國家公園管理局,湖北 神農(nóng)架 442421
土壤是陸地生態(tài)系統(tǒng)的主要碳庫(Schimel,1995;Li et al.,2019),土壤呼吸是土壤碳庫向大氣中釋放CO2的主要途徑,其微小變化都會對陸地生態(tài)系統(tǒng)和全球碳平衡產(chǎn)生重大影響(Wu et al.,2014)。在全球氣候變化大背景下,國內(nèi)外廣泛開展了森林、草原、濕地等不同土地利用類型土壤呼吸特征的監(jiān)測與研究(胡樂寧等,2012;溫軍等,2014;Li et al.,2019)。關(guān)于土壤呼吸影響機制的研究,主要集中在土壤溫度、土壤含水量等環(huán)境因子對土壤呼吸的影響(竹萬寬等,2018;趙冰清等,2013;譚炯銳等,2019),以及模擬氮沉降、添加生物炭、改變凋落物量、提升土壤溫度等人工控制試驗對土壤呼吸的影響等(Palviainen et al.,2018;孫海燕等,2018;高強等,2015;Gao et al.,2014)。但受土壤呼吸影響因素的復(fù)雜性和不確定性,嚴重限制了對土壤呼吸特征及其控制機理的深入認識。
林木死亡是森林生態(tài)系統(tǒng)中的一個重要的生態(tài)過程,發(fā)生在林木生長發(fā)育的不同階段,也是森林演替與更新的必經(jīng)階段(Teodosiu et al.,2012)。林木死亡過程中,林內(nèi)會形成一定數(shù)量的枯立木,其形成原因有很多,其中病蟲害干擾是重要因素之一(仇智虎等,2014)。枯立木的存在及其伐除方式等能夠?qū)ι指屡c演替產(chǎn)生顯著影響,進而影響森林的結(jié)構(gòu)與功能,是森林的重要組成部分。此外,枯立木是森林生態(tài)系統(tǒng)的不可或缺的碳庫來源,對森林生態(tài)系統(tǒng)碳平衡與碳循環(huán)具有重要影響。目前國外有關(guān)枯立木的研究主要是對儲量與分解率的測定,以及評價生態(tài)系統(tǒng)的健康狀況和森林經(jīng)營措施(Franklin et al,1987;Oberle et al.,2018;Farahat et al.,2018;Kuuluvainen et al.,2017),而國內(nèi)對于枯立木的研究主要集中于枯立木的形成與分布格局、環(huán)境影響、儲量特征及分解規(guī)律等方面(仇智虎等,2014;譚雪蓮等,2019;張璐等,2018;李翾然等,2019),但很少有研究將枯立木對森林土壤的影響聯(lián)系起來,關(guān)于枯立木數(shù)量及分布、分解程度與土壤呼吸的關(guān)系研究還未見報道。
林窗是森林生態(tài)系統(tǒng)中的一種中小尺度干擾,是促進森林更新、養(yǎng)分循環(huán)、功能提高的重要推動力(王家華等,2006)。以往關(guān)于林窗對森林生態(tài)系統(tǒng)地上結(jié)構(gòu)與過程影響的研究報道比較多,如林窗對森林小氣候、物種組成及生物多樣性、林下更新、生物量分配等方面的影響(劉文杰等,2000;Qin et al.,2011;鮮駿仁等,2007);近年來,林窗對細根與凋落物分解(Lin et al.,2015),以及土壤養(yǎng)分和微生物群落(周義貴等,2014)等方面影響的研究逐漸增多,但已有研究都在較小的時空尺度進行,且缺少長期定位研究。加強林窗對地下生態(tài)系統(tǒng)結(jié)構(gòu)、過程與功能的影響研究,重點闡明林窗對土壤碳氮分配、循環(huán)和固持機制,已成為林窗生態(tài)學(xué)今后的重要研究方向(管云云等,2016)。
華山松(Pinusamandii)人工林是神農(nóng)架林區(qū)的典型森林類型。自2012年以來,神農(nóng)架林區(qū)華山松大小蠹(Dendroctonusarmandi)呈暴發(fā)趨勢,對華山松林造成了嚴重危害,對林區(qū)生物多樣性、生態(tài)景觀和生態(tài)系統(tǒng)穩(wěn)定性造成了較大破壞,造成重大的經(jīng)濟和生態(tài)損失。根據(jù)洪承昊等(2017)對華山松大小蠹擴散規(guī)律調(diào)查研究發(fā)現(xiàn),在神農(nóng)架地區(qū),華山松大小蠹主要危害 30年以上的健康華山松,受侵害的林木最快可以在60 d內(nèi)枯死,受蟲害干擾后的林分在一段時間內(nèi)會出現(xiàn)大量分解程度不一的枯立木。本文以遭受大小蠹侵害的華山松人工林為研究對象,分析華山松人工林土壤呼吸變化特征,研究枯立木分布與分解程度,以及由枯立木形成的林窗對土壤呼吸的影響,可進一步了解森林土壤碳循環(huán)及其影響機制,有助于未來開展森林病蟲害生態(tài)控制治理決策的制定,并為精確估算土壤碳排放提供理論依據(jù)。
1 研究區(qū)概況
研究區(qū)域位于神農(nóng)架國家公園(110°03′—110°33′E,31°21′—31°36′N)。區(qū)內(nèi)最低海拔 398 m,最高海拔為3106.2 m,為華中第一峰。氣候?qū)儆诒眮啛釒蚺瘻貛н^渡氣候,年平均氣溫12.0 ℃,空氣相對濕度74%,降水量800—2500 mm,隨著海拔增高而增加,全年日照時數(shù)1858 h。區(qū)內(nèi)土壤類型沿海拔的升高分為 5大類:山地黃棕壤、山地棕壤、山地暗棕壤、棕色針葉林土和山地草甸土。植被由低海拔到高海拔依次分布有常綠闊葉林、常綠落葉闊葉混交林、針闊混交林、針葉林、灌叢和草甸。
華山松是神農(nóng)架地區(qū)分布的主要針葉林樹種,既分布有華山松純林,還分布著華山松與山楊(Populusdavidiana)、銳齒槲櫟(Quercusaliela)、糙皮樺(Betulaalbo-sinensis)等落葉闊葉樹種形成的針闊混交林。在垂直高度上,分布在海拔1200—2500 m,集中分布區(qū)位于海拔1500—2000 m。
2 主要研究方法
2.1 樣地設(shè)置與調(diào)查
本研究在湖北神農(nóng)架森林生態(tài)系統(tǒng)國家定位觀測研究站試驗地進行。在2019年3月,選擇遭受蟲害干擾的華山松人工林為研究對象,設(shè)置2個面積為50 m×50 m的樣地,同時將樣地劃分成10 m×10 m的樣方。所選林分于20世紀70年代中期種植,造林株行距為2 m×3 m,自2016年開始林內(nèi)出現(xiàn)大量受大小蠹侵害的林木,2017—2018年新感染蟲害木逐漸減少。為了減少樣地的空間異質(zhì)性,選擇土壤類型相同,海拔、坡向、林分年齡、樹種組成等立地條件相近的樣地。樣地基本信息及土壤理化性質(zhì)見表1。
2.2 土壤呼吸環(huán)設(shè)置與測定
在1號樣地中,在每個10 m×10 m的樣方頂點處機械布置土壤呼吸環(huán),共設(shè)置36個呼吸環(huán),呼吸環(huán)位置分布示意見圖1。在2號樣地中,分別在上、中、下坡各尋找1個面積50—100 m2的林窗,林窗主要因為大小蠹蟲害的干擾,以及其它自然因素影響而形成,在林窗中央設(shè)置3個呼吸環(huán),并在每個林窗附近的林下設(shè)置3個呼吸環(huán),共設(shè)置18個呼吸環(huán)。土壤呼吸測定環(huán)規(guī)格為直徑 20 cm、高度 8 cm,其中5 cm埋入地下。所有土壤呼吸環(huán)在2019年3月設(shè)置好,整個試驗過程中保持不動。
在2019年4—11月每月中旬選取天氣狀況較為一致的晴朗天氣測定1次土壤呼吸,每次測定時間為 09:00—16:00。土壤呼吸利用 Li-8100便攜式土壤呼吸測量系統(tǒng)測定。同時利用Li-8100配備的溫濕度探頭測定土壤 5 cm溫度以及體積含水量,每個土壤呼吸環(huán)每次測定2次取平均值。
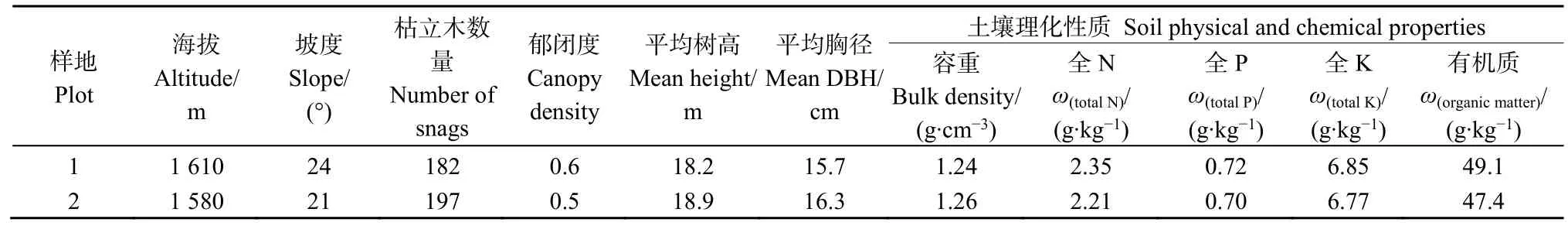
表1 樣地基本情況Table1 The setting of plots
2.3 枯立木分布及分解程度調(diào)查
2019年9月,在1號樣地中進行枯立木分布狀況及分解程度調(diào)查。根據(jù)樣地內(nèi)林木冠幅特征,以每個土壤呼吸環(huán)為中心,分別調(diào)查距離呼吸環(huán) 0—1、1—2、2—3、3—4 m內(nèi)枯立木及活立木數(shù)量。同時根據(jù)閆恩榮等(2005)提出的枯立木分解等級標準,并結(jié)合樣地華山松枯立木分解特征,分別統(tǒng)計輕度分解(細枝尚存,粗枝和樹皮較完整)、中度分解(無細枝,粗枝部分存在,樹皮松軟)、重度分解(無枝條,樹皮較少或全部脫落)枯立木的數(shù)量。
2.4 數(shù)據(jù)分析
以土壤呼吸速率作為因變量,土壤溫度和土壤含水量作為自變量,利用直線模型和指數(shù)模型分析土壤溫濕度與土壤呼吸的關(guān)系。各關(guān)系模型如下:
式中:RS表示土壤呼吸速率(μmol·m-2·s-1),t表示5 cm深度土壤溫度(℃),W表示5 cm深度土壤含水量,a、b、c表示待定參數(shù)。
土壤呼吸的溫度敏感性(Q10)計算公式為:
式中:b值為公式(2)中計算得出的常量b。
本研究以1號樣地數(shù)據(jù)分析土壤呼吸速率不同季節(jié)變化規(guī)律,土壤溫度、土壤含水量和枯立木分布對土壤呼吸的影響;以2號樣地數(shù)據(jù)比較林窗內(nèi)外土壤呼吸差異。數(shù)據(jù)處理和統(tǒng)計分析采用 Excel 2016和SPSS 19.0完成,采用ANOVA方差分析法檢驗土壤呼吸速率在不同月份間的差異顯著性,采用Pearson相關(guān)系數(shù)檢驗土壤呼吸速率與枯立木數(shù)量相關(guān)性。
3 結(jié)果與分析
3.1 土壤呼吸的月動態(tài)及季節(jié)差異
華山松人工林生長期土壤呼吸速率月變化規(guī)律具有單峰曲線特征(圖2)。從4—7月,隨著土壤溫度上升,土壤呼吸速率逐漸增加,7月達到最大值(3.80 μmol·m-2·s-1);從 8—11 月,隨著土壤溫度下降,土壤呼吸速率逐漸降低,11月達到最小值(1.15 μmol·m-2·s-1)。方差分析表明,不同月份之間土壤呼吸速率達到極顯著差異(P<0.01)。土壤呼吸在不同的季節(jié)同樣存在差異,在春季(3—5月)、夏季(6—8月)、秋季(9—11月)土壤呼吸速率分別為 1.78—2.79、2.81—3.80、1.15—2.60 μmol·m-2·s-1。可見,在夏季土壤呼吸明顯高于春季和秋季,而春季和秋季土壤呼吸速率差異不大。
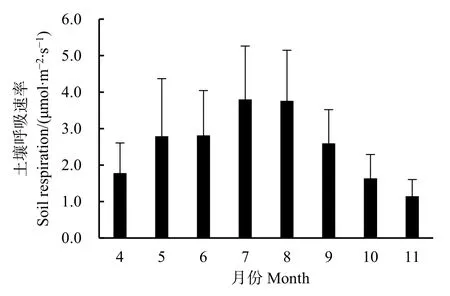
圖2 土壤呼吸速率月變化Fig.2 Month variations of soil repiration rate
3.2 土壤溫濕度對土壤呼吸的影響
3.2.1 溫度對土壤呼吸速率的影響
華山松人工林土壤呼吸速率與土壤溫度的關(guān)系如圖 3。從圖中可知,土壤呼吸與土壤溫度無論是線性相關(guān)或指數(shù)相關(guān)都達到了極顯著水平(P<0.01),線性回歸方程決定系數(shù)R2略高于指數(shù)回歸方程,說明線性模型能夠更好的模擬土壤呼吸速率與土壤溫度的關(guān)系。
3.2.2 土壤含水量對土壤呼吸的影響
觀測期華山松人工林土壤含水量變化范圍為30.4%—40.1%,不同月份間土壤含水量變幅不大,且沒有明顯變化規(guī)律。土壤呼吸速率與土壤含水量的關(guān)系如圖 4,從圖中可知,土壤呼吸速率與土壤含水量呈負相關(guān),但相關(guān)性沒有達到顯著水平(P>0.05)。
3.2.3 溫濕度對土壤呼吸交互作用
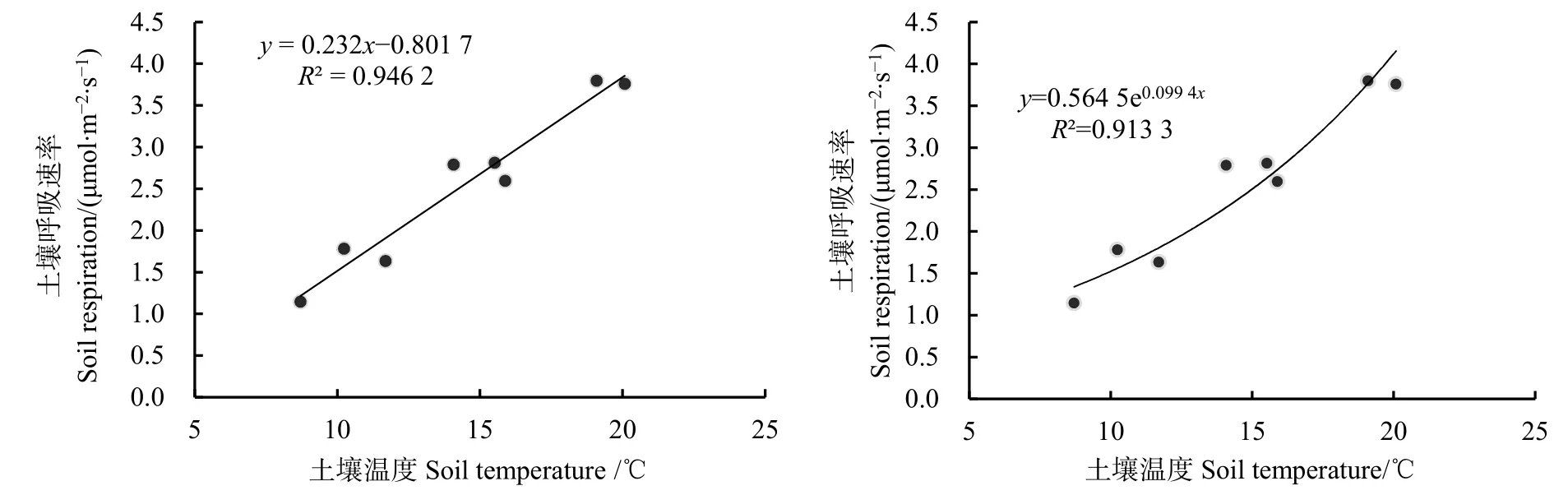
圖3 土壤呼吸與土壤溫度的關(guān)系Fig.3 Correlation between soil respiration and soil temperature
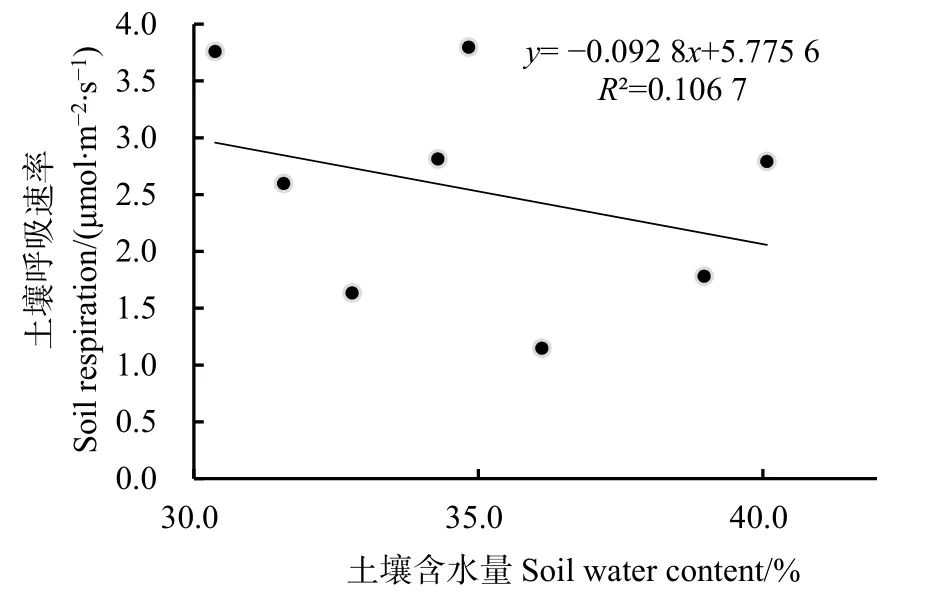
圖4 土壤呼吸與土壤含水量的關(guān)系Fig.4 Relationship between soil respiration and soil water content
采用線性的雙因素模型對土壤呼吸與土壤溫濕度關(guān)系進行回歸分析,回歸方程如下。從回歸方程可知土壤呼吸與土壤溫濕度相關(guān)性達到極顯著水平(P<0.01),且雙因素復(fù)合模型在一定程度上預(yù)測土壤呼吸的準確性更高。
Y=0.2660t+7.5913W-3.9380,
F=951.79**,R2=0.9974
3.3 枯立木分布對土壤呼吸的影響
3.3.1 枯立木數(shù)量與分布
不同土壤呼吸觀測點周圍枯立木數(shù)量呈隨機分布,調(diào)查統(tǒng)計結(jié)果見表2。距離36個觀測點4 m半徑范圍內(nèi)共計 148棵枯立木,其中輕度分解 18棵,中度分解59棵,高度分解71棵。
3.3.2 枯立木數(shù)量對土壤呼吸的影響
不同呼吸點測定的土壤呼吸速率與觀測點周邊不同距離范圍內(nèi)的枯立木及總立木數(shù)量相關(guān)性分析結(jié)果見表 3。從表中可知,土壤呼吸速率與距離觀測點不同距離枯立木數(shù)量均呈負相關(guān),在距離呼吸觀測點0—3 m的范圍內(nèi),隨著距離的增加,土壤呼吸與枯立木數(shù)量相關(guān)性逐漸增強,并與 0—3 m內(nèi)總枯立木數(shù)量相關(guān)性達到顯著水平(P<0.05)。土壤呼吸速率與不同范圍內(nèi)的立木總數(shù)均呈負相關(guān),但相關(guān)性不顯著(P>0.05)。
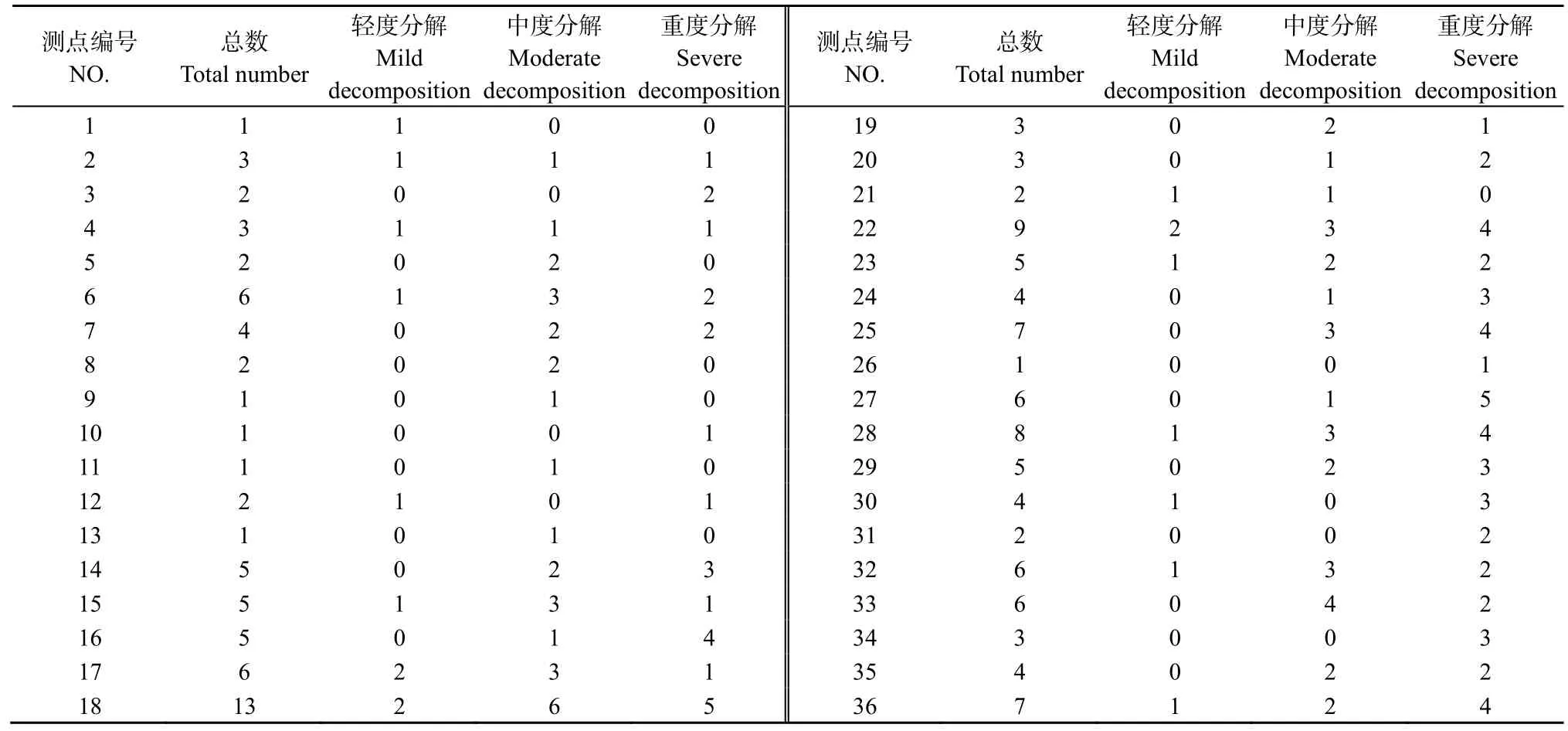
表2 不同土壤呼吸觀測點4 m范圍內(nèi)枯立木數(shù)量Table 2 Number of snags 4 meters from the each soil respiration observation point
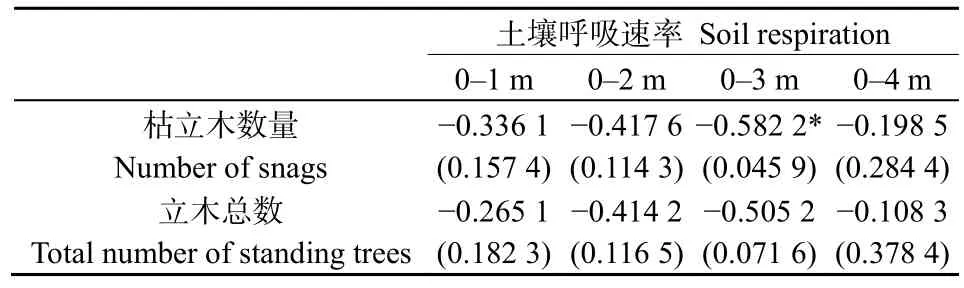
表3 土壤呼吸與立木數(shù)量相關(guān)性Table 3 The correlation between soil respiration and the quantity of standing trees
3.3.3 枯立木分解程度對土壤呼吸的影響
由于 0—3 m的范圍內(nèi)枯立木數(shù)量與土壤呼吸相關(guān)性達到顯著水平,因此在此基礎(chǔ)上進一步分析0—3 m的范圍內(nèi)枯立木分解程度與土壤呼吸速率的關(guān)系,對輕度分解數(shù)量(XL)、中度分解數(shù)量(XM)、高度分解數(shù)量(XH)與土壤呼吸速率進行多元線性回歸分析,回歸方程如下。經(jīng)檢驗回歸方程達到顯著水平,進一步對方程各自變量回歸系數(shù)進行顯著性檢驗,可知不同分解程度枯立木數(shù)量對土壤呼吸速率的影響大小依次為高度分解數(shù)量 (XH)>中度分解數(shù)量 (XM)>輕度分解數(shù)量 (XL),說明高度分解的枯立木數(shù)量對土壤呼吸速率存在較大影響。
Y= -0.0711XL-0.1333XM-0.1567XH+2.8120,F=19.2*,R2=0.5821
3.3.4 林窗內(nèi)外土壤呼吸速率差異
林窗內(nèi)與林下土壤溫度與土壤呼吸速率月變化情況見圖 5。觀測期林窗內(nèi)和林下土壤溫度平均值分別為15.2 ℃和14.0 ℃,土壤呼吸速率平均值分別為 3.43 μmol·m-2·s-1和 2.19 μmol·m-2·s-1,方差分析表明林窗內(nèi)外土壤呼吸速率差異達到極顯著水平(P<0.01),且在各個月份,林窗內(nèi)土壤呼吸速率均顯著高于林下。
根據(jù)土壤呼吸溫度敏感性系數(shù)(Q10)計算公式,可得出林窗內(nèi)和林下Q10值分別為2.94和2.46,說明隨著枯立木的大量出現(xiàn)而形成林窗,林地土壤呼吸溫度敏感性系數(shù)增加,土壤呼吸受土壤溫度的變化影響更明顯。
4 討論
4.1 土壤溫濕度與土壤呼吸
在影響土壤呼吸的眾多因子中,土壤溫度與土壤含水量兩者往往是最主要的限制因子,其主要通過影響土壤微生物及植物根系的生理活動進一步影響土壤呼吸過程(Raich et al.,1995)。本研究中華山松林土壤呼吸與土壤 5 cm處溫度的相關(guān)性分析表明,土壤呼吸與土壤溫度具有顯著的線性相關(guān)關(guān)系與指數(shù)相關(guān)關(guān)系,這與其他學(xué)者的研究結(jié)果基本一致(Li et al.,2019;胡樂寧等,2012)。華山松林土壤呼吸與土壤含水量的相關(guān)性不顯著,說明在本研究區(qū)域土壤含水量并不是土壤呼吸的限制因子,這與該區(qū)域所處氣候環(huán)境有關(guān)。神農(nóng)架國家公園位于北亞熱帶濕潤季風(fēng)區(qū),不同于干旱或半干旱地區(qū),土壤全年較為濕潤,且變化幅度不大。當土壤環(huán)境較濕潤,同時土壤水分并不是環(huán)境限制因子時,土壤溫度通常是影響土壤呼吸的最主要影響因素(Schimel,1995;Wu et al.,2014;劉益君等,2016)。本研究結(jié)果中土壤呼吸與土壤含水量呈負相關(guān),表明土壤含水量對土壤呼吸的具有一定抑制作用,其原因可能是土壤含水量過高導(dǎo)致土壤顆粒粘結(jié)性增強,使得通透性變差,減少了土壤孔隙中氧氣的有效性,同時降低土壤根系活動及微生物的活性,從而對土壤呼吸產(chǎn)生負作用(陳小平等,2018)。土壤溫度和土壤含水量的雙因素模型優(yōu)于只考慮土壤溫度或含水量的單因素模型,與以往研究結(jié)果相同(黃石德,2009;陳燦等,2017),說明土壤溫度與土壤含水量的雙因素交互作用可更好地解釋土壤呼吸的變異。
4.2 枯立木數(shù)量、分解程度與土壤呼吸
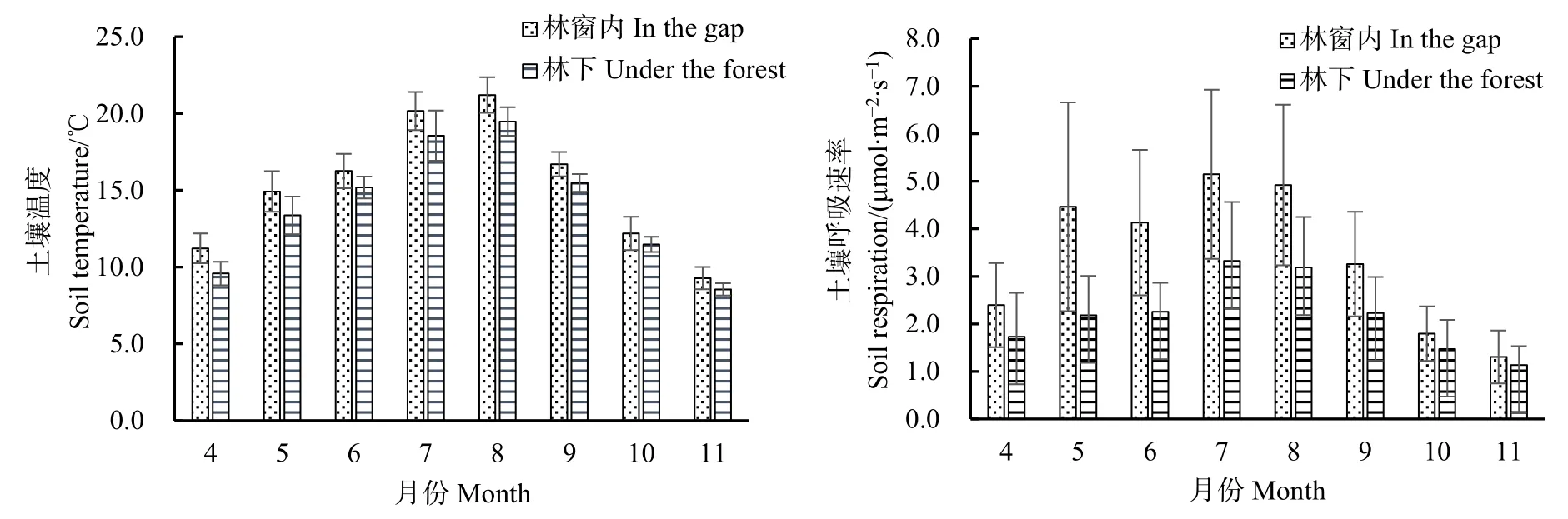
圖5 林窗內(nèi)外土壤溫度與土壤呼吸速率比較Fig.5 Comparation of soil temperature and respiration rate in/out of the gap
枯立木對土壤呼吸產(chǎn)生影響包括多方面原因:一是枯立木的出現(xiàn)改變了林冠層結(jié)構(gòu),以往研究表明冠幅及冠層結(jié)構(gòu)對土壤呼吸的影響非常顯著(Stoyan et al.,2000;王蕊等,2014),且林冠層結(jié)構(gòu)的變化將進一步改變林地土壤生境(管云云等,2016);二是直接減少了根系的自養(yǎng)呼吸;三是枯立木死亡根系分解過程中不斷釋放C、N、P等元素和化學(xué)物質(zhì),造成土壤元素在空間上的異質(zhì)性,并影響了微生物的C循環(huán)(Wiebe et al.,2014)。此外還有研究指出,枯立木的分解雖然增加了土壤中的C和N,但枯立木上流失的物質(zhì)也抑制了下方土壤的C和N循環(huán)(Bantle et al.,2014)。本研究發(fā)現(xiàn)華山松林內(nèi)枯立木數(shù)量對土壤呼吸具有負效應(yīng),并與距離土壤呼吸環(huán)3 m半徑內(nèi)枯立木數(shù)量相關(guān)性達到顯著水平(P<0.05)。通過對試驗樣地調(diào)查發(fā)現(xiàn)華山松人工林冠幅主要為5—6 m,說明華山松人工林林冠層結(jié)構(gòu)破壞是影響土壤呼吸的重要因素之一。基于距離土壤呼吸測定點3 m范圍枯立木的分解程度的多元線性回歸分析得出重度分解的枯立木對土壤呼吸的負效應(yīng)最為明顯,表明隨著枯立木分解程度增加,對土壤呼吸的影響程度逐漸加強。這可能是因為隨著枯立木的形成與逐漸分解,林木自身根系呼吸逐漸減弱,根系周圍土壤微生物數(shù)量減少,且減少了從土壤中吸收營養(yǎng)物質(zhì),代謝活動降低,同時也不利于土壤結(jié)構(gòu)的改善,導(dǎo)致土壤呼吸受到影響。關(guān)于枯立木數(shù)量分布與分解程度對土壤呼吸的影響機制,有待后續(xù)深入研究。
4.3 林窗與土壤呼吸
本研究發(fā)現(xiàn),觀測期林窗內(nèi)土壤呼吸速率均顯著高于林下,提高了約1.5倍,林窗的出現(xiàn)顯著提高了土壤呼吸速率,這與楊開軍等(2016)研究結(jié)果一致。林窗的形成導(dǎo)致林窗內(nèi)土壤溫度的增加以及濕度的降低,土壤有機質(zhì)分解速率較快(張遠彬等,2003);林窗為土壤微生物提供了更好的繁殖條件,促進生態(tài)系統(tǒng)的養(yǎng)分循環(huán)過程(費菲等,2018);林窗對凋落物的分解有顯著影響,且分解速率隨著林窗的大小而發(fā)生變化,凋落物的質(zhì)量損失速率隨林窗面積增加而增加(吳慶貴等,2016),這些因素都可能導(dǎo)致林窗內(nèi)土壤呼吸速率的增加。另外,張良輝(2010)研究發(fā)現(xiàn),針葉人工林初植密度過大導(dǎo)致林下草本和灌木植物較少,而林窗內(nèi)擁有更多的灌草植物和土壤動物,這對林窗土壤呼吸的增加也有一定貢獻。而Saner et al.(2009)對馬來西亞熱帶雨林的研究表明,林下土壤呼吸速率達林窗內(nèi)的2倍,與本研究結(jié)果相反。這可能是由于研究的森林類型及其所處的氣候條件差異很大造成的,而且林窗內(nèi)具體環(huán)境狀況比較復(fù)雜,取決于林窗大小、形成時間、物種組成等多種因素的共同作用。
土壤呼吸溫度敏感性系數(shù)(Q10)受土壤生物、底物、環(huán)境因子三大影響因素的影響(楊慶朋等,2011)。本研究中,華山松林窗內(nèi)土壤溫度升高,土壤呼吸溫度敏感性系數(shù)高于林下,說明隨著枯立木的大量出現(xiàn)形成林窗,土壤呼吸受土壤溫度變化的影響更明顯,這與黃石德(2009)和陳燦等(2017)的研究結(jié)果一致,說明了神農(nóng)架華山松林冠層對保持土壤呼吸的穩(wěn)定性具有重要作用。而楊慶朋等(2011)對以往多數(shù)研究總結(jié)發(fā)現(xiàn)土壤呼吸的溫度敏感性隨著溫度的升高而降低,Chen et al.(2005)通過對不同氣候帶 38個地點的土壤呼吸數(shù)據(jù)進行整合分析,結(jié)果表明不同氣候帶的土壤呼吸溫度敏感性系數(shù)均隨著土壤溫度的升高而降低,這與本研究結(jié)果相反,造成研究結(jié)果的差異可能是多數(shù)森林土壤呼吸溫度敏感性的研究僅局限在林下,而忽略了林窗這個復(fù)雜環(huán)境對土壤呼吸的影響。
5 結(jié)論
(1)神農(nóng)架華山松人工林土壤呼吸速率呈現(xiàn)明顯的單峰型季節(jié)動態(tài)變化,峰值出現(xiàn)在7月;土壤呼吸速率與土壤溫度具有極顯著的線性關(guān)系和指數(shù)關(guān)系(P<0.01),與土壤含水量相關(guān)性不顯著(P>0.05),土壤溫濕度雙因素復(fù)合模型可以更準確預(yù)測土壤呼吸速率。
(2)土壤呼吸速率與枯立木數(shù)量呈負相關(guān),且受土壤呼吸環(huán)3 m半徑范圍內(nèi)的枯立木數(shù)量的顯著影響(P<0.05);0—3 m范圍內(nèi)枯立木分解程度越高,土壤呼吸速率受影響程度越大,重度分解的枯立木對土壤呼吸速率影響最大。
(3)隨著枯立木的出現(xiàn)逐漸形成林窗,導(dǎo)致林窗內(nèi)外土壤呼吸呈極顯著差異(P<0.01),林窗的出現(xiàn)提高了林窗內(nèi)土壤呼吸速率;林窗內(nèi)土壤呼吸溫度敏感性系數(shù)(Q10)高于林下,林窗的形成提升了土壤呼吸溫度敏感性。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學(xué)生數(shù)理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
天津外國語大學(xué)學(xué)報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當代陜西(2021年2期)2021-03-29 07:41:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設(shè)計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學(xué)報(2017年2期)2017-07-05 08:13:02
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
