不同栽培模式對夏玉米冠層結構及光合性能的影響
2020-11-26 12:35:26李靜王洪章許佳詣劉鵬張吉旺趙斌任佰朝
中國農業科學 2020年22期
關鍵詞:結構
李靜,王洪章,許佳詣,劉鵬,張吉旺,趙斌,任佰朝
(山東農業大學農學院/作物生物學國家重點實驗室,山東泰安 271018)
0 引言
【研究意義】玉米作為我國第一大糧食作物,在維護國家糧食安全中發揮重要作用,高產栽培模式探究對進一步提高玉米單產保證增產至關重要[1],合理增大種植密度是玉米高產的有效途徑[2]。光照是影響作物生長發育的關鍵環境因子,它通過影響葉片發育、激活光合作用關鍵酶、增強生理反應需要的能量供應等過程調節作物光合作用[3]。密植群體易造成葉片間相互遮蔭[4],導致群體內光分布不合理,影響葉片結構發育,直接影響葉片光合性能及群體光能利用效率,形成不合理的群體冠層結構[5]。葉綠體是作物進行光合作用的場所,群體內光照條件會對葉綠體結構產生影響,進而影響葉片光合性能[6]、同化物的積累及產量形成[7]。探究不同栽培模式群體內夏玉米冠層結構特性、葉片發育特點與光合性能的差異,闡明不同群體冠層結構對夏玉米葉片超微結構、葉片光合性能的影響,對進一步研發夏玉米高產栽培技術具有重要意義。【前人研究進展】增大種植密度、調整氮肥用量及施肥方式是研究較多的提高產量的栽培措施。單純增大種植密度,會導致玉米葉片之間相互遮蔭,中下部葉片受光不足,加快下部葉片及根系衰老速度[4,8]。氮肥合理運籌[9]、優化株行距配置[10-12]可調節葉片著生狀態,構建合理的群體冠層結構,中下部葉片接收更多光能[13],改善群體內光環境。綜合密植和優化肥料運籌的栽培措施可提高氮肥利用率[14]和光能生產效率[15]。光照會通過影響葉片發育、調節葉綠體超微結構,進而影響葉片光合作用[16],葉片受光不足時,葉綠體排布變得不規則,結構膨脹、基粒片層排列異常、基粒個數和片層數減少,葉綠體膜開始溶解,導致葉片早衰[17-18],影響其光合作用。【本研究切入點】盡管有較多學者對夏玉米群體結構、光合性能進行了研究,但多集中在密度、肥料等單一措施的影響,關于不同栽培措施對夏玉米群體發育及光合性能的綜合影響的研究相對較少,難以系統解釋夏玉米生長發育及產量形成過程中所受外界因素的綜合影響,難以精準確定增產措施。【擬解決的關鍵問題】本文通過研究夏玉米不同栽培模式的群體冠層特性、葉片發育狀況及其光合能力,從所處光環境及葉綠體結構兩方面分析不同群體內葉片光合能力及產量差異的原因,為進一步制定可構建高效冠層結構的夏玉米高產栽培模式提供理論依據。
1 材料與方法
1.1 試驗區概況
本試驗于2018—2019年在山東省泰安市岱岳區馬莊鎮(35°58′41″N,116°58′22″E)進行,播前土壤耕層養分含量為有機質 9.22 g·kg-1,堿解氮 70.54 mg·kg-1,速效磷 35.75 mg·kg-1,速效鉀 137.57 mg·kg-1。該區域為溫帶季風性氣候,種植制度為一年兩熟,夏玉米生育期內(6—10月)日平均氣溫、日光合有效輻射量、降雨量詳見圖 1。室內試驗在山東農業大學作物生物學國家重點實驗室進行。
1.2 試驗設計
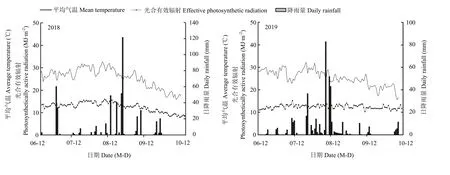
圖1 玉米生育期內試驗地的日平均氣溫、日光合有效輻射、日降雨量Fig. 1 Daily average temperature, effective photosynthetic radiation, and daily rainfall of the test site during the maize growth period
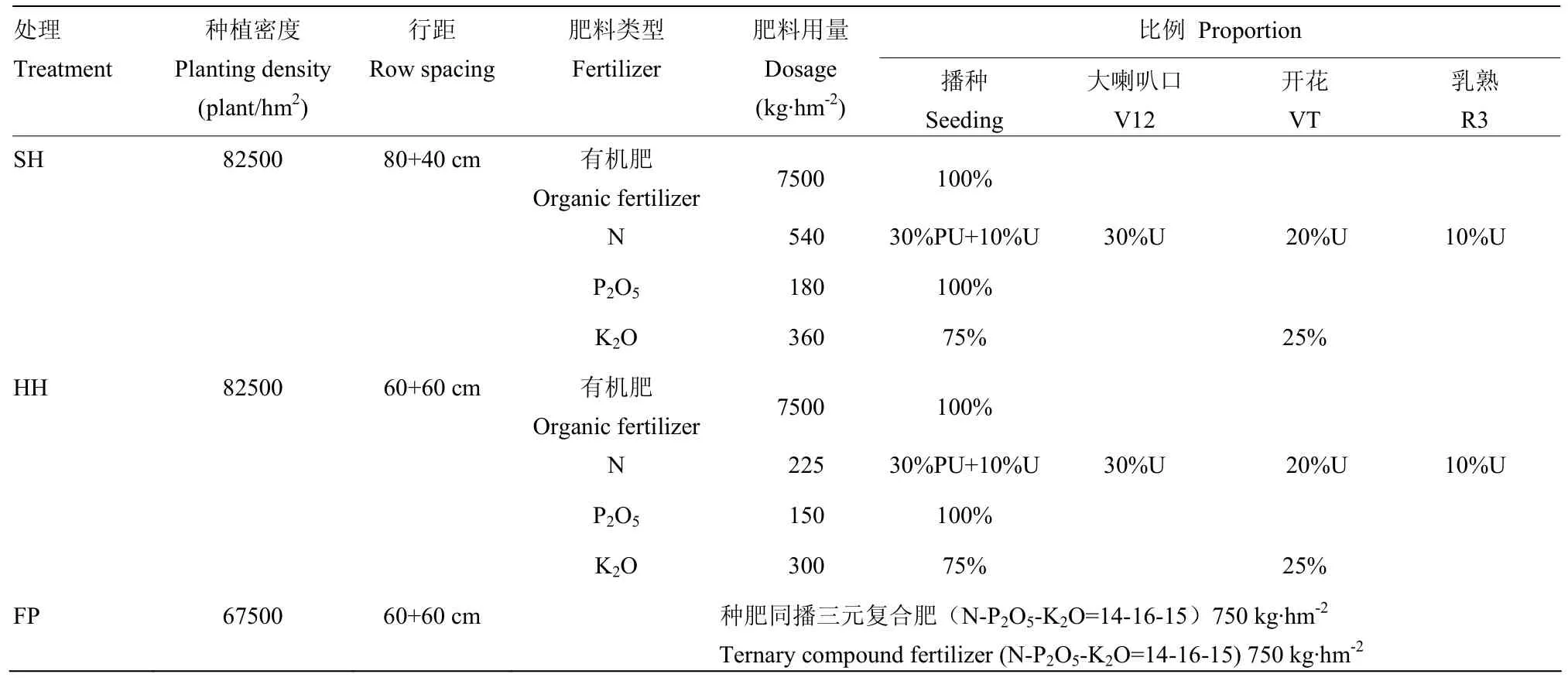
表1 密度及肥料運籌Table 1 Density and fertilizer management
本試驗以玉米品種登海605為材料,在同一地塊結合種植密度、肥水管理及種植方式設置超高產(SH)、高產高效(HH)、農戶習慣(FP)3種栽培模式,各處理的具體種植密度、肥料管理詳見表1。試驗所用氮肥為包膜緩控尿素(PU,含N 42%)和普通尿素(U,含N 46%),磷肥為過磷酸鈣(含P2O512%),鉀肥為硫酸鉀(含K2O 51%),有機肥料為山東友邦肥業科技有限公司生產的商品有機肥(含有機碳(干基)304 g·kg-1、P2O531.2 g·kg-1、K2O 30.4 g·kg-1、C/N為11.2)。試驗為隨機區組設計,各試驗小區長18 m、寬12 m,3次重復,各小區之間設立1 m的隔離帶。在小麥收獲后旋耕(25 cm)滅茬,整平播種。夏玉米生育期內根據土壤墑情采用微噴帶統一噴灌,遇澇及時排水。統一采用病蟲害預防方案進行病蟲害防控。
1.3 測定項目與方法
1.3.1 地上部干物質積累量 玉米生長至拔節期(V6)、大喇叭口期(V12)、吐絲期(R1)、吐絲后30 d(R1+30 d)、成熟期(R6)時,取各處理內長勢一致的玉米植株5株,將植株按莖稈、葉片、雌穗(穗軸和籽粒)、雄穗、苞葉分開,105℃殺青30 min,80℃烘至恒重并稱重,計算不同生育時期群體干物質積累量。
1.3.2 莖葉夾角、葉向值和葉面積指數 R1期分別于各小區選取長勢一致的植株10株,穗位葉、穗位上葉及下葉為中部葉層,中部葉層以上為上部葉層,中部葉層以下為下部葉層。測量各層葉片葉長(Li)、沿葉片方向葉基部至葉片最高點的距離(Lf),用量角器測量莖葉夾角,并計算葉向值(LOV)。
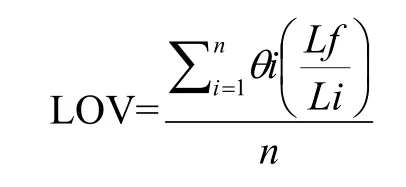
式中,θi為葉片與水平面夾角,Lf為沿葉片方向葉基部至葉片最高點的距離,Li為葉片伸直時長度,n為葉片數。
拔節期(V6)每個處理內選取長勢一致的植株5株,定株。分別于V6、V12、R1、R1+30 d、R6期測量所選定植株綠葉的葉長、葉寬。單株葉面積(m2)=∑完全展開葉的葉長(m)×葉寬(m)×0.75+∑未完全展開葉的葉長(m)×葉寬(m)×0.5;葉面積指數(LAI)=(單株葉面積×單位面積株數)/占地面積。
1.3.3 群體冠層結構 采用大田切片法測定R1期群體冠層垂直結構[16],在每個處理內選取2 m2(1 m×2 m)內的植株,從基部起以40 cm為每層高度,按照層高分層割取樣品,每層按照莖稈、葉片、雌穗、雄穗分樣。在烘箱內105℃殺青30 min,80℃烘干至恒重,并稱重。
1.3.4 冠層光能截獲率 采用SunScan(Delta,英國)冠層分析儀,在玉米生長至 R1期,選擇晴朗無云的天氣,于10:00—13:00測定各處理群體冠層內不同葉層及冠層頂部的光合有效輻射(PAR)。
光能截獲率=PARi/TPAR×100%
式中,PARi為某葉層的PAR;TPAR為冠層頂部PAR。
1.3.5 葉綠體超微結構 在玉米R1期,在各處理中選取長勢一致的植株,在其穗位葉片中段(避開葉脈)割取邊長為0.5 cm的方形葉片,用3.3%戊二醛固定液(0.1 mol·L-1pH 7.2的磷酸緩沖液配制)固定,抽真空,4℃下固定24 h。用磷酸緩沖鹽溶液洗3次,每次20 min,再將材料轉移到鋨酸中,4℃固定4 h,用磷酸緩沖鹽溶液沖洗 3次。常規梯度(45%、55%、70%、85%、95%、100%)乙醇系列脫水,環氧丙烷置換。Epon812樹脂包埋,70℃下聚合 8 h,LKB-V型切片機切割超薄切片,經醋酸雙氧鈾和檸檬酸鉛雙重染色后,用日立-600 型透射電鏡隨機選取視野觀察并拍照。
1.3.6 凈光合速率 在玉米R1期,選擇晴朗無云的天氣,于9:00—11:00使用CIRAS-Ⅲ(PP System,美國)光合儀,每個處理選具有代表性植株 10—15株,測定其穗位葉凈光合速率(Pn)。
1.3.7 籽粒產量及產量構成因素 于玉米R6期,在每個處理內選取5個9 m2(5 m×1.8 m)具有代表性玉米帶,將其全部果穗收獲、晾曬,測定其產量和產量構成因素。
1.4 數據分析與作圖
采用 Microsoft Excel 2010進行數據處理;采用DPS進行數據統計分析,用 LSD法做顯著性分析(α=0.05);用Sigmaplot 14.0作圖。
2 結果
2.1 不同栽培模式對夏玉米群體冠層結構的影響
栽培方式影響玉米葉片著生狀態及生長發育,各葉層莖葉夾角均表現為FP>SH>HH,葉向值則表現為SH>HH>FP,SH和HH模式群體上部葉片上沖,利于中、下部葉片接收光能(表2)。全生育期內葉面積指數呈現先增大后降低的變化趨勢,在吐絲期達到最大值,各個生育期內均表現為SH>HH>FP,在吐絲期后處理之間呈顯著性差異(圖2)。相比于FP模式,SH、HH模式群體中單位面積內干物質積累量顯著提高;SH、HH模式群體內植株干重在120 cm以下的比例有顯著增加,穗位葉及以下葉片的長勢較好(圖3),2年試驗不同處理間變化趨勢一致。
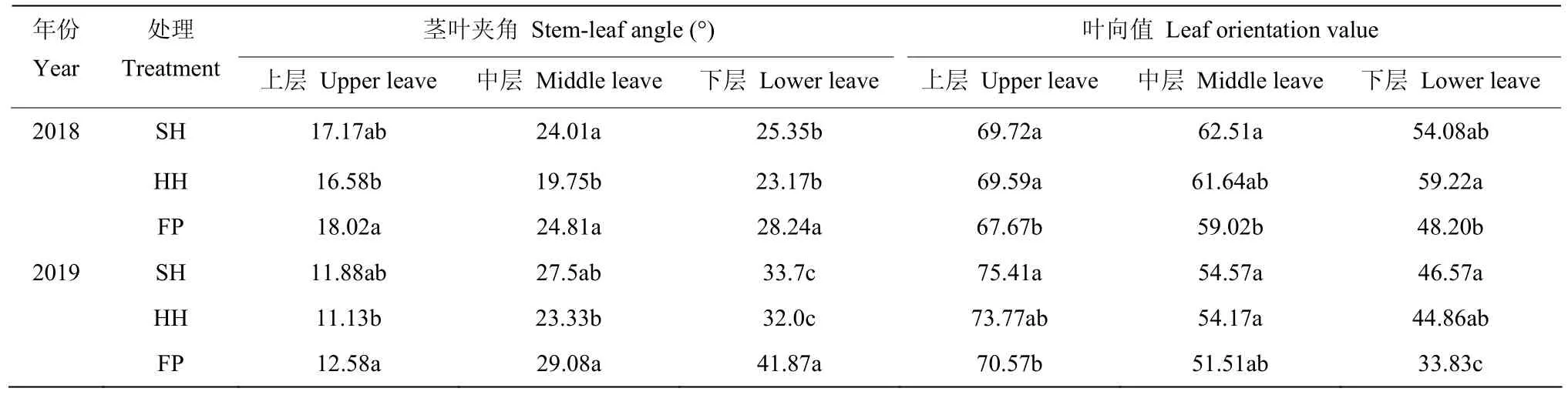
表2 不同栽培模式對夏玉米葉片著生狀態的影響Table 2 Effects of different cultivation modes on the implantation status of summer maize
2.2 不同栽培模式夏玉米群體內光分布的差異
各冠層高度光能截獲率均表現為SH>HH>FP,SH模式的光能截獲率最大,漏光損失最小,HH模式次之,FP模式最易造成漏光損失,2年試驗規律一致。2年試驗SH、HH、FP模式中部葉層較上部葉層光能截獲率分別平均增大39.67%、47.21%、45.04%,HH模式中、上部葉層結構最優(圖4)。
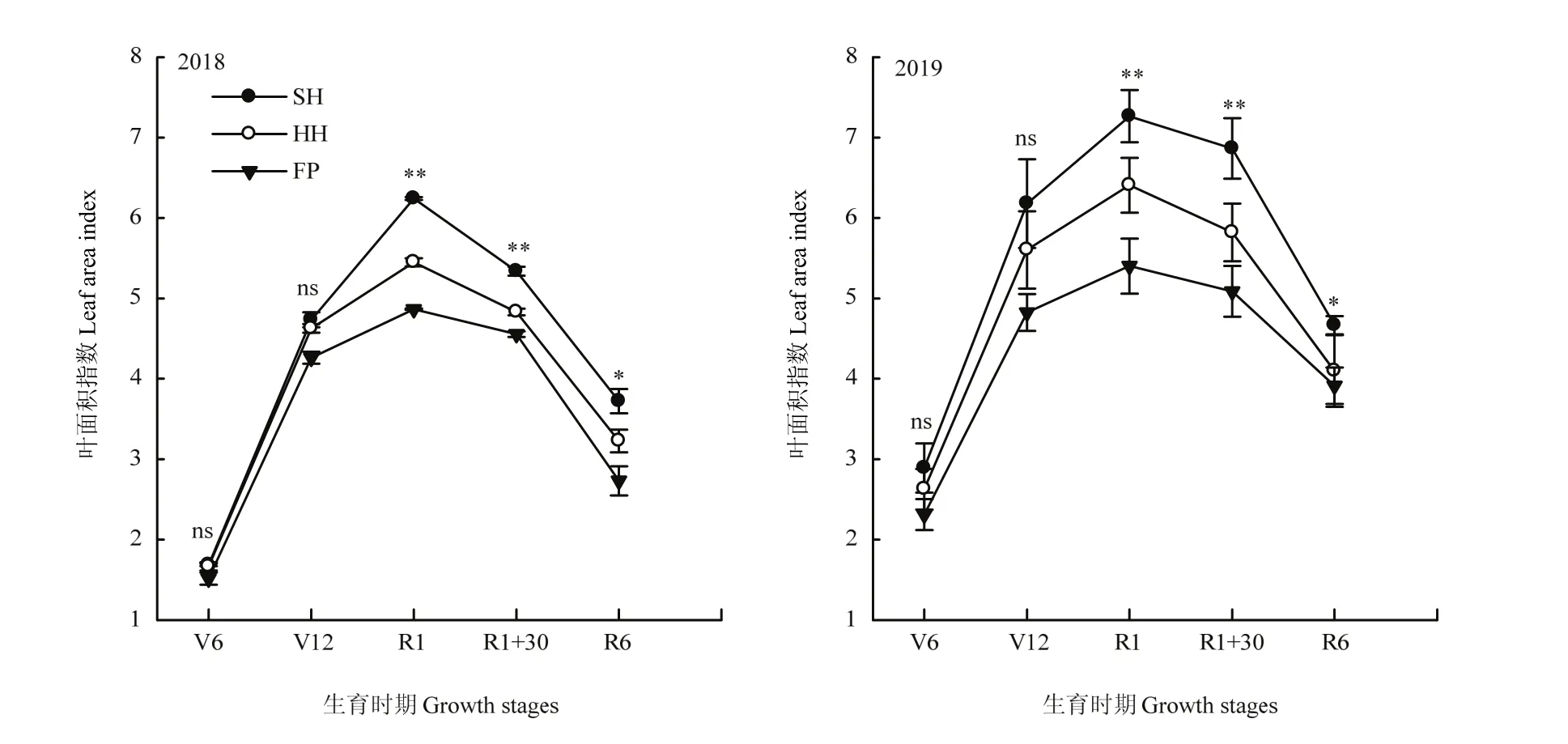
圖2 不同栽培模式群體葉面積指數在各生育時期的變化Fig. 2 Changes of leaf area index of populations in different cultivation modes at different growth stages
2.3 不同栽培模式對夏玉米葉綠體超微結構的影響
不同處理葉片葉綠體在外形上無明顯差異,均為正常的“紡錘形”;SH、HH模式的葉綠體內部結構發育良好,FP模式發育相對較差。SH和HH模式的類囊體發育良好數目較多,基粒片層和基質片層排列整齊、清晰,細胞內膜結構完整;FP模式的葉綠體中類囊體發育相對較差,未發育完全的類囊體數目占比較大,且其基質片層模糊,且排列不齊(圖5)。
2.4 不同栽培模式夏玉米穗位葉凈光合速率的差異
夏玉米不同栽培模式的穗位葉吐絲期凈光合速率表現為 SH>HH>FP,且處理之間差異顯著,2年大田試驗呈現相同的變化趨勢。2年試驗中,SH模式較FP模式分別增大26.09%、24.34%;HH模式較 FP模式分別增大 15.07%、18.02%;SH模式較HH模式分別增大9.57%、5.48%(圖6)。
2.5 不同栽培模式對夏玉米干物質積累量及產量的影響
不同栽培模式干物質積累量表現為 SH>HH>FP。在2年試驗中,干物質積累量SH、HH模式較FP模式平均增加27.74%、7.45%;對比玉米開花前、開花后干物質積累量的比例,與FP模式相比,SH、HH模式在開花后干物質積累量顯著提高,SH、HH、FP模式干物質在吐絲后積累量占全生育期的比例分別為 64.38%、60.03%、58.04%(表 3)。SH、HH模式較 FP模式可顯著提高夏玉米籽粒產量,SH模式較FP模式平均增產14.20%,HH模式較FP模式平均增產4.91%。與FP模式相比,SH、HH模式的穗粒數及千粒重降低,單位面積穗數顯著提高。SH、HH模式籽粒產量提高的主要原因是單位面積穗數的增加(表4)。
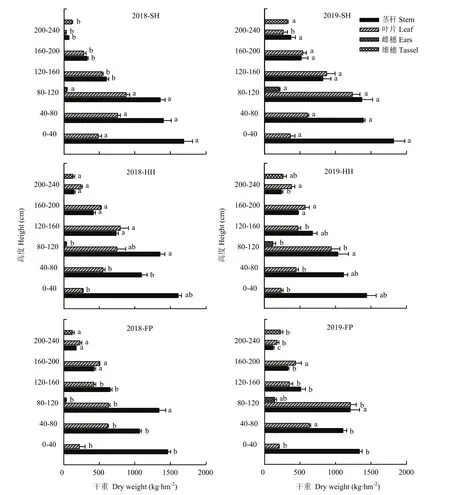
圖3 不同栽培模式下夏玉米吐絲期冠層結構垂直分布Fig. 3 Vertical distribution of canopy structure of summer maize during silking stage under different cultivation modes
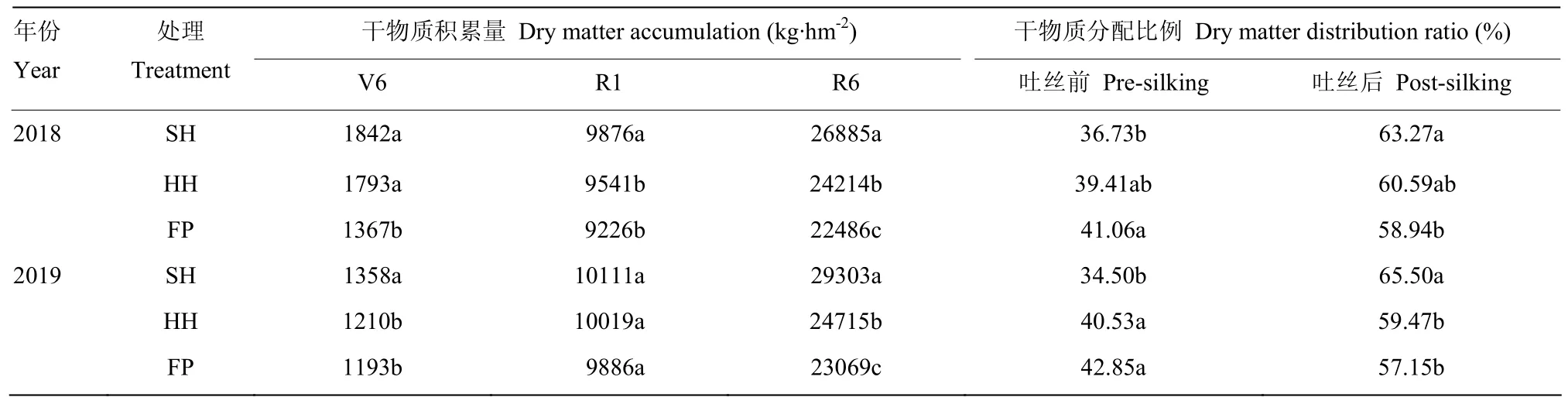
表3 不同栽培模式夏玉米干物質積累量及吐絲前后分配比例Table 3 Dry matter accumulation and distribution ratio of summer maize before and after silking in different cultivation modes
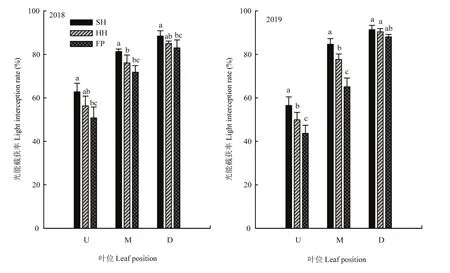
圖4 不同栽培模式群體光能截獲率Fig. 4 Light energy interception rate of different cultivation modes
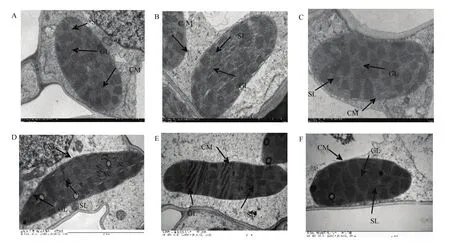
圖5 不同栽培模式群體內葉片葉綠體的發育狀況Fig. 5 Development of leaf chloroplasts in different cultivation mode populations

圖6 不同栽培模式中夏玉米穗位葉吐絲期凈光合速率Fig. 6 Net photosynthetic rate of panicle leaves of summer maize during silking stage under different cultivation modes
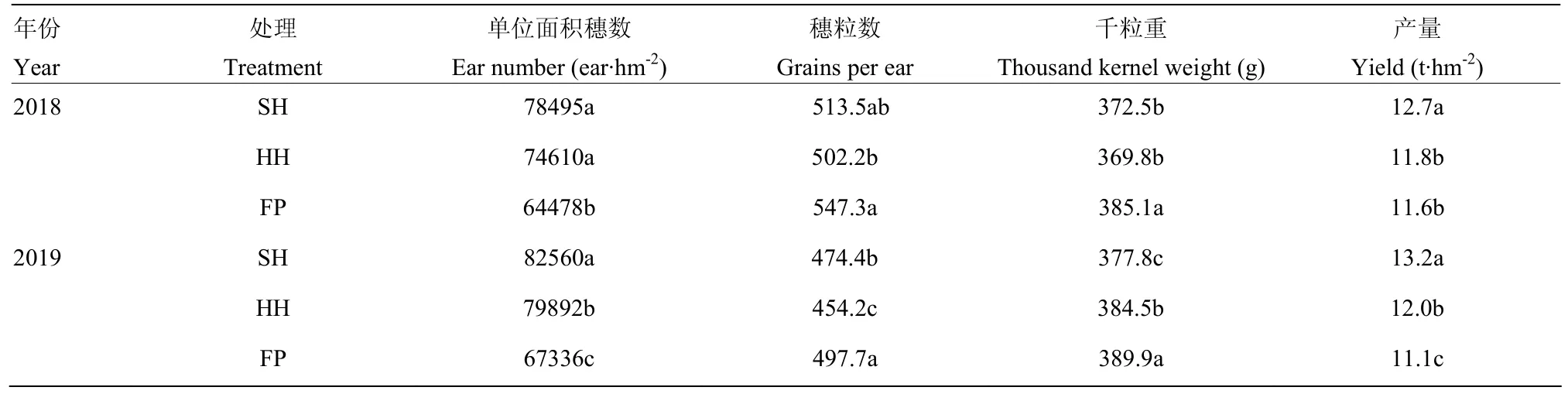
表4 不同栽培模式玉米籽粒產量及產量構成因素Table 4 Grain yield and yield components under different cultivation modes
3 討論
葉片是植株進行光合作用的主要場所,是植物吸收、利用光能的最重要器官[19],葉面積指數是反映植株生長發育和光能利用率的重要指標,夏玉米要實現高產就要保持較高的葉面積指數[20-23],調節葉片著生狀態和伸展方向,構建合理的冠層結構[21,24]。合理的冠層結構有利于葉片發育、形成適宜的群體內環境,可接收充足的光照,提高葉光合能力,從而提高干物質積累量及產量[25]。種植密度過高,上部葉片發育良好、生長健壯[26],導致植株間葉片相互遮蔭[4],群體內光照不足,中下部葉片光能利用能力降低。適宜的肥料運籌、種植方式可調節植株生長發育狀況[13,27],調整群體冠層結構使上層葉片上沖,群體內不同高度葉層的光照分布趨于合理化,保證葉片處于適宜的光環境[28],以提高光能利用率。本研究中,通過統籌各栽培措施構建的2種高產栽培模式,與農戶生產模式相比具有較為合理的冠層結構。SH、HH模式葉面積指數顯著大于 FP模式,且具有更長的葉面積指數高值持續期(圖 2)。SH、HH模式群體上層葉片莖葉夾角小于FP模式、葉向值大于FP模式(表2)。對于120 cm以下葉片干物質積累量,SH模式顯著大于FP模式,HH模式大于FP模式。SH、HH模式調節了上部葉片的著生狀態,使上部葉片上沖,中下部葉層可接受充足的光能,既防止漏光損失、又減緩了高密植群體內葉片相互遮蔭,SH模式中、下層葉片生長發育狀況顯著優于HH和FP模式,這是SH、HH模式可實現高產但增產水平不同的重要原因之一。
葉綠體是葉片進行光合作用的主要細胞器,葉綠體超微結構與植株葉片光合性能密切相關。發育良好的葉綠體呈紡錘形,靠近細胞膜排布在細胞內,類囊體結構完整,葉綠體基粒數目增加、基粒片層和基質片層排列緊密[29]。當受光不足時,葉片細胞及葉綠體結構會被破壞,導致光合系統受損,從而影響光合性能[18,30]。施用適量的氮、鉀肥可保持番茄[31]、甘蔗[32]的葉綠體超微結構。在本研究中,SH、HH模式的葉綠體內部結構發育良好,FP模式發育相對較差。SH和HH模式的類囊體發育良好且數目較多,基粒片層和基質片層排列整齊、清晰,細胞內膜結構完整;FP模式的葉綠體類囊體發育相對較差,未發育完全的比例較高,基質片層模糊且排列不齊(圖 5)。進一步說明SH、HH模式2個群體冠層結構適宜,群體內光環境良好,未造成葉片之間相互遮蔭影響葉綠體結構發育,而且SH、HH模式的葉綠體結構更有利于進行光合作用。HH模式的葉綠體結構發育良好,但其凈光合速率顯著低于SH模式,這可能與光合相關酶的活性有關。
與農戶生產模式相比,通過優化種植密度和肥料運籌等措施構建的超高產模式和高產高效模式可顯著促進根系發育、提高氮肥利用率[14]和光能生產效率[15]。本試驗結果表明,超高產模式和高產高效模式可實現增產的原因是優化了群體冠層結構,促進植株及葉綠體結構的發育,延緩葉片衰老,提高了葉片光合能力。
4 結論
相比于農戶習慣管理模式,超高產模式、高產高效模式實現產量提高的原因是優化了群體冠層結構,促進葉片的發育,保證葉綠體結構的完整性,顯著提高了葉片光合能力。與超高產模式相比,高產高效模式減小肥料施用量,其群體結構適宜且葉片發育良好,是更有助于利用光能的栽培模式。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
