乙酰乳酸合成酶抗性突變的分子動力學(xué)模擬
2020-12-11 09:10:33趙奇鄧培淵郭運宏陳麗培羅青
南方農(nóng)業(yè)學(xué)報 2020年9期
趙奇 鄧培淵 郭運宏 陳麗培 羅青

摘要:【目的】研究乙酰乳酸合成酶(ALS)及Pro197Ser(P197S)突變體與抑制劑苯磺隆(TBM)的分子結(jié)合模式,闡明其分子作用機制,為基于ALS的新型除草劑研發(fā)和雜草防控等提供參考。【方法】運用AutoDock 4.2進行ALS和P197S與TBM多構(gòu)象對接,采用YASARA平臺進行分子動力學(xué)模擬,利用MMPBSA.py模塊計算ALS_TBM和P197S_TBM體系復(fù)合物的結(jié)合自由能,并以Amber 12分析關(guān)鍵氨基酸、結(jié)合模式和相互作用力。【結(jié)果】P197S_TBM突變體系接觸氨基酸殘基數(shù)量增多,接觸位點偏集中于C端,氫鍵數(shù)量更多、鍵長更短;P197S_TBM體系的均方根偏差(RMSD)較低,250~320位氨基酸殘基區(qū)域和350~430位氨基酸殘基區(qū)域均方根漲落(RMSF)下降明顯;P197S_TBM體系的結(jié)合P197S自由能更低,親和力貢獻大的氨基酸殘基數(shù)量增加。【結(jié)論】P197S突變引起ALS與TBM結(jié)合位點和結(jié)合模式發(fā)生變化,接觸氨基酸殘基和氫鍵增多,促結(jié)合能量值增加,突變體系更穩(wěn)定。417~423位氨基酸殘基在ALS與TBM結(jié)合過程中是較集中貢獻能量的氨基酸團。
關(guān)鍵詞: 乙酰乳酸合成酶;定點突變;苯磺隆;分子動力學(xué)模擬
中圖分類號: S482.4? ? ? ? ? ? ? ? ? ? ? ? ? ? 文獻標(biāo)志碼: A 文章編號:2095-1191(2020)09-2167-07
Molecular dynamics simulation on resistance mutation of acetolactate synthase
ZHAO Qi1, DENG Pei-yuan1, GUO Yun-hong2, CHEN Li-pei1, LUO Qing1
(1School of Life Science, Zhengzhou Normal University, Zhengzhou? 450044, China; 2Department of Architectural Engineering, Zhengzhou Railway Vocational & Technical College, Zhengzhou? 450044, China)
Abstract:【Objective】Molecular dynamics model of acetolactate synthase(ALS) and ALS mutant Pro197Ser(P197S)with inhibitor tribenuron-methy(TBM) were studied to illustrate the molecular mechanism of their recognition and provide reference for research and development of new herbicide as well as the prevention and control of weeds based on ALS. 【Method】AutoDock 4.2 was used to dock ALS and P197S with TBM, molecular dynamics simulation was conduc-ted by YASARA platform,binding free energy of complexof ALS_TBM and P197S_TBM systems was calculated with MMPBSA. py, key amino acids, combination patterns and interactions were analyzed with Amber12 package. 【Result】In P197S-TBM mutation system, the number of contact residues increased,the binding sites were close to the C-terminal region,and hydrogen bonds also increased, the length of hydrogen bond length was shorter. Root mean square deviation(RMSD)in P197S_TBM system was low, root mean square fluctuation(RMSF) of 250-320 and 350-430 amino acid residues regions decreased obviously. The binding P197S free energy in P197S_TBM system was lower, while the number of amino acid residues with large affinity increased. 【Conclusion】The P197S mutation causes changes in the binding sites and binding patterns of ALS and TBM, increases contact amino acid residues, hydrogen bonds and binding energy, and mutation system is more stable. 417-423 amino acid residues is the amino acid cluster that contributes during the binding of ALS and TBM.
Key words: acetolactate synthase; site-directed mutation; tribenuron-methy; molecular dynamics simulation
Foundation item: Youth Program of Humanities and Social Sciences of Ministry of Education(19YJCZH263); Key Scientific Research Project of Colleges of Henan Department of Education(19A180033)
0 引言
【研究意義】乙酰乳酸合成酶(ALS)廣泛存在于植物和其他生物體內(nèi),ALS抑制劑抑制植物體內(nèi)合成3種支鏈氨基酸,影響蛋白和DNA的合成,從而導(dǎo)致植物細胞有絲分裂不能正常完成,致使植物停止生長直至死亡(黃新發(fā),2003;付三雄等,2019)。以ALS為靶標(biāo)的除草劑能高效殺死雜草,對人類和動物安全無害。自1987年發(fā)現(xiàn)第一例雜草對ALS除草劑產(chǎn)生抗性以來,截至1997年共報道了159種雜草生物對ALS除草劑產(chǎn)生抗性(鄧維,2017)。Yu和Powles(2014)研究指出,靶標(biāo)位點突變和非靶標(biāo)抗性中的代謝抗性是雜草產(chǎn)生ALS除草劑抗性的機制。在我國,隨著以ALS為靶標(biāo)的苯磺隆抑制劑類除草劑連年使用,抗ALS抑制劑類除草劑的問題日益惡化,因此,開發(fā)新型ALS靶標(biāo)抑制劑顯得尤為重要,而了解ALS與除草抑制劑的結(jié)合模式是延緩雜草抗性種群發(fā)展(張樂樂等,2016)、新型除草劑研發(fā)及雜草防控的基礎(chǔ)工作。【前人研究進展】針對ALS,前人大多在ALS基因分子水平上進行功能研究,如克隆植物ALS基因、比較ALS基因序列,以及探尋哪些氨基酸改變會引起雜草產(chǎn)生ALS抑制劑類除草劑抗性。Yu等(2012)研究認為磺酰脲類除草劑主要在Pro197位點發(fā)生突變,有14種不同的突變類型,其中突變頻率最高的是Pro突變?yōu)镾er;黃新發(fā)(2003)的研究中亦有同樣結(jié)論。Panozzo等(2013)研究表明,雜草抗ALS除草劑的突變位點為Ala122、Pro197、Ala205、Asp376、Arg377、Trp574、Ser653和Gly654,共有29種氨基酸取代類型,其中Pro197突變位點是最常見的突變類型。張樂樂等(2016)比較了苯磺隆抗性和敏感薺菜種群ALS基因序列差異,得出抗性差異與197和574 2個位點突變有關(guān)。鄧維(2017)在雜草播娘蒿ALS基因突變多樣性的基礎(chǔ)上開展突變對ALS功能的影響研究,結(jié)果表明Pro197Ser在抗性突變種群的比例為28.6%,在抗性群體中位居第二。此類ALS突變研究結(jié)果表明,Pro197Leu/Ser/Arg/Gln/Ala突變已在黑麥草、地膚、野蘿卜、藺草和牛繁縷上對磺酰脲類除草劑產(chǎn)生抗性(Guttieri et al.,1995;Yu et al.,2008;金濤,2012;辛潔等,2019)。以上研究側(cè)重于探究突變產(chǎn)生抗性的基因基礎(chǔ),而少有進行突變基因與抑制劑除草劑的作用模式研究。近期,楊冬臣等(2019)開展了ALS與抑制劑除草劑分子結(jié)合模式的研究,通過同源建模方法構(gòu)建了ALS的三維結(jié)構(gòu),并使用分子對接和分子動力學(xué)方法預(yù)測反之莧ALS與煙嘧磺隆分子的結(jié)合模式,認為影響二者分子結(jié)合穩(wěn)定性的因素有分子間氫鍵、疏水作用及通道形態(tài)等,推測雜草ALS產(chǎn)生抗性的突變位點有Val196、Met200、Phe206和Lys256,但該研究未進行突變前后與抑制劑類除草劑分子結(jié)合模式的對比分析。【本研究切入點】前人對Pro197突變的研究主要集中在了解除草劑抗性變化方面。Pro197突變?yōu)榻z氨酸(Pro197Ser)是高頻突變的一種,此前雖有Pro197Ser與煙嘧磺隆分子抑制劑類型的分子模擬研究,但鮮見與苯磺隆類型抑制劑的分子模擬,更缺少突變前后ALS與除草劑分子結(jié)合模式變化的相關(guān)研究。【擬解決的關(guān)鍵問題】選用Pro197Ser(P197S)高頻突變,采用分子對接和分子動力學(xué)模擬探討野生型ALS及P197S突變體ALS與磺酰脲類抑制劑——苯磺隆(TBM)的結(jié)合模式,展示靜態(tài)對接結(jié)果和動態(tài)分子結(jié)合過程特征,以期闡明P197S突變產(chǎn)生TBM抗性的分子機制,同時為基于ALS的新型抑制劑研發(fā)提供參考。
1 材料與方法
1. 1 體系搭建
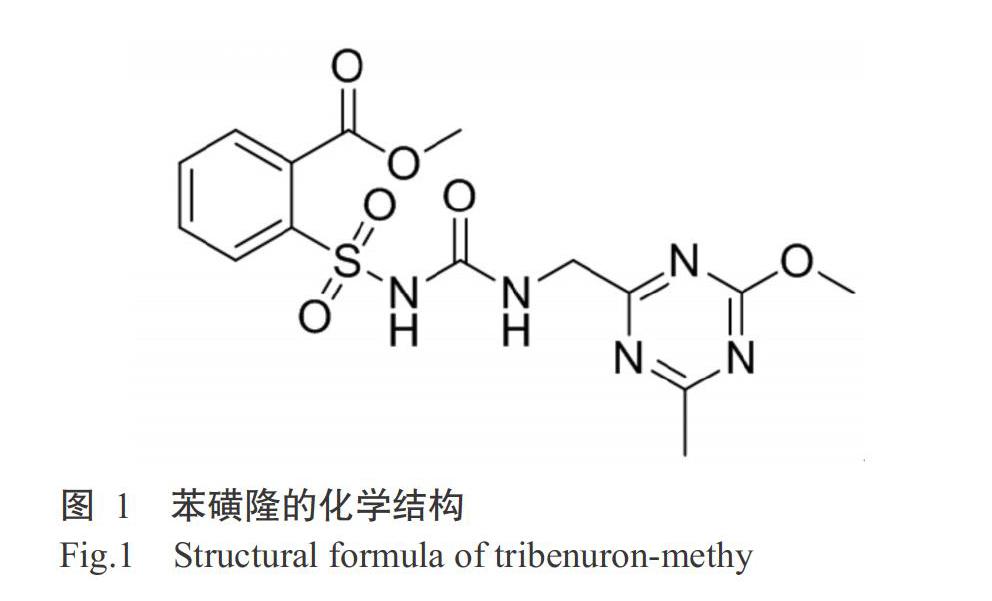
下載擬南芥ALS晶體結(jié)構(gòu)為參考(PDB代碼:1YHY),導(dǎo)入YASARA分子模擬軟件(成都分迪科技有限公司,版本17.1.28)優(yōu)化結(jié)構(gòu),去除水分子、加氫和添加缺失原子處理后作為野生型擬南芥ALS核心結(jié)構(gòu),使用最陡下降法和共軛梯度法進行能量優(yōu)化處理,獲得野生型ALS,利用Pymol軟件突變P197S。運用ChemBioDraw(ChemBioOffice 10.0)模塊構(gòu)建苯磺隆分子結(jié)構(gòu)(圖1),使用ChemBio 3D構(gòu)建立體空間結(jié)構(gòu)和能量優(yōu)化,優(yōu)化后的結(jié)構(gòu)作為初始配體結(jié)構(gòu),采用TBM表示。
1. 2 多構(gòu)象分子對接
為了使對接結(jié)果與真實情形更吻合,本研究采用多構(gòu)象對接方法,首先對WT和P197S 2個受體進行2020 ps的MD模擬,每隔200 ps取構(gòu)象與苯磺隆進行對接,均完成10次對接。使用AutoDock 4.2完成多構(gòu)象對接,拉馬克遺傳算法全局搜索與局部搜索2種方式結(jié)合運用,對100個對接結(jié)果進行全局優(yōu)化;選取對接能量最低的10個構(gòu)象分析確定結(jié)合差異不明顯,將最低對接能構(gòu)象作為對接最終結(jié)果。WT_TBM表示W(wǎng)T與TBM的對接結(jié)果,P197S_TBM表示P197S與TBM的對接結(jié)果。
1. 3 分子動力學(xué)模擬
對WT、P197S、WT_TBM和P197S_TBM進行MD模擬。運用YASARA分子模擬平臺,采用Amber 12力場,將模擬對象溶解于TIP3P顯性水模型,保持體系電中性。使用條件:pH 7.4、300 K、NVT正則系綜,通過郎格萬方法控制體系溫度。MD模擬40 ns,積分步長2.5 fs,采樣間隔10 ps。
1. 4 結(jié)合自由能算法
采用Amber 12中的MMPBSA.py模塊計算WT_TBM和P197S_TBM體系結(jié)合自由能,選取動力學(xué)模擬穩(wěn)定體系的構(gòu)象用于分析(鄧培淵等,2008;趙奇等,2018)。公式如下:
ΔGbind =ΔGcomplex-ΔGreceptor-ΔGligand
ΔG =ΔEgas+ΔGsolve-TΔS
ΔEgas=ΔEele+ΔEvdw
ΔGsolve =ΔGpb+ΔGnopolar
式中,ΔGbind為結(jié)合自由能,ΔGcomplex為復(fù)合物的結(jié)合自由能,ΔGreceptor為受體結(jié)合自由能,ΔGligand為配體的結(jié)合自由能,ΔEvdw為范德華力能,ΔEele為靜電作用力能;ΔGsolve為溶劑化能,包括極性溶劑化貢獻能(ΔGpb)和非極性溶劑化貢獻能(ΔGnopolar);ΔEgas為氣相內(nèi)能;TΔS為體系熵變值,根據(jù)本研究目的,熵變值忽略為0。
1. 5 能量分解
基于單個氨基酸能量分解的方法,利用Amber 12描述PxylPBP2和Z9-14:Ac活性口袋關(guān)鍵氨基酸。氨基酸殘基能量貢獻通過公式換算為4個能量項,公式如下:
ΔG =ΔEele+ΔEvdw+ΔGpb+ΔGnopolar
2 結(jié)果與分析
2. 1 結(jié)合模式預(yù)測結(jié)果
設(shè)置Grid Box包含模型,采用126×126×126對接體系格點,0.0375 nm格點間距,多構(gòu)象分子對接后進行聚類分析,依據(jù)對接條件設(shè)計進行25次對接,并對對接結(jié)果進行聚類分析,且每個聚類至少具有5.0 ?的均方根偏差(RMSD)差異。WT_TBM體系聚類得到18個不同的復(fù)雜構(gòu)象,P197S_TBM體系聚類得到13個不同的復(fù)雜構(gòu)象。對接結(jié)果見表1,表中列出前3類結(jié)果。
選擇對接能最強的結(jié)合模式作為WT_TBM和P197S_TBM最初的結(jié)合模式(圖1和圖2)。由對接結(jié)果可知,WT_TBM體系中接觸的氨基酸殘基主要有Lys220、His221、Leu241、Gly245、Leu246、Leu273、Gly275、Y276、Arg279、Met280、Pro281、Ile396和Asp397;P197S_TBM體系中接觸的氨基酸殘基主要有Phe458、Ala461、Gln566、His567、Glu594、Asp595、Ile597、Thr616、Ile640、Cys641、Pro642、His643、Gln644和Arg667。與WT_TBM相比,P197S_TBM體系接觸的氨基酸殘基數(shù)量增多,且接觸位點偏集中于C端。
對比圖1和圖2可知,WT突變?yōu)镻197S后,與TBM的結(jié)合模式發(fā)生了明顯變化,更貼近loop功能區(qū)(圖2-B中圈出部分)。在WT_TBM體系中Arg246與復(fù)合物N端、O端分別形成2.46 ?和2.28 ?的氫鍵,與Lys220形成3.3 ?的π鍵。在P197S_TBM體系中His643與復(fù)合物O端形成2個長度分別為2.09 ?和2.36 ?的氫鍵,Gln644、Arg667與O端各形成1個2.0 ?和2.0 ?的氫鍵。二者相比,突變后氫鍵數(shù)量更多、鍵長更短,這些氫鍵對復(fù)合物穩(wěn)定性起著非常重要的作用。
2. 2 MD軌跡比較分析結(jié)果
通過分子動力學(xué)模擬,從RMSD和氨基酸殘基均方根漲落(RMSF)能進一步了解對接結(jié)果和對接復(fù)合物的穩(wěn)定性。MD軌跡結(jié)果(圖3)表明,WT_TBM和P197S_TBM體系總勢能在150 ps后趨于穩(wěn)定,650 ps后P197S_TBM體系的RMSD達收斂平衡;WT_TBM體系在10000 ps前達到一個相對的收斂平衡,之后發(fā)生較大幅度的提升,且從17000 ps逐步趨于平穩(wěn)到收斂平衡。在整個模擬時間段內(nèi),WT_TBM體系的RMSD均比P197S_TBM體系的高,表明P197S_TBM體系運動性比WT_TBM體系低且更穩(wěn)定。對P197S_TBM體系和WT_TBM體系的氫鍵分析結(jié)果顯示,P197S_TBM體系氨基酸殘基His643、Gln644、Arg667與O端形成的4個氫鍵有利于降低P197S_TBM體系的柔性,與P197S_TBM體系RMSD分析結(jié)論相符。
RMSF能反應(yīng)對接復(fù)合物的柔性和局部運動特征,數(shù)值越大,表明作用過程中柔性越強。圖4展示了RMSF的變化特征,相對于WT_TBM體系,P197S_TBM體系中有215個氨基酸殘基RMSF下降,且在250~320位氨基酸殘基區(qū)域和350~430位氨基酸殘基區(qū)域較集中,其他區(qū)域較分散。表明突變后中部區(qū)域的柔性下降較集中,同時形成氫鍵的644位點RMSF下降,有利于loop功能區(qū)的柔性下降。
2. 3 2個分子動力學(xué)體系的疊合分析結(jié)果
通過YASARA進行疊合分析,結(jié)果(圖5)顯示,突變前后分子結(jié)構(gòu)有2處(圖5-A)發(fā)生變化:第1處為665~668位氨基酸區(qū)域(圖5-B),該處區(qū)域突變后結(jié)構(gòu)朝向相反方向;第2處為273~278位氨基酸區(qū)域(圖5-C),該處表現(xiàn)疊合結(jié)構(gòu)間距離較遠。
2. 4 結(jié)合自由能分析結(jié)果
采用MM-PBSA計算WT_TBM體系和P197S_ TBM體系的ΔGbind。由表2可知,WT_TBM體系的結(jié)合自由能為-4.18 kcal/mol,突變后P197S_TBM體系的結(jié)合自由能為-8.70 kcal/mol,ΔGbind為負值,說明2種復(fù)合物均自發(fā)形成鍵合作用。比較二者結(jié)合過程中釋放的自由能可知突變體系中的結(jié)合力更強,說明P197S位點突變促進了ALS與苯磺隆結(jié)合。從2個體系的能量組成來看,WT_TBM體系結(jié)合的主要驅(qū)動力來自靜電能(ΔEele=-134.68 kcal/mol)、范德華力能(ΔEvdw=-51.61 kcal/mol)和非極性溶劑能(ΔGnopolar=-33.27 kcal/mol),而溶劑化能和極性溶劑能不利于野生體系結(jié)合的力量。突變體系的促結(jié)合能量與抑制結(jié)合能量類型均相同,促結(jié)合能量項數(shù)值均比野生型能量分項數(shù)值略大,抑制結(jié)合能量略小,故表現(xiàn)為數(shù)值更低的總結(jié)合自由能。
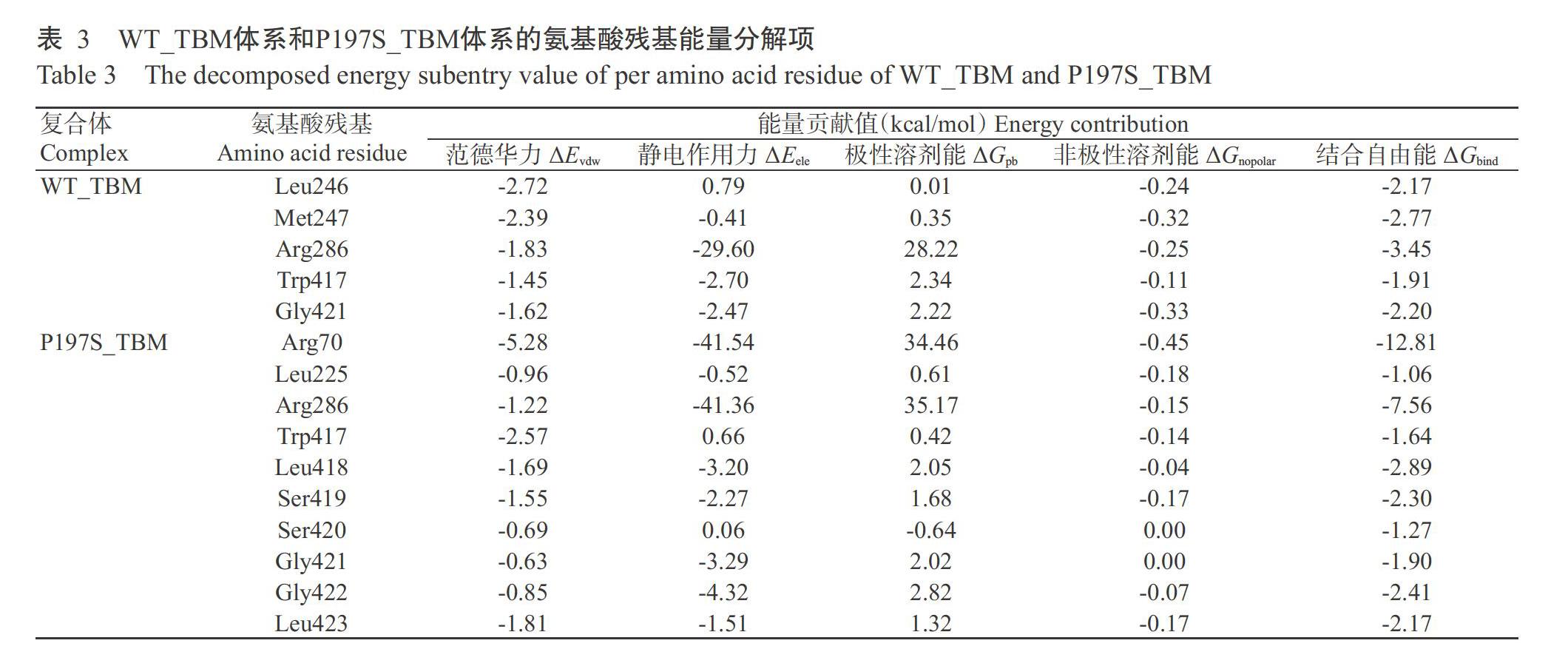
2. 5 單個氨基酸殘基的能量分解結(jié)果
進一步通過計算單個氨基酸能量以明確各氨基酸殘基對分子親和力的貢獻。表3列出能量貢獻值接近或大于1.00 kcal/mol的氨基酸殘基。WT_ TBM體系中能量貢獻突出的氨基酸殘基由大到小為:Arg286>Met247>Gly421>Leu246>Trp417,其中Arg286的能量貢獻分項最大的是靜電作用力(ΔEele=-29.60 kcal/mol)。P197S_TBM體系中能量貢獻突出的氨基酸殘基由大到小為:Arg70>Arg286>Leu418>Gly422>Ser419>Leu423>Gly421>Trp417>Ser420>Leu225,其中Arg70的能量貢獻分項最大的是靜電作用力(ΔEele=-41.54 kcal/mol),對復(fù)合物體系能量貢獻位列第一,能量貢獻占表3所列氨基酸總能量的55.2%。突變后,親和力貢獻大的氨基酸數(shù)量由5個增至10個。值得一提的是,WT_TBM體系中Arg70的自由能貢獻低于1.00 kcal/mol,僅為-0.76 kcal/mol,突變后增加至-12.81 kcal/mol,主要體現(xiàn)為靜電作用能量的大幅增加,由原來的0.79 kcal/mol轉(zhuǎn)變?yōu)?41.54 kcal/mol,由不利于結(jié)合的作用轉(zhuǎn)變?yōu)榇龠M結(jié)合。對比2個體系還可看出,突變后417~423位氨基酸變成較集中貢獻能量的氨基酸團。
3 討論
ALS由催化亞基和調(diào)節(jié)亞基組成,催化亞基是酶的活性中心,調(diào)節(jié)亞基本身無催化活性,但能反饋抑制調(diào)節(jié)催化亞基的活性(Duggleby et al.,2008)。眾多學(xué)者對催化亞基與ALS不同抑制劑結(jié)合進行了深入研究,結(jié)果表明,ALS的催化部位在ALS形成通道內(nèi)部且遠離通道口部位,作為抑制劑的除草劑不能直接與催化位點結(jié)合,而是與橫跨通道入口附近的ALS除草劑結(jié)合區(qū)域結(jié)合(McCourt et al.,2006)。該橫跨區(qū)域共有18個氨基酸殘基,Pro197是其中之一,這也是本研究選擇該突變位點的依據(jù)。除草劑與ALS的結(jié)合區(qū)域遠離催化部位,才會存在很多(多達十幾個)阻礙除草劑結(jié)合的突變位點。已有的研究也表明多個氨基酸突變均會引起除草劑抗性(Uchino et al.,2007;Liu et al.,2013)。正是由于結(jié)合部位遠離催化部位,因此這些位點突變并不影響催化部位的功能。Powles和Yu(2010)研究表明,P197S突變未對ALS酶的催化功能造成不利影響。本研究中,苯磺隆與野生型ALS的對接結(jié)果表明,苯磺隆連接在ALS通道入口附近,與McCourt等(2006)、Duggleby等(2008)的研究結(jié)果一致。而其通道入口的位置會阻止底物進入ALS活性中心,缺少底物則致使ALS的酶活產(chǎn)物——支鏈氨基酸合成受阻,影響雜草有絲分裂的正常進行,導(dǎo)致雜草死亡而達到除草目的。
本研究中突變體系P197S_TBM的對接結(jié)果表明,突變后ALS與TBM接觸的氨基酸殘基數(shù)量增加,結(jié)合位點偏C端區(qū)域,結(jié)合位置顯示苯磺隆仍被阻止進入ALS的活性通道,與Uchino等(2007)、Yu等(2012)、Liu等(2013)的研究結(jié)果吻合。P197S_TBM體系MD軌跡數(shù)值更低,結(jié)合自由能更低,結(jié)合更穩(wěn)定。基于本研究結(jié)果,突變導(dǎo)致苯磺隆的結(jié)合位置發(fā)生明顯變化,且突變后形成復(fù)合體更穩(wěn)定牢固。本研究中計算機模擬結(jié)果與除草劑不能到達其催化作用位點而導(dǎo)致抗藥性機理的推測吻合(Dinelli et al.,2005),但產(chǎn)生抗藥性的詳細過程仍需進一步深入研究。此外,ALS靶標(biāo)的抗藥性機制遠比分子結(jié)合模式的解釋復(fù)雜,已有研究表明不同類型的除草劑與除草劑結(jié)合區(qū)域結(jié)合時,其構(gòu)象朝向會有差異,結(jié)合區(qū)域的突變氨基酸類型不同,引起的結(jié)合能變化不同,帶來的結(jié)果也不同。有些突變僅造成ALS構(gòu)象的朝向發(fā)生變化,引起突變體與除草劑結(jié)合能力下降;而有些氨基酸殘基突變,如已被大家熟知的Trp574Leu是改變活性通道形狀而產(chǎn)生除草劑抗藥性。本研究就P197S單位點突變產(chǎn)生苯磺隆抗性的作用機制開展了計算機模擬分析,但具體的抗性機制還需從分子結(jié)構(gòu)和結(jié)合模式等因素之外的除草劑吸收、傳導(dǎo)及代謝途徑等多個非靶標(biāo)因素進行綜合研究。
4 結(jié)論
P197S突變引起ALS與TBM結(jié)合位點和結(jié)合模式發(fā)生變化,接觸氨基酸殘基數(shù)量增加,氫鍵更多,突變體系柔性更低,結(jié)合更穩(wěn)定;促體系結(jié)合的驅(qū)動力類型未發(fā)生變化,促結(jié)合能量項數(shù)值增加,抑制結(jié)合的能力分項降低;417~423位氨基酸殘基在ALS與TBM結(jié)合過程中是較集中貢獻能量的氨基酸團。
參考文獻:
鄧培淵,王輝,袁洪哲,王廣軍,李長看. 2018. 人血清白蛋白與氯氰菊酯相互作用的分子模擬研究[J]. 農(nóng)藥學(xué)學(xué)報,20(1):11-17. [Deng P Y,Wang H,Yuan H Z,Wang G J,Li C K. 2018. Molecular modeling studies of interactions between human serum albumin and cypermethrin[J]. Chinese Journal of Pesticide Science,20(1):11-17.]
鄧維. 2017. 抗苯磺隆播娘蒿抗性機理及抗性突變對乙酰乳酸合成酶功能影響[D]. 北京:中國農(nóng)業(yè)大學(xué). [Deng W. 2017. The resistance mechanisms of tribenuron-methyl-resistant flixweed(Descurainia sophia L.) and effects of resistance-endowing mutations on ALS functionality[D]. Beijing: China Agricultural University.]
付三雄,周曉嬰,戚存扣. 2019. 苯磺隆對甘藍型油菜的殺雄效果及對其靶標(biāo)ALS活性的影響[J]. 江西農(nóng)業(yè)學(xué)報,31(2):8-12. [Fu S X,Zhou X Y,Qi C K. 2019. Male-steri-le-inducing efficiency of tribenuron-methyl and its effect on activity of acetolactate synthase in Brassica napus[J].Acta Agriculturae Jiangxi,31(2):8-12.]
黃新發(fā). 2003. 除草劑靶標(biāo)乙酰乳酸合成酶對抑制劑敏感性及其分子機理的初步研究[D]. 杭州:浙江大學(xué). [Huang X F. 2003. Preliminary study on the susceptibility of herbicide-targeted acetolactate synthase to inhibitors and its molecular mechanism[D]. Hangzhou: Zhejiang University.]
金濤. 2012. 麥田雜草薺菜對苯磺隆抗性分子機制的研究[D]. 泰安:山東農(nóng)業(yè)大學(xué). [Jin T. 2012. Molecular me-chanism of Capsella bursa-pastoris resistant to tribenuron-methyl in wheat fields[D]. Taian:Shandong Agricultural University.]
辛潔,徐小博,王磊,朱曄榮,王勇. 2019. ALS 抑制劑類除草劑的抗性研究概述[J]. 安徽農(nóng)業(yè)科學(xué),47(4):17-21. [Xin J,Xu X B,Wang L,Zhu Y R,Wang R. 2019. Study on the resistance of ALS inhibitor herbicides[J]. Journal of Anhui Agricultural Sciences,47(4):17-21.]
楊冬臣,霍靜倩,張哲,齊萌,張金林. 2019. 反枝莧乙酰乳酸合成酶與煙嘧磺隆分子結(jié)合模式分析及抗性位點預(yù)測[J]. 農(nóng)藥學(xué)學(xué)報,21(1):26-34. [Yang D C,Huo J Q,Zhang Z,Qi M,Zhang J L. 2019. Binding modes of acetolactate synthase with nicosulfuron in Amaranthus retroflexus L. and potential resistance sites prediction[J]. Chinese Journal of Pesticide Science,21(1):26-34.]
張樂樂,郭文磊,李偉,趙寧,劉偉堂,王金信. 2016. 薺菜對乙酰乳酸合成酶抑制劑類除草劑的抗性水平及其分子機制[J]. 農(nóng)藥學(xué)學(xué)報,18(6):717-723. [Zhang L L,Guo W L,Li W,Zhao N,Liu W T,Wang J X. 2016. Resistance to acetolactate synthase-inhibiting herbicides in Cap-sella bursapastoris and its molecular resistance mechanism[J]. Chinese Journal of Pesticide Science,18(6):717-723.]
趙奇,鄧培淵,李玉華,楊玉珍,楊宗渠,雷志華. 2018. 小菜蛾性信息素結(jié)合蛋白與順-9-十四碳烯酸酯作用的分子模擬[J]. 南方農(nóng)業(yè)學(xué)報,49(11):2193-2197. [Zhao Q,Deng P Y,Li Y H,Yang Y Z,Yang Z Q,Lei Z H. 2018. Molecular simulation of interactions between sex pheromone binding proteins and cis-9-tetradecenoic acid ester in Plutella xylostella[J]. Journal of Southern Agriculture,49(11):2193-2197.]
Dinelli G,Bonetti A,Marotti I,Minelli M,Catizone P. 2005. Possible involvement of herbicide sequestration in the resistance to diclofop-methyl in Italian biotypes of Lolium spp.[J]. Pesticide Biochemistry and Physiology,81(1):1-12.
Duggleby R G,McCourt J A,Guddat L W. 2008. Structure and mechanism of inhibition of plant acetohydroxyacid synthase[J]. Plant Physiology and Biochemistry,46(3):309-324.
Guttieri M I,Eberlein C V,Thil D C. 1995. Diverse mutations in the acetolactate synthase gene confer chlorsulfuron resistance in kochina(Kochia scoparia) biotypes[J].Weed Science,43(2):175-178.
Liu W T,Bi Y L,Li L X,Yuan G H,Du L,Wang J X. 2013.Target-site basis for resistance to acetolactate synthase inhibitor in water chickweed(Myosoton aquaticum L.)[J]. Pesticide Biochemistry and Phsinology,107(1):50-54.
McCourt J A,Pang S S,King-Scott J,Guddat L W,Duggleby R G. 2006. Herbicide-binding sites revealed in the structure of plant acetohydroxyacid synthase[J]. Proceedings of the National Academy of Sciences of the United States of America,103(3):569-573.
Panozzo S,Scarabel L,Tranel P J,Sattin M. 2013. Target-site resistance to ALS inhibitors in the polyploidy species Echinochloa crus-galli[J]. Pesticide Biochemistry and Phy-siology,105(2):93-101.
Powles S B,Yu Q. 2010. Evolution in action plants resistant to herbicides[J]. Annual Review of Plant Biology,61:317-347.
Uchino A,Ogata S,Kohara H,Yoshida S,Yoshioka T,Watanabe H. 2007. Molecular basis of diverse responses to ace-tolactate synthase-inhibiting herbicides in sulfonylurea-resistant biotypes of Schoenoplectus juncoides[J]. Weed Biology and Management,7(2):89-96.
Yu Q,Han H P,Li M,Purba E,Walsh M J,Powles S B. 2012. Resistance evaluation for herbicide resistance-endowing acetolactate synthase(ALS) gene mutations using Raphanus raphanistrum populations homozygous for specific ALS mutations[J]. Weed Research,52(2):178-186.
Yu Q,Han H P,Powles S B. 2008. Mutations of the ALS gene endowing resistance to ALS-inhibiting herbicides in Lolium rigidum populations[J]. Pest Management Science,64(12):1229-1236.
Yu Q,Powles S B. 2014. Resistance to AHAS inhibitor herbicides:Current understanding[J]. Pest Management Scien-ce,70(9):1340-1350.
(責(zé)任編輯 麻小燕)
