圓斑星鰈(Verasper variegatus)wt1基因的克隆與表達分析*
2020-11-24 01:18:42楊珍珍陳四清張盛農(nóng)劉長琳葛建龍
海洋與湖沼 2020年6期
楊珍珍 邊 力 張 巖 陳四清 常 青 張盛農(nóng) 劉長琳 葛建龍
圓斑星鰈(Verasper variegatus)wt1基因的克隆與表達分析*
楊珍珍1, 2邊 力1, 3張 巖1陳四清1, 3①常 青1張盛農(nóng)1劉長琳1葛建龍1
(1. 中國水產(chǎn)科學研究院黃海水產(chǎn)研究所 青島 266071;2. 上海海洋大學水產(chǎn)與生命學院 上海 201306;3. 青島海洋科學與技術試點國家實驗室 海洋漁業(yè)科學與食物產(chǎn)出過程功能實驗室 青島 266071)
圓斑星鰈()雌魚生長快, 成熟雌魚個體大小是雄魚的2倍以上, 開展性別相關基因的功能研究, 對于探究圓斑星鰈性別決定機制, 建立單性培育技術具有重要意義。本研究獲得了及兩個同源基因,基因全長為3263bp, 預測開放閱讀框(ORF)長為1245bp, 編碼415個氨基酸, 5′-UTR和3′-UTR分別長372bp和1640bp;基因全長為2312bp, 預測開放閱讀框(ORF)長為1281bp, 編碼427個氨基酸, 5’-UTR和3’-UTR分別長369bp和659bp。基因編碼氨基酸分子量為46.2kDa, 理論等電點為9.24, 無跨膜結構及信號肽, 在ORF末端有4個鋅指結構, 編碼KTS三肽;基因編碼氨基酸分子量為46.95kDa, 理論等電點為8.99, 無跨膜結構及信號肽, 在ORF末端有4個鋅指結構, 并且編碼 KTS三肽。基因表達結果表明:和基因主要在圓斑星鰈性腺中表達, 精巢的表達高于卵巢, 腎臟的表達量顯著高于其他組織, 推測基因和基因在性腺和腎臟發(fā)育過程及功能方面均發(fā)揮重要作用;在早期發(fā)育階段,基因在原腸期之前微弱表達, 從原腸早期開始逐漸上升至神經(jīng)胚期表達量達到最高, 之后逐漸下降, 直至孵化階段, 推測基因在圓斑星鰈原始生殖細胞分化過程及性腺發(fā)育中發(fā)揮重要作用。
圓斑星鰈;;基因克隆;表達分析
腎母細胞瘤系嬰幼兒最常見的惡性腫瘤之一, 多發(fā)生于3歲以內兒童(Pelletier, 1991), 發(fā)病率約為1/10000(Sharma, 1992)。1899年Max Wilms首次對腎母細胞瘤進行了詳細描述(Kreidberg, 1993), 因此, 又將其命名為Wilms瘤。1990年Gessler等分離克隆出一個與Wilms瘤相關的基因(Gessler, 1990), 命名為Wilms’tumor 1()。基因可編碼含有鋅指結構的蛋白質, 在性腺發(fā)育及分化過程中識別、結合和調控雄性性別決定基因(Hossain, 2001)類固醇生成因子-1(sf-1) (Wilhelm, 2002)及苗勒管抑制物(Hossain, 2003)等性別發(fā)育關鍵基因。人類基因突變會導致性腺和腎臟等器官發(fā)育異常及相關功能障礙(Klamt, 1998), 導致Denys-Drash綜合征、Frasier綜合征及小兒腎癌等疾病。敲除小鼠基因后, 其性腺及腎臟等器官均出現(xiàn)發(fā)育缺陷, 甚至導致腎功能衰竭(Wiener, 1996; Moore,1999)。由于魚類特有的基因組復制,基因分化為了和兩個同源基因, 目前已在斑馬魚()(Bollig, 2006)、青鳉()(Klüver,2009)、半滑舌鰨()(張紅等, 2014)、牙鲆()(孫近近, 2016)、黃鱔()(胡青等, 2014)及尼羅羅非魚()(江東能, 2016)等開展了基因的克隆及功能研究。
圓斑星鰈俗稱花斑寶、花片等, 屬脊索動物門(Chordata)、硬骨魚綱(Osteichthyes)、鰈形目(Pleuronectiformes)、鰈科(Pleuronectidae)、星鰈屬()。其外觀漂亮, 體型較大, 食性范圍廣, 生長速度快, 出肉率高, 抗病能力強, 肉質細膩鮮美, 鰭邊堅韌有彈性, 富含多種氨基酸、維生素及微量元素, 屬于名貴的海水品種, 目前已成為我國北方優(yōu)良的養(yǎng)殖對象。在生長過程中, 雄性個體3年可性成熟, 雌性個體需要4年才可達到性成熟, 而且雌性個體的生長明顯快于雄性個體。目前, 在圓斑星鰈中僅對基因(柳學周等, 2013)、基因(張樂樂, 2018)、基因(張樂樂等, 2018)及基因(楊珍珍等, 2020)做了相關研究, 其性別決定和生殖調控的分子機制仍不清楚, 因此我們利用RACE末端擴增技術克隆了圓斑星鰈和基因cDNA全長并分析了其序列結構特征, 利用實時熒光定量PCR技術對其不同發(fā)育時期的胚胎、仔魚以及雌雄成魚的不同組織進行表達分析, 以闡明和基因在圓斑星鰈性腺分化及發(fā)育中的作用, 為研究圓斑星鰈性別決定及單性繁育等提供新的靶標基因。
1 材料與方法
1.1 樣品制備及wt1基因全長序列克隆
從山東省煙臺天源水產(chǎn)有限公司采集圓斑星鰈不同發(fā)育時期的胚胎, 包括單細胞期(卵裂前胚盤隆起時期)(0h)、2細胞期(4h)、8細胞期(10h)、16細胞期(12h)、32細胞期(13h)、桑椹期(18h)、高囊胚期(20h)、低囊胚期(26h)、原腸早期(27h)、原腸中期(29h)、原腸晚期(77h)、神經(jīng)胚期(98h)、晶體形成期(136h)、心跳出現(xiàn)期(163h)、孵化前期(188h)、脫膜孵化期(197h), 不同時期的仔稚魚(孵化后5d, 10d, 20d, 30d, 45d, 58d, 68d, 78d, 92d)及雌雄成魚[體長(24.0±0.5)cm, 體重(330±20)g]的不同組織(腦、眼、鰓、心、肝、腸、脾、性腺、腎及肌肉), 將采集的樣品用液氮充分研磨, 根據(jù)Trizol(TaKaRa)實際說明書提取各組織的總RNA, 經(jīng)Thermo紫外分光光度計(NanoDrop 2000)和1.2%的瓊脂糖凝膠電泳檢測, 將高質量的RNA用SMARTTMRACE cDNA Amplification RACE (TaKaRa)試劑盒分別合成3’RACE和5’RACE cDNA。之后根據(jù)本實驗室構建的轉錄組數(shù)據(jù)庫獲得和基因的EST序列, 利用Primer Primer 5.0軟件設計和基因的3′RACE和5′RACE特異性引物(表1), 以圓斑星鰈成魚性腺組織cDNA為模板分別進行擴增, PCR反應體系(20μL)為: Premix TaqTM(LA TaqTMVersion 2.0) 10μL、3’或5’特異性引物0.8μL、UPM(或NUP) 0.8μL、RACE-cDNA 1μL及ddH2O 6.4μL。PCR反應條件為: 94°C 5min;94°C 30s, 3’或5’特異性引物溫度30s, 72°C 1min, 35個循環(huán);72°C 10min。擴增產(chǎn)物經(jīng)1.2%瓊脂糖凝膠電泳檢測, 并在紫外線下切膠, 并用SanPrep柱氏DNA凝膠回收試劑盒回收, 利用PMD18-T Vector試劑盒(TaKaRa)進行連接(PMD18-T Vector 1μL, Solution I 5μL及純化產(chǎn)物4μL), 16°C反應3h。之后將連接液加入融化好的DH5α (Code No 9057, TaKaRa)中培養(yǎng), 挑取陽性單克隆, 經(jīng)菌落PCR鑒定后, 篩選目的菌液送至華大基因進行測序。
1.2 wt1基因序列分析
使用ORF Finder (http://www.ncbi.nlm.nih.gov/ projects/gorf/orfig.cgi), ExPASy (https://web.expasy. org/compute_pi/), SMART (http://smart.emblheidelberg. de/), Signal4.1 (http://www.cbs.dtu.dk/services/SignalP/)和NCBI (http://www.ncbi.nlm.nih.gov/Structure/cdd/ wrpsb.cgi)在線生物學軟件分析基因的開放閱讀框、分子量、理論等電點、信號肽及跨膜結構域。并用DNAMAN (Zemann,2006)和MEGA5.2 (Tamura,2011)軟件, 進行多重序列比對及構建系統(tǒng)進化樹。
1.3 wt1基因的表達特征分析
根據(jù)已獲得的基因全長序列, 設計熒光定量特異性引物,-actin作為內參基因(表1)。使用Applied BiosystemsTM7500 Real Time PCR instrument定量儀檢測, 反應程序: 95°C 10min;95°C 30s, 95°C 5s, 60°C 34s, 40個循環(huán);95°C 15s;60°C 1min;95°C 15s。采用2–DDCT的計算方法分析相對表達量, 通過SPSS 20.0軟件進行單因素分析, 利用OriginPro 2017作圖,<0.05代表具有顯著性差異。
2 結果
2.1 圓斑星鰈wt1基因全長和序列分析
通過擴增獲得基因全長為3263bp, 經(jīng)預測開放閱讀框(ORF)長為1245bp, 編碼415個氨基酸, 5’-UTR和3’-UTR分別長372bp和1640bp。經(jīng)過生物學分析, 推斷的415個氨基酸分子量為46.2kDa, 理論等電點為9.24, 無跨膜結構及信號肽, ORF末端有4個鋅指結構, 編碼KTS三肽(圖1)。基因全長為2312bp, ORF長1281bp, 編碼427個氨基酸, 5’-UTR和3’-UTR分別長369bp和659bp。的415個氨基酸分子量為46.95kDa, 理論等電點為8.99, 無跨膜結構及信號肽, ORF末端有4個鋅指結構, 編碼KTS三肽(圖2)。
表1 本研究中所使用的引物
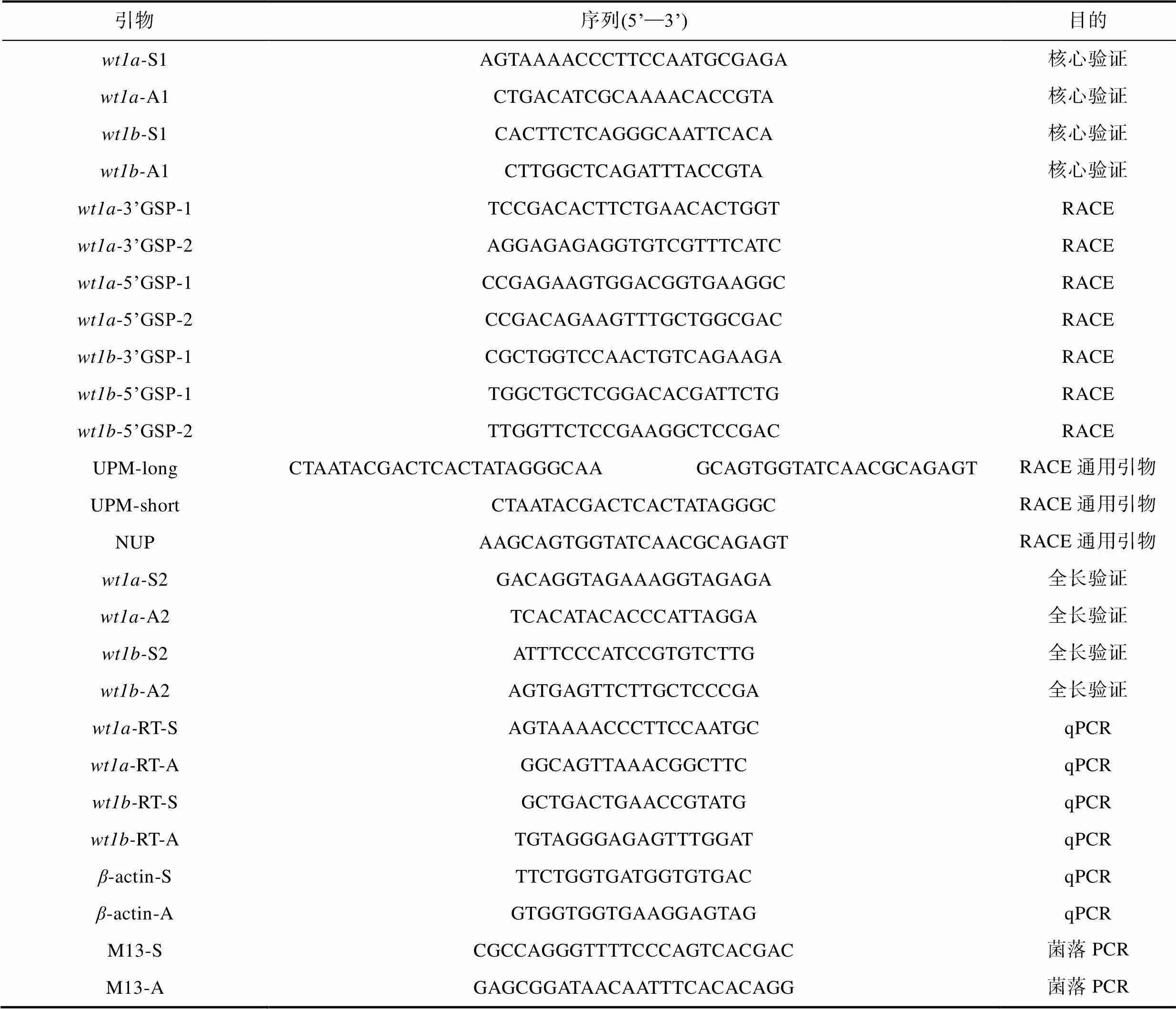
Tab.1 Nucleotide sequences of the PCR primers used in this study
2.2 wt1基因同源性和系統(tǒng)發(fā)育分析


圖1 wt1a基因cDNA序列全長及其編碼的氨基酸序列
注: 起始密碼子(ATG)與終止密碼子(TGA)用單線邊框標出, KTS是選擇性剪接, 鋅指結構區(qū)用單下劃線標出, polyA結尾用雙下劃線標出

圖2 wt1b基因cDNA序列全長及其編碼的氨基酸序列
注: 起始密碼子ATG與終止密碼子TGA用單線邊框標出, KTS是選擇性剪接, 鋅指結構區(qū)用單下劃線標出, polyA結尾用雙下劃線標出
2.3 Wt1基因在不同發(fā)育時期及不同組織中的表達分析
胚胎發(fā)育期研究結果顯示(圖6),基因在原腸期之前微弱表達, 從原腸早期開始逐漸上升至神經(jīng)胚期表達量達到最高, 之后開始呈下降趨勢, 直至孵化階段;基因從原腸晚期開始表達量逐漸升高, 上升至神經(jīng)胚期后開始下降, 并逐漸穩(wěn)定。研究基因和基因在仔稚魚不同發(fā)育時期中的表達量, 結果顯示(圖7)基因在孵后10d時的表達量顯著高于本實驗中的其他時期,基因在孵后5d時的表達量顯著高于本實驗中的其他時期, 之后逐漸下降。

圖3 wt1a基因編碼氨基酸與其他物種wt1a基因編碼氨基酸的多序列比對
注: 黑色邊框為鋅指結構區(qū), 紅色邊框為KTS。各物種WT1a蛋白序列登錄號: 尼羅羅非魚(XP_013121197.1)、點帶石斑魚(AFV66805.1)、大黃魚(XP_027128155.1)、黃尾鰤(XP_023262779.1)、黃鱔(XP_020460125.1)、斑馬魚(NP_571121.1)和青鳉(NP_001098171.1)

圖4 wt1b基因編碼氨基酸與其他物種wt1b基因編碼氨基酸的多序列比對
注: 黑色邊框為鋅指結構區(qū), 紅色邊框為KTS。各物種WT1b蛋白序列登錄號: 牙鲆(XP_019954195.1)、大菱鲆(AWP01692.1)、尼羅羅非魚(ARU81315.1)、黃鱔(XP_020471329.1)、大黃魚(XP_010740189.1)、點帶石斑魚(AFV66806.1)、攀鱸(XP_026234955.1)、紅鰭東方鲀(XP_011617397.1)和青鳉(NP_001098390.1)
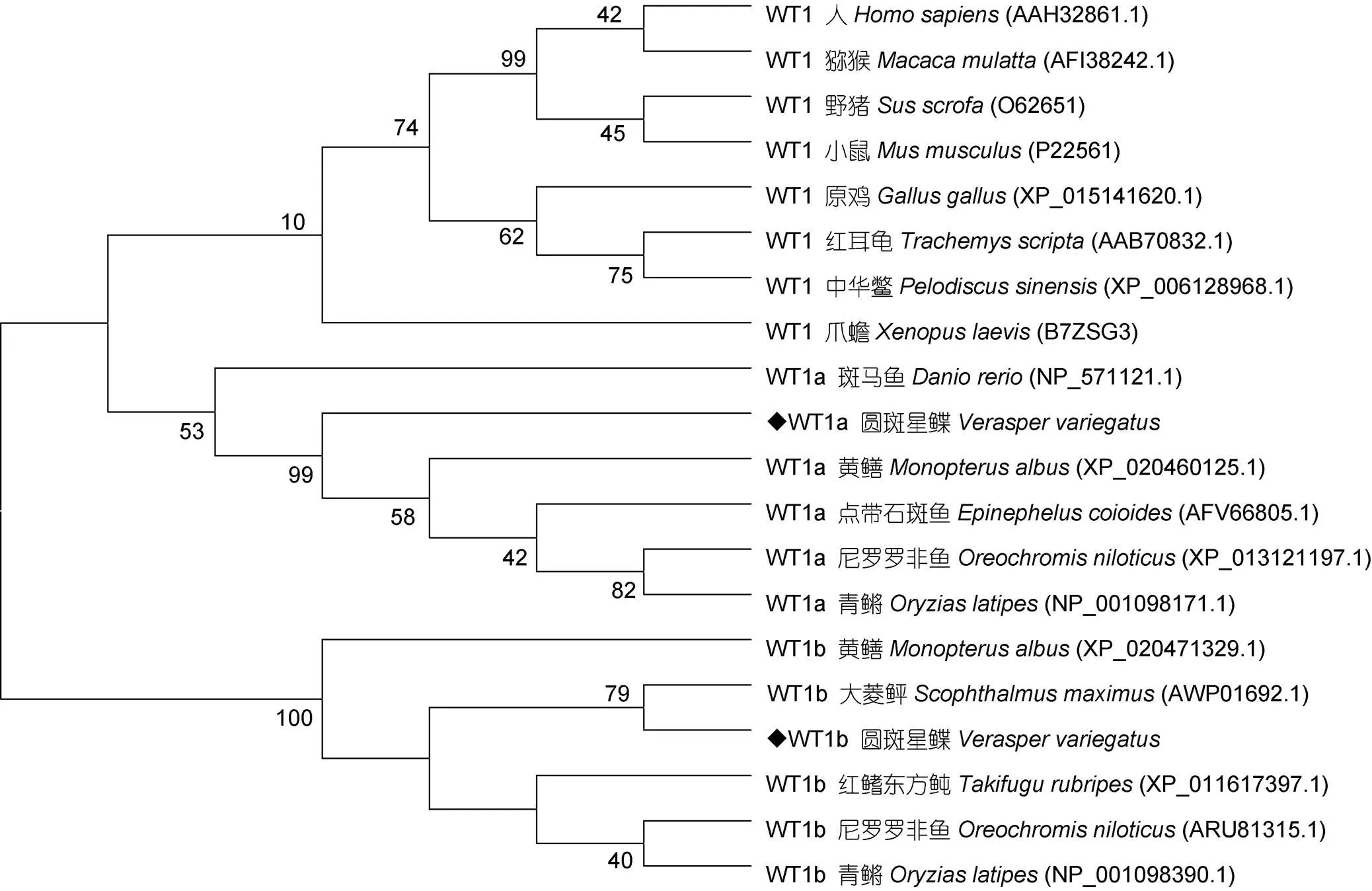
圖5 wt1基因編碼氨基酸序列的系統(tǒng)進化樹
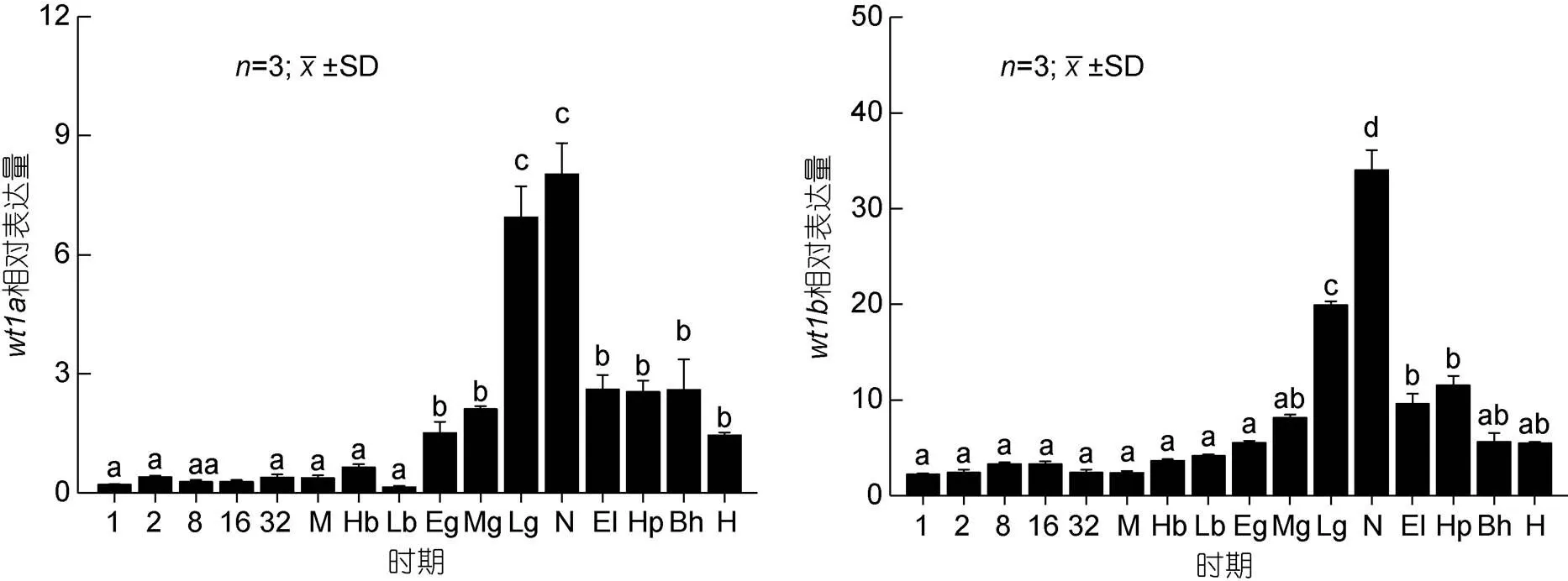
圖6 wt1 基因在胚胎發(fā)育不同階段中的表達
注: 1: 單細胞期; 2: 2細胞期; 8: 8細胞期; 16: 16細胞期; 32: 32細胞期; M: 桑椹胚期; Hb: 高囊胚期; Lb: 低囊胚期; Eg: 原腸早期; Mg: 原腸中期; Lg: 原腸晚期; N: 神經(jīng)胚期; El: 晶體形成期; Hp: 心跳出現(xiàn)期; Bh: 孵化前期; H: 脫膜孵化。不同字母間表示差異顯著(<0.05), 下同
組織表達結果顯示(圖8)基因和基因在雌雄成魚的各個組織中均有表達。基因和基因在雄性成魚精巢組織的表達量最高, 其次是雌性成魚的卵巢組織, 腎組織中也有較高表達, 3個組織的表達量均顯著高于其他組織。
3 討論
本研究用RACE方法獲得了圓斑星鰈和基因全長, 分別命名為基因和基因。兩個同源基因在ORF末端均具有連續(xù)四個(Cys)2(His)2型鋅指結構, 編碼KTS三肽。同源性分析發(fā)現(xiàn)基因和基因編碼氨基酸在羧基端鋅指結構區(qū)域均具有很高的相似性。基因系統(tǒng)分析結果顯示, 哺乳類、鳥類及爬行類聚為一大支, 魚類的WT1a及WT1b分別為一大支, 其中圓斑星鰈WT1b與大菱鲆WT1b聚為一支, 關系較近。此外, 相較于WT1b, 魚類WT1a與哺乳類等的WT1關系較近, 該結果與半滑舌鰨(張紅等, 2014)相似。研究發(fā)現(xiàn)基因可通過鋅指結構與DNA、RNA 結合, 從而正常行使其功能(Morrison,2008)。此外由于基因具有多個翻譯起始位點及不同的選擇性剪接方式, 可編碼多種蛋白亞型(Dong, 2015), 其中較為常見的+KTS亞型蛋白主要在RNA加工過程中起重要作用, 并在小鼠的性別決定中起重要作用(Hammes, 2001)。綜上, 我們推測基因和基因在圓斑星鰈性腺發(fā)育中具有重要作用。
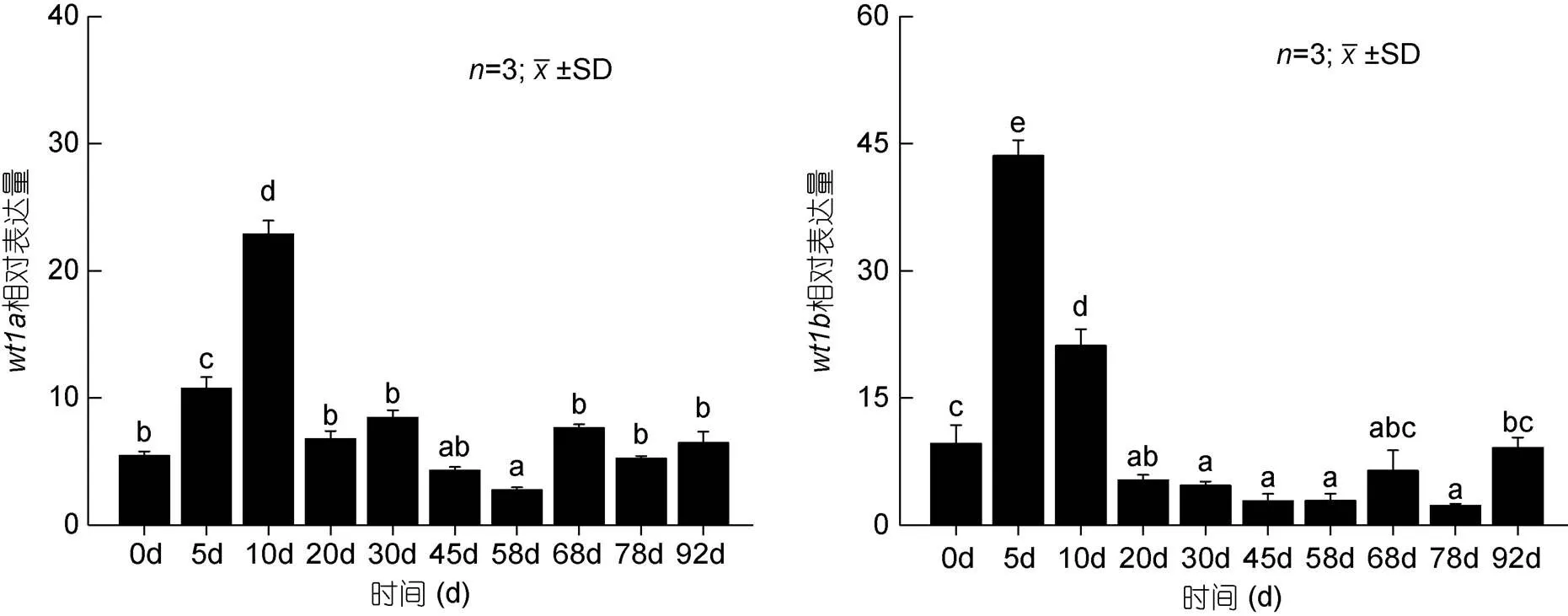
圖7 wt1基因在不同發(fā)育時期仔魚中的表達
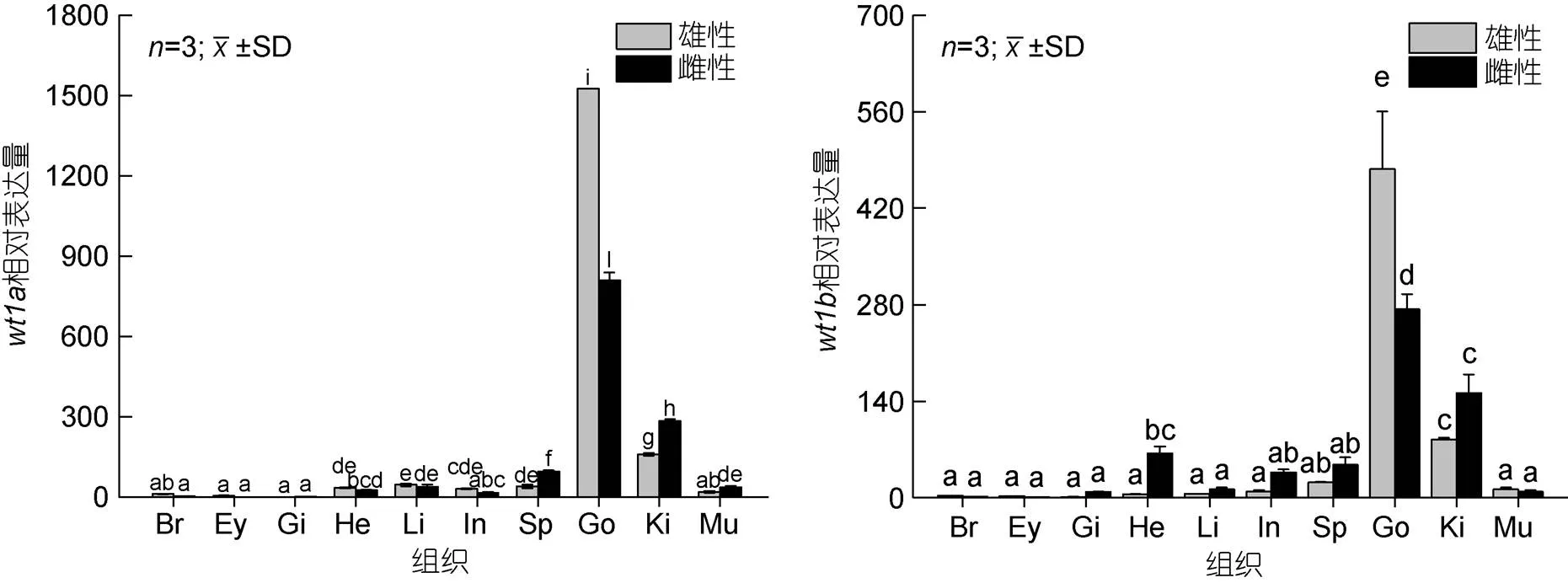
圖8 wt1基因在不同組織中的表達
注: Br: 腦; Ey: 眼; Gi: 鰓; He: 心臟; Li: 肝臟; In: 腸; Sp: 脾; Go: 性腺; Ki: 腎; Mu: 肌肉
在哺乳動物中,基因的突變會導致其性腺及腎臟等器官發(fā)育異常。如小鼠中缺失基因后, 其腎單位將無法形成, 導致間充質細胞無法正常增殖(Davies, 2004)。性腺分化過程中, 僅在小鼠精巢的支持細胞及卵巢的顆粒細胞中表達的基因可以促進生殖細胞的成熟, 小鼠基因缺失后, 精原細胞的分裂過程將被中斷(Zheng, 2013),
基因在原腸期前表達低, 從原腸早期開始逐漸上升至神經(jīng)胚期表達量達到最高, 之后逐漸下降, 該結果與斑馬魚(Bollig, 2006)、半滑舌鰨(張紅等, 2014)及牙鲆(孫近近, 2016)的表達模式相似。研究發(fā)現(xiàn), 神經(jīng)胚期是原始生殖細胞分化的關鍵時期(康現(xiàn)江等, 2010), 因此推測基因在圓斑星鰈原始生殖細胞分化過程中發(fā)揮作用。基因從原腸晚期開始表達量逐漸升高, 至神經(jīng)胚期后下降, 并逐漸穩(wěn)定, Bollig等(2006)研究發(fā)現(xiàn)在斑馬魚胚胎時期檢測到基因在12h(原腸期之后)時的表達量顯著增加, 同時原位雜交結果顯示基因在間介中胚層(原腸胚末期)表達, 而間介中胚層將分化為泌尿生殖系統(tǒng)的主要器官, 因此推測基因在性腺分化形成中發(fā)揮作用。作為同源性基因,和基因在圓斑星鰈胚胎發(fā)育過程中的神經(jīng)胚期表達量均達到最高, 表明和基因均在圓斑星鰈原始生殖細胞分化過程中發(fā)揮作用,基因在圓斑星鰈胚胎發(fā)育中表達量顯著上升時間較早, 因此推測基因在圓斑星鰈胚胎發(fā)育中較早地發(fā)揮作用, 仍需進一步驗證。
基因在孵后10d時的表達量顯著高于其他時期, 這一表達模式與牙鲆相似, 在牙鲆中基因對其生殖脊細胞的存活起著重要作用(孫近近, 2016), 因而基因可能在圓斑星鰈性腺的發(fā)育過程中起作用;基因在孵后5d時的表達量顯著高于其他時期, 之后逐漸下降, 其表達量顯著升高的時間早于基因, 推測在仔稚魚發(fā)育過程中基因更早地發(fā)揮作用, 但因相關研究較少, 其在圓斑星鰈性腺發(fā)育中的作用仍需進一步研究。
基因和基因在精巢與卵巢中的表達量均顯著高于其他組織, 且精巢的表達量顯著高于卵巢, 其次在腎臟組織中的表達量顯著高于其他組織。在斑馬魚(Bollig, 2006)和青鳉(Klüver,2009)中,基因在性腺及腎臟中均具有較高表達, 且在青鳉中發(fā)現(xiàn)基因的突變會導致原始生殖細胞數(shù)量減少。在半滑舌鰨(張紅等, 2014)中,基因在性腺及腎臟中也有較高表達, 且該研究顯示基因在性腺分化及發(fā)育過程中持續(xù)表達。在牙鲆(孫近近, 2016)中,基因主要在性腺中表達, 同樣在精巢中的表達量高于卵巢, 表明基因在性腺發(fā)育及功能維持方面起重要作用。在黃鱔(胡青等, 2014)中,基因在性腺及腎臟中高表達, 且該基因對黃鱔精母細胞及卵母細胞的生長及成熟起一定的作用。綜上, 我們推測基因和基因在性腺和腎臟發(fā)育過程及功能維持方面發(fā)揮重要作用。
4 結論
本研究用RACE方法獲得了圓斑星鰈和基因全長, 分別命名為基因和基因, 其均具有4個鋅指結構, 編碼 KTS三肽。基因表達結果表明:和基因主要在圓斑星鰈性腺中表達, 精巢的表達高于卵巢, 腎臟的表達量顯著高于其他組織, 推測基因和基因在性腺和腎臟發(fā)育過程及功能方面均發(fā)揮重要作用;在早期發(fā)育階段,基因在圓斑星鰈原始生殖細胞分化過程及性腺發(fā)育中發(fā)揮重要作用。基因在性腺分化過程的發(fā)揮作用仍需進一步研究。
江東能, 2016. Gsdf和Wt1在羅非魚性別分化和性腺發(fā)育中的功能研究. 重慶: 西南大學博士學位論文, 53—74
孫近近, 2016. 牙鲆性腺相關miR-200b及dazl、wt1a基因的鑒定與表達. 上海: 上海海洋大學碩士學位論文, 29—53
楊珍珍, 邊 力, 張 巖等, 2020. 圓斑星鰈基因的克隆與表達分析. 漁業(yè)科學進展, 41(2): 103—112
張 紅, 陳松林, 劉 洋等, 2014. 半滑舌鰨基因的克隆與性別分化期的表達分析. 中國水產(chǎn)科學, 21(1): 26—36
張樂樂, 2018. 圓斑星鰈性腺分化過程及性別相關基因3、9的研究. 上海: 上海海洋大學碩士學位論文, 12—28
張樂樂, 邊 力, 常 青等, 2018. 圓斑星鰈9基因的克隆與表達. 漁業(yè)科學進展, 39(6): 72—80
胡 青, 楊嬌艷, 高 宇等, 2014. 黃鱔WT1基因序列分析及在性腺發(fā)育過程中的表達. 華中農(nóng)業(yè)大學學報, 33(1): 73—79
柳學周, 徐永江, 廖梅杰等, 2013. 圓斑星鰈促性腺激素釋放激素基因克隆及表達特性. 中國水產(chǎn)科學, 20(1): 12—24
康現(xiàn)江, 寧靜允, 穆淑梅等, 2010. 半滑舌鰨早期胚胎性腺原基分化的組織學. 河北大學學報(自然科學版), 30(3): 296—300, 306
Bollig F, Mehringer R, Perner B, 2006. Identification and comparative expression analysis of a secondgene in zebrafish. Developmental Dynamics, 235(2): 554—561
Davies J A, Ladomery M, Hohenstein P, 2004. Development of an siRNA-based method for repressing specific genes in renal organ culture and its use to show that the Wt1 tumour suppressor is required for nephron differentiation. Human Molecular Genetics, 13(2): 235—246
Dong L H, Pietsch S, Englert C, 2015. Towards an understanding of kidney diseases associated withmutations. Kidney International, 88(4): 684—690
Gessler M, Poustka A, Cavenee W, 1990. Homozygous deletion in Wilms tumours of a zinc-finger gene identified by chromosome jumping. Nature, 343(6220): 774—778
Hammes A, Guo J K, Lutsch G, 2001. Two splice variants of the Wilms’ tumor l gene have distinct functions during sex determination and nephron formation. Cell, 106(3): 319—329
Hossain A, Saunders G F, 2001. The human sex-determining geneis a direct target of. The Journal of Biological Chemistry, 276(20): 16817—16823
Hossain A, Saunders G F, 2003. Role of Wilms Tumor 1 (WT1) in the transcriptional regulation of the Mullerian-Inhibiting substance promoter. Biology of Reproduction, 69(6): 1808—1814
Klamt B, Koziell A, Poulat F, 1998. Frasier syndrome is caused by defective alternative splicing ofleading to an altered ratio of WT1 +/-KTS splice isoforms. Human Molecular Genetics, 7(4): 709—714
Klüver N, Herpin A, Braasch L, 2009. Regulatory back-up circuit of medaka Wt1 co-orthologs ensures PGC maintenance. Developmental Biology, 325(1): 179—188
Kreidberg J A, Sariola H, Loring J M, 1993. WT-1 is required for early kidney development. Cell, 74(4): 679—691
Moore A W, McInnes L, Kreidberg J A, 1999. YAC complementation shows a requirement forin the development of epicardium, adrenal gland and throughout nephrogenesis. Development, 126(9): 1845—1857
Morrison A A, Viney R L, Ladomery M R, 2008. The post-transcriptional roles of WT1, a multifunctional zinc-finger protein. Biochimica et Biophysica Acta (BBA) - Reviews on Cancer, 1785(1): 55—62
Pelletier J, Schalling M, Buckler A J, 1991. Expression of the Wilms’ tumor gene WT1 in the murine urogenital system. Genes & Development, 5(8): 1345—1356
Sharma P M, Yang X, Bowman M, 1992. Molecular cloning of rat Wilms’ Tumor complementary DNA and a study of messenger RNA expression in the urogenital system and the brain. Cancer Research, 52(22): 6407—6412
Tamura K, Peterson D, Peterson N, 2011. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution, 28(10): 2731—2739
Wiener J S, Marcelli M, Lamb D J, 1996. Molecular determinants of sexual differentiation. World Journal of Urology, 14(5): 278—294
Wilhelm D, Englert C, 2002. The Wilms tumor suppressor WT1 regulates early gonad development by activation of. Genes & Development, 16(14): 1839—1851
Zemann A, de Bekke A O, Kiefmann M, 2006. Evolution of small nucleolar RNAs in nematodes. Nucleic Acids Research, 34(9): 2676—2685
Zheng Q S, Wang X N, Wen Q, 2013.deficiency causes undifferentiated spermatogonia accumulation and meiotic progression disruption in neonatal mice. Reproduction, 147(1): 45—52
CLONING AND EXPRESSION OF THEGENE IN SPOTTED HALIBUT ()
YANG Zhen-Zhen1, 2, BIAN Li1, 3, ZHANG Yan1, CHEN Si-Qing1, 3, CHANG Qing1, ZHANG Sheng-Nong1, LIU Chang-Lin1, GE Jian-Long1
(1. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China; 2. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306, China; 3. Laboratory for Marine Fisheries Science and Food Production Processes, Pilot National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266071, China)
The spotted halibut () is a rare and valuable marine fish species that inhabits the coast of the northern China. Mature females are much larger than males because of their faster growth. It will create substantial economic benefits to establish an all-female breeding technique for. A better understanding of sex-related genes will contribute to the improvement of a single-sex breeding technique. In this study, we successfully isolated thegene of, which is namedand. The total length ofwas3263bp, including a 1245bp open reading frame (ORF), encoding 415 amino acids, the 5′UTR was 372bp and the 3′UTR was 1640bp. The total length ofwas 2312bp, including a 1281bp open reading frame (ORF), encoding 427 amino acids, the 5′UTR was 369bp and the 3′UTR was 659bp. Through biological analysis, thegene encoded amino acids, with a predicted molecular weight of 46.2kDa and an isoelectric point of 9.24, no transmembrane structure or signal peptide site was detected, there were four zinc finger structures at the end of ORF and encode KTS. Thegene encoded amino acids, with a predicted molecular weight of 46.95kDa and an isoelectric point of 8.99, no transmembrane structure or signal peptide site was detected, there were four zinc finger structures at the end of ORF and encode KTS. Real-time fluorescence quantitative PCR technique was used to analyze the expression patterns of thegene at different stages of embryo and larvae. The results show that the expression level ofandgenes in gonads was significantly higher than other tissues’, and the expression level in the testis was significantly higher than in the ovary. Therefore, we speculated thatandgenes play important roles in gonad and kidney development and function. In the early stage of development,gene weakly expressed before the early gastrula, then the expression level gradually rose to the highest level in the neural embryo stage, followed by a decline until the hatching stage. It is speculated thatgene might played a role in the differentiation process of primitive germ cells and gonad development.
;; gene cloning; expression analysis
*中國水產(chǎn)科學研究院黃海水產(chǎn)研究所基本科研業(yè)務費項目, 20603022016005號。楊珍珍, 碩士研究生, E-mail: 1163603557@qq.com
陳四清, 博士生導師, 研究員, E-mail: chensq@ysfri.ac.cn
2020-03-04,
2020-06-08
S917.4
10.11693/hyhz20200300055
