貴州小麥品種(系)粒重相關(guān)基因TaCwi-A1、TaSus2-2B和TaGW2-6A等位變異類型鑒定
2020-12-11 09:10:33楊雪敏李魯華任明見徐如宏
南方農(nóng)業(yè)學(xué)報(bào) 2020年9期
楊雪敏 李魯華 任明見 徐如宏


摘要:【目的】鑒定貴州小麥品種(系)中粒重相關(guān)基因TaCwi-A1、TaSus2-2B和TaGW2-6A的等位變異類型,篩選含高粒重基因型的小麥品種(系),為貴州小麥粒重的遺傳改良和高粒重品種選育提供參考。【方法】以252份小麥品種(系)為材料,分別利用TaCwi-A1、TaSus2-2B和TaGW2-6A基因的分子標(biāo)記(CWI22/CWI21、TaSus2-1/TaSus2-2和Hap-6A-P1/Hap-6A-P2)引物進(jìn)行PCR擴(kuò)增,利用毛細(xì)管電泳檢測擴(kuò)增產(chǎn)物,鑒定分析這3個(gè)基因的等位變異類型及分布頻率,并結(jié)合籽粒性狀測定結(jié)果,篩選含高粒重基因變異類型的小麥種質(zhì)。【結(jié)果】252份小麥種質(zhì)材料的粒重平均值為39.87 g,其中,有56份材料屬于大粒種質(zhì)(>45.00 g),僅有13份檢測到等位變異;173份材料屬于中粒種質(zhì)(30.00~45.00 g),有24份檢測到等位變異;23份材料屬于小粒種質(zhì)(<30.00 g),均未檢測到等位變異。252份小麥材料中,含有TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位變異的材料37份,占供試材料總數(shù)的14.7%,其中TaCwi-A1基因等位變異類型材料12份(包括TaCwi-A1a變異類型8份,TaCwi-A1b變異類型4份),占供試材料總數(shù)的4.8%;TaSus2-2B基因等位變異類型材料16份(包括TaSus2-2BH變異類型材料2份,TaSus2-2BL變異類型材料14份),占供試材料總數(shù)的6.4%;TaGW2-6A基因等位變異類型材料10份(包括Hap-6A-A變異類型4份,Hap-6A-G變異類型6份),占供試材料總數(shù)的4.0%;等位變異組合類型僅有1份材料(惠光2-2-2),為TaCwi-A1b/HAP-6A-A,占供試材料總數(shù)的0.4%。平均粒重最高的變異類型為TaCwi-A1b,其次是HAP-6A-G變異類型。對于平均粒重,TaCwi-A1b變異類型顯著高于TaCwi-A1a變異類型(P<0.05,下同),Hap-6A-G變異類型顯著高于Hap-6A-A變異類型,TaSus2-2BH變異類型也高于TaSus2-2BL變異類型,但差異不顯著(P>0.05)。【結(jié)論】從貴州小麥品種(系)檢測到的粒重基因等位變異整體較少,表明貴州小麥種質(zhì)遺傳多樣性較低,高粒重品種的選育工作開展不夠,今后應(yīng)重視小麥粒重基因等位變異綜合效應(yīng)研究。鑒定出含有TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位變異類型、粒重>45.00 g的品種(系)13份,可應(yīng)用于貴州小麥粒重的遺傳改良和高粒重品種選育。
關(guān)鍵詞: 小麥;粒重;基因;等位變異;分子標(biāo)記;毛細(xì)管電泳;鑒定
中圖分類號: S512.103.53? ? ? ? ? ? ? ? ? ? ? ? ? ? ?文獻(xiàn)標(biāo)志碼: A 文章編號:2095-1191(2020)09-2071-11
Identification and analysis of allele variation types of kernel weight related genes TaCwi-A1,TaSus2-2B and TaGW2-6A of Guizhou wheat varieties(lines)
YANG Xue-min, LI Lu-hua, REN Ming-jian, XU Ru-hong*
(College of Agriculture, Guizhou University/Guizhou Branch of National Wheat Improvement Center,
Guiyang? 550025,China)
Abstract:【Objective】The allele variation types of kernel weight related genes TaCwi-A1,TaSus2-2B and TaGW2-6A in Guizhou wheat varieties(lines) were identified,and wheat varieties(lines) containing high kernel weight genotypes were screened to provide reference for genetic improvement of Guizhou wheat kernel weight and breeding of high kernel weight varieties. 【Method】With 252 wheat varieties(or lines) as the materials,using molecular marker of genes TaCwi-A1, TaSus2-2B and TaGW2-6A(CWI22/CWI21, TaSus2-1/TaSus2-2 and Hap-6A-P1/Hap-6A-P2) primers for PCR amplification,amplification products were detected using capillary electrophoresis,allele variation types and distribution of the three genes were identified, combined with the measuring results of grain traits, wheat germplasm with high kernel weight mutation types were screened. 【Result】The average grain weight of 252 wheat germplasm materials was 39.87 g. Among them,56 materials belonged to large kernel germplasms(>45.00 g),and only 13 of them detected allele variation.Among the 173 medium kernel materials(30.00-45.00 g),allele variation was detected in 24. The rest 23 materials belonged to small kernel germplasm(<30.00 g),and no allele variation was detected.Among the 252 wheat materials,37 contained allelic variation of TaCwi-A1, TaSus2-2B and TaGW2-6Agenes,accounting for 14.7% of the total tested mate-rials. Among them,12 contained allele variation of TacWI-A1 gene(including 8 of TaCwi-A1a and 4 of TaCwi-A1b),accou-nting for 4.8% of the total tested materials. There were 16 allele materials of TaSus2-2B gene(including 2 allele materials of TaSus2-2BH and 14 allele materials of TaSus2-2BL),accounting for 6.4% of the total number of tested materials. There were 10 allele variation types of TAGW2-6A(including 4 HAP-6A-A variation types and 6 HAP-6A-G variation types),accou-nting for 4.0% of the total number of materials tested.There was only one allele variation combination type of material(Huigang 2-2-2),which was Tacwi-A1b/HAP-6A-A,accounting for 0.4% of the total number of materials tested. The varia-tion type with the highest mean kernel weight was TACwi-A1b,the second type was HAP-6A-G variation. For average grain weight,the variation type of Tacwi-A1b was significantly higher than that of Tacwi-A1a(P<0.05,the same as below),the variation type of HAP-6A-G was significantly higher than that of HAP-6A-A,and the variation type of TaSus2-2BH was also higher than that of TaSus2-2BL,but the difference was not significant(P>0.05). 【Conclusion】The allele variation diversity of grain weight genes detected from Guizhou wheat varieties(lines) is relatively low,indicating that the genetic diversity of Guizhou wheat germplasm is low,and the breeding of high-kernel-weight varieties is not enough. In the future,more attention should be paid to the comprehensive effects of the allele variation of wheat weight. Thirteen varie-ties(lines) containing allele types of TaCwi-A1,TaSus2-2B and TaGW2-6A genes with grain weight of >45.00 g were identified,which can be used for genetic improvement of Guizhou wheat grain weight and breeding of high-kernel-weight varie-ties.
Key words: wheat; kernel weight; gene; allele variation; molecular markers; capillary electrophoresis; identification
Foundation item:National Key Research and Development Plan “Seven Crops Breeding” Key Special Project(2017YFD0100900);Introducing Talents Research Project of Guizhou University(GDRJHZ〔2017〕49)
0 引言
【研究意義】小麥(Triticum aestivum L.)是重要的糧食作物,全球約40%的人口以其為主食(http://faost at3.fao.org/home)。小麥高效生產(chǎn)才能實(shí)現(xiàn)全球糧食和營養(yǎng)安全(Hunter et al.,2017)。粒重是小麥產(chǎn)量的主要構(gòu)成因素,當(dāng)小麥粒重增加1 g時(shí),每公頃小麥產(chǎn)量可增加140~160 kg,因此,粒重是小麥產(chǎn)量的重要影響因素,培育高粒重品種是小麥育種的重要目標(biāo)(Tian et al.,2006;Dong et al.,2012)。貴州生態(tài)環(huán)境復(fù)雜,處于氣候潮濕多雨的西南麥區(qū),小麥粒重在不同年度間差異明顯,且粒重是受多個(gè)微效基因控制的數(shù)量性狀,僅靠表型選擇聚合粒重優(yōu)異等位基因較困難(劉永偉等,2017a)。借助分子標(biāo)記技術(shù),對影響粒重的相關(guān)基因進(jìn)行分子標(biāo)記輔助選擇,可極大提高貴州小麥產(chǎn)量育種的效率。因此,利用粒重相關(guān)基因的功能標(biāo)記對小麥性狀改良及高產(chǎn)育種具有重要意義(司文潔等,2019)。【前人研究進(jìn)展】關(guān)于小麥粒重相關(guān)基因的研究已被廣泛報(bào)道,尤其以粒重相關(guān)基因TaCwi-A1、TaSus2-2B和TaGW2-6A的研究較多。韓利明等(2011)利用小麥粒重基因TaGW2-6A位點(diǎn)的CAPS分子標(biāo)記檢測來自21個(gè)國家的745份小麥種質(zhì)材料,結(jié)果發(fā)現(xiàn)TaGW2-6A位點(diǎn)的CAPS分子標(biāo)記能很好區(qū)分其等位變異類型Hap-6A-A和Hap-6A-G,可作為粒重輔助選擇的有效位點(diǎn);Jiang等(2011)克隆得到小麥TaSus2基因,并證明該基因與小麥產(chǎn)量性狀相關(guān)聯(lián),其在胚乳發(fā)育過程中表達(dá)量較高;Su等(2011)通過同源克隆方法從小麥中獲得水稻OsGW2基因的同源基因TaGW2,研究發(fā)現(xiàn)該基因?qū)π←溋V仄鹭?fù)調(diào)控作用;Ma等(2012)也采用同源克隆方法獲得小麥TaCwi-A1基因的全長cDNA,并對其進(jìn)行QTL分析,證明其可解釋粒重4.8%的表型差異;相吉山等(2014)、劉永偉等(2017b)研究發(fā)現(xiàn),含有TaCwi-A1a基因的小麥粒重顯著高于含有TaCwi-A1b基因的小麥粒重;劉永偉等(2017a)研究發(fā)現(xiàn),TaGW2-6A、TaCwi-A1和TaSus2-2B基因與產(chǎn)量相關(guān),對小麥粒寬、粒重等重要產(chǎn)量性狀均發(fā)揮調(diào)控作用,直接影響小麥的產(chǎn)量;時(shí)佳等(2018)、仝靖洋等(2018)利用TaGW2-6A、TaCwi-A1和TaSus2-2B基因的分子標(biāo)記研究其與小麥粒重的相關(guān)性,結(jié)果發(fā)現(xiàn)這些標(biāo)記均能有效預(yù)測小麥粒重大小,可用于小麥粒重的分子輔助選擇研究。【本研究切入點(diǎn)】目前有關(guān)利用分子標(biāo)記鑒定貴州小麥TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位變異類型的研究鮮見報(bào)道。【擬解決的關(guān)鍵問題】將分子標(biāo)記和毛細(xì)管電泳檢測技術(shù)相結(jié)合,對252份小麥品種(系)中TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位變異類型進(jìn)行鑒定,篩選含高粒重基因型的小麥種質(zhì),為小麥高粒重品種選育和分子輔助選擇研究提供參考。
1 材料與方法
1. 1 試驗(yàn)材料
供試材料為252個(gè)小麥品種(系),分別來自貴州貴陽、興義、畢節(jié)、安順和惠水等地,其中部分材料由國家小麥改良中心貴州分中心選育,名稱和來源見表1。主要試劑:DNF-910 dsDNA Kit/s試劑盒購自成都百樂科技有限公司;2×Taq PCR Master Mix反應(yīng)試劑購自生工生物工程(上海)股份有限公司;DL2000 DNA Marker購自貴州彌勒天根生物科技有限公司;905-33-DNA-0~500 bp Mthds試劑盒購于研諾邏輯集成電路設(shè)計(jì)有限公司;50×TAE溶液購自北京索萊寶科技有限公司。主要設(shè)備儀器:移液器(Eppendorf,德國)、Genova Nano分光光度計(jì)(Jenway,英國)、T100TM_Thermal Cycler PCR儀(Bio-Rad,美國)、Fragment Analyzer毛細(xì)管電泳系統(tǒng)(AATI,美國)、高速離心機(jī)(Thermo,美國)、水平電泳槽(DYCP-32B,北京六一儀器廠)、電泳儀(DYY-6C,北京六一生物技術(shù)有限公司)、電熱恒溫水浴鍋(DK-98-Ⅱ,天津泰斯特儀器有限公司)、高速冷凍離心機(jī)(Thermo,美國)和搖床(GS-20,杭州米歐儀器有限公司)。
1. 2 試驗(yàn)方法
1. 2. 1 小麥籽粒性狀測定 于2017─2018年將供試小麥材料種植于國家小麥改良中心貴州分中心科研基地。成熟期,每份材料隨機(jī)選取500粒左右的種子,用SC-G型多功能種子分析儀測量粒重、粒長和粒寬,每份材料的每一項(xiàng)指標(biāo)均重復(fù)測量3次,計(jì)算平均值。
1. 2. 2 基因組DNA提取 每份材料隨機(jī)取10粒種子進(jìn)行發(fā)芽,采用植物基因組DNA提取試劑盒提取其DNA,以分光光度計(jì)檢測其濃度,并稀釋至50 ng/μL備用。
1. 2. 3 分子標(biāo)記檢測 利用前人已開發(fā)的分子標(biāo)記檢測TaCwi-A1、TaSus2-2B和TaGW2-6A基因的等位變異,所用分子標(biāo)記引物的相關(guān)信息如表2所示,由生工生物工程(上海)股份有限公司合成。以上述提取的DNA為模板進(jìn)行PCR擴(kuò)增。反應(yīng)體系20.0 μL:10×PCR Buffer 2.0 μL,2.5 mmol/L dNTPs 0.4 μL,25 mmol/L MgCI2 1.6 μL,10 μmol/L正、反向引物各1.0 μL,2.5 U/μL Taq DNA聚合酶0.5 μL,50 ng/μL DNA模板2.0 μL,用ddH2O補(bǔ)足至20.0 μL。將反應(yīng)體系置于T100 PCR儀上進(jìn)行PCR擴(kuò)增。擴(kuò)增程序:94 ℃預(yù)變性3 min;94 ℃ 30 s,退火溫度和時(shí)間見表2,72 ℃ 1 min,進(jìn)行30個(gè)循環(huán);72 ℃延伸10 min。
Hap-6A-P1和Hap-6A-P2為CAPS分子標(biāo)記,參照Su等(2011)的方法檢測TaGW2-6A基因的這2個(gè)CAPS分子標(biāo)記:用Hap-6A-P1引物進(jìn)行第一輪PCR擴(kuò)增,然后將第一輪擴(kuò)增的PCR產(chǎn)物適當(dāng)稀釋后作為模板,用Hap-6A-P2引物進(jìn)行第二輪PCR擴(kuò)增。第二輪PCR擴(kuò)增完畢后,用限制性內(nèi)切酶Taq I酶切第二輪PCR擴(kuò)增產(chǎn)物,PCR產(chǎn)物和酶切片段分別用毛細(xì)管電泳進(jìn)行分離。
1. 2. 4 毛細(xì)管電泳檢測 上述PCR擴(kuò)增產(chǎn)物使用Fragment AnalyzerTM毛細(xì)管電泳系統(tǒng)進(jìn)行檢測及分離。取2.0 μL PCR擴(kuò)增產(chǎn)物,加入22.0 μL Buffer TE。檢測程序:預(yù)運(yùn)行電壓和時(shí)間是6 kV和30 s;Marker進(jìn)樣電壓和時(shí)間為5 kV和10 s;樣品進(jìn)樣電壓和時(shí)間分別是5 kV和10 s;分離時(shí)電壓和時(shí)間是6 kV和45 min。
1. 3 統(tǒng)計(jì)分析
使用SPSS和PROSize 3.0進(jìn)行數(shù)據(jù)統(tǒng)計(jì)分析。粒重、粒長、粒寬等試驗(yàn)數(shù)據(jù)使用SC-G型多功能種子分析儀進(jìn)行獨(dú)立樣本檢驗(yàn)。擴(kuò)增結(jié)果使用PRO-Size 3.0進(jìn)行統(tǒng)計(jì)分析。
2 結(jié)果與分析
2. 1 小麥籽粒表型性狀測定結(jié)果
252份小麥材料的粒重平均為39.87 g,粒長平均為6.89 mm,粒寬平均為3.07 mm,其中以Y08選7的粒重最大(57.93 g),以中2012-14的粒重最小(21.55 g)(表3)。根據(jù)李立會和李秀全(2006)的小麥粒重劃分標(biāo)準(zhǔn),有56份材料屬于大粒種質(zhì)(>45.00 g),173份材料屬于中粒種質(zhì)(30.00~45.00 g),23份材料屬于小粒種質(zhì)(<30.00 g)。從變異系數(shù)來看,以粒重的變異系數(shù)較大,為16.8%,表明252份小麥品種的粒重遺傳變異較大。總體來說,252份小麥材料中以低粒重品種居多,說明對貴州小麥品種(系)進(jìn)行粒重鑒定及選育改良非常必要。
2. 2 TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位變異類型檢測結(jié)果
對252份小麥材料TaCwi-A1、TaSus2-2B和TaGW2-6A基因的等位變異類型檢測結(jié)果如表3所示。56份大粒種質(zhì)中僅有13份(ZY96-3、貴農(nóng)10-6、貴麥15、貴紫2號、貴農(nóng)10-6-16-2、貴農(nóng)10-6-16-6、貴農(nóng)10-6-16-7、貴農(nóng)15、貴農(nóng)15選-2-4、TP無芒-1、豐優(yōu)99號、貴麥15-1和惠光2-2-2)檢測到等位變異類型,這些種質(zhì)可用于貴州小麥粒重的遺傳改良和高粒重品種選育,173份中粒種質(zhì)中有24份檢測到等位變異類型,23份小粒種質(zhì)中均未檢測到等位變異類型。
2. 2. 1 TaCwi-A1等位變異類型檢測結(jié)果 根據(jù)Ma等(2012)的研究報(bào)道,TaCwi-A1基因存在2種等位變異類型,其中TaCwi-A1a變異類型用CWI22標(biāo)記引物能擴(kuò)增出402 bp的目的片段,TaCwi-A1b變異類型用CWI21標(biāo)記引物能擴(kuò)增出404 bp的目的片段。對CWI22和CWI21標(biāo)記引物的擴(kuò)增產(chǎn)物進(jìn)行毛細(xì)管電泳檢測,結(jié)果發(fā)現(xiàn),252份小麥材料中有8份材料(豐優(yōu)99、貴麥15、貴農(nóng)10-6、貴農(nóng)12-7、貴農(nóng)33、貴優(yōu)麥、興義98-2-4和ZY96-3)能擴(kuò)增出402 bp的目的片段(表3和圖1),說明這8份材料為TaCwi-A1基因的TaCwi-A1a變異類型,占供試材料總數(shù)的3.2%;有4份材料(TP無芒-1、貴農(nóng)10-6-16-6、貴紫2號和惠光2-2-2)能擴(kuò)增出404 bp的目的片段(表3和圖2),說明這4份材料為TaCwi-A1基因的TaCwi-A1b變異類型,占供試材料的1.6%。其他材料未檢測到到這2個(gè)目的片段。
2. 2. 2 TaSus2-2B基因等位變異類型檢測結(jié)果 根據(jù)Jiang等(2011)的研究報(bào)道,TaSus2-2B基因存在2種等位變異類型,其中,TaSus2-2BH變異類型用TaSus2-2BH標(biāo)記引物能擴(kuò)增出423 bp的目的片段,TaSus2-2BL變異類型用TaSus2-2BL標(biāo)記引物能擴(kuò)增出381 bp的目的片段。對TaSus2-1和TaSus2-2標(biāo)記引物的PCR擴(kuò)增產(chǎn)物進(jìn)行毛細(xì)管電泳檢測,252份小麥材料中有2份材料(貴農(nóng)10-19和貴紫4號)能擴(kuò)增出423 bp的目的片段(表3和圖3),說明這2份材料為TaSus2-2B基因的TaSus2-2BH變異類型,占供試材料總數(shù)的0.8%;有14份材料(163-15、TP-16、TP-3、貴麥15、貴農(nóng)08-9選10、貴農(nóng)10-3-16-9、貴農(nóng)10-4-16-20、貴農(nóng)12、貴農(nóng)15、貴農(nóng)15選-2-4、惠TP分-1、節(jié)四燕、張08-6特優(yōu)-2和張10選-4-1)能擴(kuò)增出381 bp的目的片段(表3和圖4),說明這14份材料為TaSus2-2B基因的TaSus2-2BL變異類型,占供試材料總數(shù)的5.6%。其他材料未檢測到這2個(gè)目的片段。
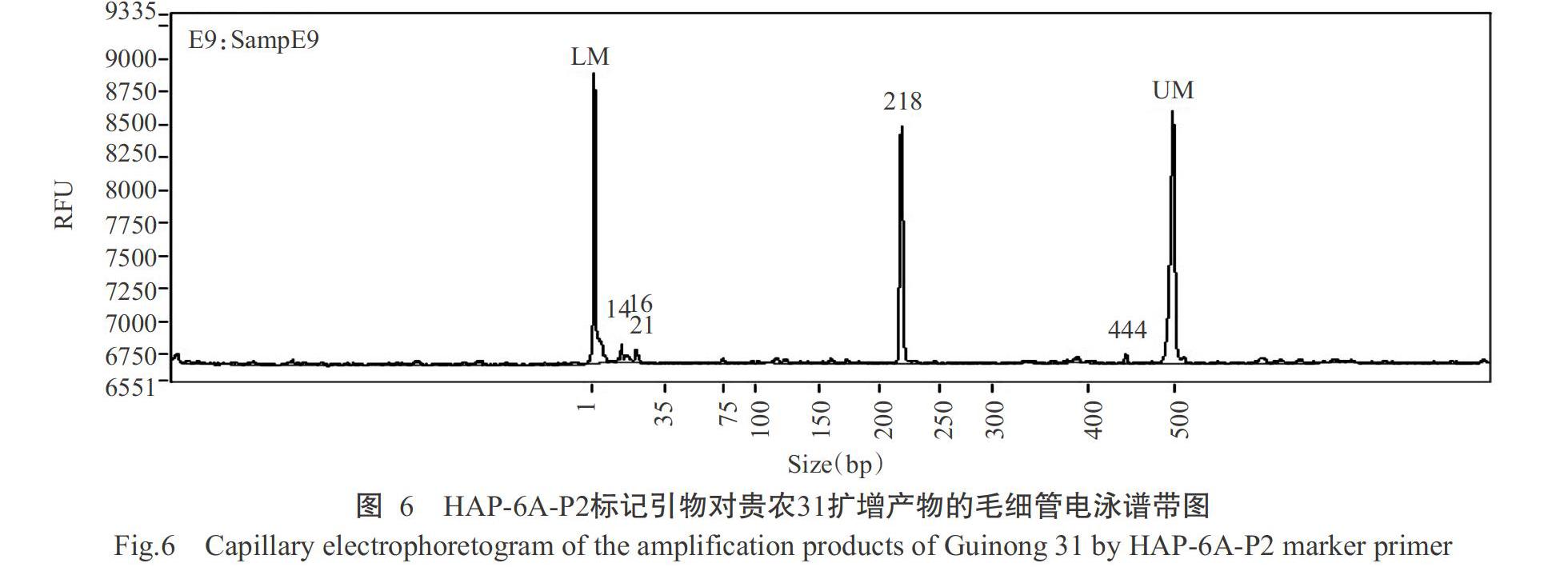
2. 2. 3 TaGW2-6A基因等位變異類型檢測結(jié)果 根據(jù)Su等(2011)的研究報(bào)道,TaGW2-6A基因利用相應(yīng)的CAPS分子標(biāo)記能檢測到Hap-6A-A和Hap-6A-G 2種等位變異類型。其中Hap-6A-A變異類型能檢測到167 bp的目的片段,Hap-6A-G變異類型能檢測到218 bp的目的片段。對Hap-6A-P1和Hap-6A-P2標(biāo)記引物的PCR擴(kuò)增產(chǎn)物進(jìn)行毛細(xì)管電泳檢測,結(jié)果發(fā)現(xiàn),252份材料中有4份材料(貴農(nóng)10-18、貴農(nóng)30-16、惠光2-2-2和黔09197)能檢測到167 bp的目的片段(表3和圖5),說明這4份材料為TaGW2-6A基因的Hap-6A-A變異類型,占供試材料總數(shù)的1.6%;有6份材料(貴農(nóng)10-6-16-9、貴農(nóng)10-6-16-2、貴農(nóng)10-6-16-7、貴農(nóng)31、貴農(nóng)優(yōu)麥2號和惠紫1號)能檢測到218 bp的目的片段(表3和圖6),說明這6份材料為TaGW2-6A基因的Hap-6A-G變異類型,占供試材料總數(shù)的2.4%。其他材料未檢測到這2個(gè)目的片段。
2. 3 等位變異類型統(tǒng)計(jì)及籽粒表型差異顯著性分析結(jié)果
TaCwi-A1、TaSus2-2B和TaGW2-6A基因的等位變異類型統(tǒng)計(jì)結(jié)果如表4所示。252份小麥材料中,含有TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位變異的材料有37份,占供試材料總數(shù)的14.7%,其中TaSus2-2B基因等位變異類型材料16份,占供試材料總數(shù)的6.4%,TaCwi-A1基因等位變異類型材料有12份,占供試材料總數(shù)的4.8%,TaGW2-6A基因等位變異類型材料有10份,占供試材料總數(shù)的4.0%;等位變異組合類型僅有1份材料(惠光2-2-2),為TaCwi-A1b/HAP-6A-A,占供試材料總數(shù)的0.4%;有43份粒重高于45.00 g的材料均未檢測到TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位變異。平均粒重最重的變異類型為TaCwi-A1b,為47.60 g,其次是HAP-6A-G變異類型,平均粒重為44.04 g。
由表4還可知,TaCwi-A1b變異類型材料的平均粒重(47.60 g)顯著高于TaCwi-A1a(41.95 g)變異類型(P<0.05,下同),Hap-6A-G變異類型材料的平均粒重(44.04 g)顯著高于Hap-6A-A變異類型(40.77 g),說明TaCwi-A1b和Hap-6A-G變異類型均能明顯增加小麥的粒重。TaSus2-2BH變異類型材料的平均粒重(43.47 g)也高于TaSus2-2BL變異類型(41.75 g),但差異不顯著(P>0.05,下同)。在TaCwi-A1、TaSus2-2B和TaGW2-6A基因的等位變異類型中,僅TaSus2-2BH變異類型材料的平均粒長(7.21 mm)顯著高于TaSus2-2BL變異類型材料(7.05 mm),其他變異類型材料的平均粒長和平均粒寬差異均不顯著。
3 討論
本研究對貴州252份種質(zhì)材料中粒重相關(guān)基因(TaCwi-A1、TaSus2-2B和TaGW2-6A)等位變異的類型及分布頻率進(jìn)行研究,結(jié)果發(fā)現(xiàn)14.7%的供試材料含有TaCwi-A1a、TaCwi-A1b、TaSus2-2BH、TaSus2-2BL、HAP-6A-A和HAP-6A-G 6種變異類型,其中以TaSus2-2BL變異類型材料最多,占供試材料總數(shù)的5.6%,以TaSus2-2BH變異類型材料最少,占供試材料總數(shù)的0.8%,且從所有材料中只檢測到1份材料為等位變異組合類型TaCwi-A1b/HAP-6A-A,僅占供試材料總數(shù)的0.4%。與前人研究結(jié)果相比,本研究中各基因等位變異類型所占比例均較低。如劉永偉等(2017b)研究發(fā)現(xiàn),黃淮麥區(qū)小麥中TaCwi-A1a變異類型所占比列為65.03%;李娜等(2018)研究顯示,在64個(gè)青海小麥品種中,62.5%的材料為HAP-6A-A變異類型;時(shí)佳等(2018)研究發(fā)現(xiàn),219份新疆小麥種質(zhì)材料中,40.18%的材料為TaSus2-B2A變異類型;仝靖洋等(2018)從262份供試材料中檢測出8種等位變異組合類型,遠(yuǎn)多于本研究檢測出的等位變異組合類型。存在較大差異的原因除了供試材料不同外,很可能是本研究中的供試材料主要是地方品種,高代相近品系較多,整體遺傳多樣性較低,導(dǎo)致檢測到的粒重基因等位變異較少。這反映出貴州小麥高粒重品種的選育工作開展不夠,今后應(yīng)重視小麥粒重基因等位變異綜合效應(yīng)研究。
本研究還發(fā)現(xiàn),TaSus2-2BH變異類型材料的平均粒重高于TaSus2-2BL變異類型材料,與Jiang等(2011)、Ma等(2012)的研究結(jié)果一致;TaCwi-A1b變異類型材料的平均粒重顯著高于TaCwi-A1a變異類型材料,與Jiang等(2011)、Ma等(2012)的研究結(jié)果存在差異;Hap-6A-G變異類型材料的平均粒重高于Hap-6A-A變異類型材料,與李娜等(2018)的研究結(jié)果相同,而與Su等(2011)的研究結(jié)果不同;在TaCwi-A1、TaSus2-2B和TaGW2-6A基因的等位變異類型中,僅TaSus2-2BH變異類型材料的平均粒長顯著高于TaSus2-2BL變異類型材料,其他變異類型材料的平均粒長和平均粒寬差異均不顯著,與仝靖洋等(2018)的研究結(jié)果相似,而與劉永偉等(2017b)的結(jié)果不同。上述與前人研究結(jié)果不一致的主要原因是供試材料中檢測到的各等位變異類型材料份數(shù)較少,且測定的粒重僅為1年數(shù)據(jù),不同等位變異類型對不同材料粒重的貢獻(xiàn)率僅作為參考,今后應(yīng)擴(kuò)大樣本量,進(jìn)一步開展不同變異類型的綜合效應(yīng)研究,用于小麥高粒重品種分子輔助選擇育種。此外,本研究發(fā)現(xiàn)43份粒重高于45.00 g的材料均未檢測到TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位變異,針對這些材料可開展多年多點(diǎn)試驗(yàn)鑒定是否含有其他粒重相關(guān)的新基因,將這些新基因應(yīng)用于小麥高粒重品種遺傳改良。
據(jù)報(bào)道,貴州小麥育種在“十一五”期間一直徘徊不前,如果還是在現(xiàn)有的遺傳背景下繼續(xù)進(jìn)行品種選育,則很難有重大突破(英敏和余虎,2012),應(yīng)通過拓寬高粒重的親本遺傳背景來提高小麥品種的產(chǎn)量潛力(何昀昆等,2008)。大量研究證實(shí),TaCwi-A1a、TaSus2-B2H等均為優(yōu)異等位變異類型,其對粒重有顯著增效作用,今后可加大聚合多個(gè)優(yōu)異等位變異組合類型的種質(zhì)資源收集、鑒定和篩選研究,提高其在貴州小麥種質(zhì)資源中的比例,開展小麥高粒重品種的遺傳改良,從而有效提高貴州小麥產(chǎn)量整體水平。
4 結(jié)論
從貴州小麥品種(系)檢測到的粒重基因等位變異整體較少,表明貴州小麥種質(zhì)遺傳多樣性較低,高粒重品種的選育工作開展不夠,今后應(yīng)重視小麥粒重基因等位變異綜合效應(yīng)研究。鑒定出含有TaCwi-A1、TaSus2-2B和TaGW2-6A基因等位變異類型、粒重>45.00 g的品種(系)13份,可應(yīng)用于貴州小麥粒重的遺傳改良和高粒重品種選育。
參考文獻(xiàn):
韓利明,楊芳萍,夏先春,閻俊,張勇,曲延英,王忠偉,何中虎. 2011. 株高、粒重及抗病相關(guān)基因在不同國家小麥品種中的分布[J]. 麥類作物學(xué)報(bào),31(5):824-831. [Han L M,Yang F P,Xia X C,Yan J,Zhang Y,Qu Y Y,Wang Z W,He Z H. 2011. Distribution of plant height,grain weight and disease-resistance related genes in wheat varie-ties in different countries[J]. Journal of Triticeae Crops,31(5):824-831.]
何昀昆,田曉琴,何慶才. 2008. 貴州省小麥育種研究與分析[J]. 種子,(10):95-97. [He Y K,Tian X Q,He Q C. 2008. Research and analysis of wheat breedingin Guizhou[J]. Seed,(10):95-97.]
李立會,李秀全. 2006. 小麥種質(zhì)資源描述規(guī)范和數(shù)據(jù)標(biāo)準(zhǔn)[M]. 北京:中國農(nóng)業(yè)出版社. [Li L H,Li X Q. 2006. Descriptors and data standard for wheat[M]. Beijing:China Agricultural Press.]
李娜,魏樂,曹東,陳文杰,張波,張懷剛,劉寶龍. 2018. 來源于人工合成小麥的TaGW2-6A等位變異類型的鑒定及分析[J]. 分子植物育種,16(2):518-525. [Li N,Wei L,Cao D,Chen W J,Zhang B,Zhang H G,Liu B L. 2018. Identification and analysis of TaGW2-6A allelic variation from synthetic wheat[J]. Molecular Plant Breeding,16(2):518-525.]
劉永偉,周碩,王雪征,孫果忠,董福雙,柴建芳,李春杰,趙和,王海波. 2017a. 小麥粒重相關(guān)分子標(biāo)記Xgwm46等位變異的鑒定與評價(jià)[J]. 麥類作物學(xué)報(bào),37(1):7-15. [Liu Y W,Zhou S,Wang X Z ,Sun G Z,Dong F S,Chai J F,Li C J,Zhao H,Wang H B. 2017a. Identification and evaluation of molecular marker Xgwm46 allelic variations associated with kernel weight in wheat[J]. Journal of Triticeae Crops,37(1):7-15.]
劉永偉,周碩,王雪征,孫果忠,朱金永,韓秋芬,李春杰,趙和,王海波. 2017b. 粒重基因TaCwi-A1等位變異在黃淮麥區(qū)小麥品種(系)中的分布及功能分析[J]. 華北農(nóng)學(xué)報(bào),32(2):131-137. [Liu Y W,Zhou S,Wang X Z,Sun G Z,Zhu J Y,Han Q F,Li C J,Zhao H,Wang H B. 2017b. Functional analysis and distribution of allelic varia-tions of TaCwi-A1 gene related to kernel weight in Ye-llow and Huai river valleys facultative wheat zone[J]. Acta Agriculturae Boreali-Sinica,32(2):131-137.]
時(shí)佳,白璐,任毅,穆培源,梁曉東,瑪依拉,耿洪偉. 2018. 新疆小麥TaGW2-6A、TaCwi-A1、TaSus2-2B等位變異對粒重的影響及應(yīng)用[J]. 分子植物育種,16(3):848-858. [Shi J,Bai L,Ren Y,Mu P Y,Liang X D,Ma Y L,Di H W. 2018. Effects and application of allelic variation of TaGW2-6A,TaCwi-A1 and TaSus2-2B on grain weight of Xinjiang wheat[J]. Molecular Plant Breeding,16(3):848-858.]
司文潔,吳林楠,郭利建,周夢蝶,劉香利,馬猛,趙惠賢. 2019. 小麥粒重相關(guān)基因TaCYP78A5功能標(biāo)記開發(fā)及驗(yàn)證[J]. 作物學(xué)報(bào),45(12):1905-1911. [Si W J,Wu L N,Guo L J,Zhou M D,Liu X L,Ma M,Zhao H X. 2019. Development and validation of the functional marker of grain weight-related gene TaCYP78A5 in wheat(Triticum aestivum L.)[J]. Acta Agronomica Sinica,45(12):1905-1911.]
仝靖洋,李少鵬,劉勝杰,張琳雪,梁園園,張哲,聶小軍,李學(xué)軍,王中華,高欣. 2018. 小麥粒重基因等位變異的高通量分子檢測及組合分析[J]. 麥類作物學(xué)報(bào),38(11):1300-1308. [Tong J Y,Li S P,Liu S J,Zhang L X,Liang Y Y,Zhang Z,Nie X J,Li X J,Wang Z H,Gao X. 2018.High-throughput molecular detection and analysis of allelic variation combinations related to grain weight gene of wheat(Triticum aestivum L.)[J]. Journal of Triticeae Crops,38(11):1300-1308.]
相吉山,穆培源,桑偉,聶迎彬,徐紅軍,莊麗,崔鳳娟,韓新年,鄒波. 2014. 小麥粒重基因TaCwi-A1功能標(biāo)記CWI22、CWI21的驗(yàn)證及應(yīng)用[J]. 中國農(nóng)業(yè)科學(xué),47(13):2671-2709. [Xiang J S,Mu P Y,Sang W,Nie Y B,Xu H J,Zhuang L,Cui F J,Han X N,Zou B. 2014. Validation and application of function markers CWI22 and CWI21 of Tacwi-A1 gene related to wheat kernel weight[J]. Scientia Agricultura Sinica,47(13):2671-2709.]
英敏,余虎. 2012. “十一五”貴州小麥區(qū)試結(jié)果分析與評價(jià)[J]. 種子,(5):101-102. [Ying M,Yu H. 2012. Analysis and evaluation on wheat regional trial in Guizhou Pro-vince during ‘11 th five-year plan[J]. Seed,(5):101-102.]
Dong Y M,Jun Y,Zhong H H,Ling W,Xian C X. 2012. Characterization of a cell wall invertase gene TaCwi-A1 on common wheat chromosome 2A and development of functional markers[J]. Molecular Breeding,29:43-52.
Hunter M C,Smith R G,Schipanski M E,Atwood L W,Mortensen D A. 2017. Agriculture in 2050:Recalibrating targets for sustainable intensification[J]. Bioscience,67(4):385-390.
Jiang Q Y,Hou J,Hao C Y,Wang L F,Ge H M,Dong Y S,Zhang X Y. 2011. The wheat(T. aestivum) sucrose synthase 2 gene(TaSus2) active in endosperm development is associated with yield traits[J]. Functional & Integrative Genomics,11:49-61.
Ma D Y,Yan J,He Z H,Wu L,Xia X C. 2012. Characterization of a cell wall invertase gene TaCwi-A1 on common wheat chromosome 2A and development of functional markers[J]. Molecular Breeding,29:43-52.
Su Z Q,Hao C Y,Wang L F,Dong Y C,Zhang X Y. 2011. Identification and development of a functional marker of TaGW2 associated with grain weight in bread wheat(Triti-cum aestivum L.)[J]. Theoretical & Applied Genetics,122(1):211-223.
Tian J C,Deng Z Y,Hu R B,Wang Y X. 2006. Yield components of super wheat cultivars with different types and the path coeffificient analysis on grain yield[J]. Acta Agro-nomica Sinica,32(11):1699-1705.
(責(zé)任編輯 陳 燕)
