GA信號因子 SlMYB33對番茄開花時間及果實大小的影響
2020-12-14 07:45:02張波,張杰,張顏,梁燕
西北農業學報 2020年12期
關鍵詞:植物
張 波,張 杰,張 顏,梁 燕
(西北農林科技大學 園藝學院,陜西楊凌 712100)
番茄(SolanumlycopersicumL.)是世界上栽培和消費最廣泛的蔬菜作物之一。在番茄生產過程中,花期調控與果實發育至關重要,其與產量、熟性等密切相關。赤霉素(GA)是一類重要的植物激素,其在植物開花誘導和果實發育過程起著重要作用[1-4]。
隨著GA信號轉導途徑中幾個關鍵調控因子的鑒定和功能分析,人們對GA調控植物生長發育的分子機制有了較為深入的認識[5-8]。GID1(GIBBERELLIN-INSENSITIVE DWARF1)是GA的受體,其能感知GA信號;DELLA蛋白屬于GA信號和植物生長發育的關鍵抑制因子,而GA促進植物的生長發育是通過降解DELLA蛋白來完成的[9]。在GA信號傳導途徑中,具有生物活性的GA首先結合GID1受體,然后與DELLA蛋白相互作用,形成GA-GID1-DELLA三元復合體[9-11],最后該復合體通過泛素化途徑使DELLA蛋白發生水解,消除DELLA的抑制作用,并誘導DELLA下游基因的表達,進而調控植物生長發育[12-16]。例如,DELLA蛋白在植物成花過程中會抑制一些開花誘導基因的表達,而GA促使其快速降解,最終促成花轉變[17-20]。
GAMYB是GA信號途徑中的正向調節因子,屬于DELLA蛋白的下游基因。GAMYB最初是從大麥糊粉層中分離出來的,可以誘導許多GA響應基因的表達[21-23]。近年來,多項研究表明GAMYB在GA調控植物花發育的過程中起著關鍵作用[1]。例如,毒麥LtGAMYB主要在莖尖表達,其轉錄水平在開花誘導期間與GA含量同時上調,表明GA可能通過GAMYB的轉錄調控來誘導植物開花[24]。擬南芥中有3個GAMYB基因:AtMYB33、AtMYB65和AtMYB101[24-25]。擬南芥從短日照條件轉移到長日照條件或在短日照下經外源GA處理均誘導開花,與此同時,AtMYB33和花分生組織屬性基因LEAFY(LFY)在莖尖的表達水平上升,且AtMYB33的表達模式與LFY在莖尖重疊,但表達時間先于LFY[26-27]。此外,AtMYB33可以與LFY啟動子中一個特定的8-bp序列結合[28],表明GAMYB可通過LFY基因的轉錄激活調控植物開花[28-29]。
目前,雖然GAMYB在調控植物開花方面的功能研究已取得了較大進展[8,30-37],但其在番茄中的功能尚不清楚。另外,GAMYB調控果實發育的研究較少,難以理解其在GA調控果實發育中的作用和機制。筆者實驗室鑒定了番茄中的一個GAMYB同源基因SlMYB33,并進行了初步分析,SlMYB33基因定位在6號染色體上,包含3個外顯子和2個內含子,CDS序列全長1515bp,編碼504個氨基酸。SlMYB33氨基酸序列中包含1個R2R3重復序列和2個GAMYB家族特有的保守元件,證明SlMYB33屬于GAMYB家族,但對其功能的探討還非常有限。本研究在前期工作的基礎上,首先通過GUS組織化學染色對SlMYB33的表達進行分析;其次通過該基因在番茄中的過量表達分析其生物學功能,旨在了解GAMYB家族在番茄中是否具有保守或特異的作用。
1 材料與方法
1.1 材 料
1.1.1 植物材料 以西北農林科技大學園藝學院番茄育種課題組保存的Micro-Tom和哥倫比亞生態型(Col)擬南芥為研究材料。Micro-Tom種植在組培室內,白天25 ℃,夜間18 ℃;擬南芥Col種植在光照培養箱內,白天22 ℃,夜間 16 ℃;光周期為光照16 h/黑暗8 h。
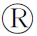
1.2 方 法
1.2.1 GUS組織化學染色 克隆SlMYB33的啟動子序列,將其替換pBI121載體的35S啟動子,構建pSlMYB33∶GUS融合表達載體。利用蘸花法將融合載體轉入到擬南芥Col中[38],經PCR鑒定獲得陽性植株。選取轉基因植株的花序,置于X-Glu溶液中染色過夜(37 ℃、避光),然后在脫色液(乙醇和乙酸體積比為 3∶1)中脫色約3 h,其間適時更換脫色液。脫色之后在體視顯微鏡下觀察染色情況。
1.2.2 過量表達載體構建 過表達載體選擇的是pCAMBIA2307載體。用限制性內切酶XbaⅠ和KpnⅠ對pCAMBIA2307載體以及pMD18-T-SlMYB33進行雙酶切,回收目的片段,用T4連接酶連接載體片段和回收基因片段,連接過夜后轉入DH5α中,37 ℃培養,挑取單斑,進行菌落PCR,選擇符合目的條帶的菌液進行測序。
1.2.3 番茄遺傳轉化 將構建好的SlMYB33過表達載體通過電擊法轉入農桿菌,然后利用筆者課題組已建立的番茄遺傳轉化體系進行轉化:農桿菌侵染5 min,共培養1.5~2.0 d,通過抗生素和分化培養基進行陽性芽篩選,3~4 d后長出2~5 mm的不定芽移栽生根培養基,之后進行煉苗得到轉基因植株。分化培養基:MS + ZT(2 mg/L) + Kan (50 mg/L) + IAA (0.2 mg/L) +6-BA(2 mg/L) + CB (400 mg/L);生根培養基:1/2 MS + IAA (0.2 mg/L)。
2 結果與分析
2.1 SlMYB33在花和果實中的表達特征分析
為預測SlMYB33基因的生物學功能,首先對其表達特征進行分析。構建 pSlMYB33∶GUS融合表達載體并轉入擬南芥中,然后利用GUS組織化學染色觀察該基因在花和果實中的表達情況。結果發現(圖1),與對照相比轉基因植株的雄蕊和柱頭被染成藍色,說明SlMYB33主要在雄蕊和柱頭中表達。
A. SlMYB33在野生型擬南芥中的GUS組織化學分析;標尺=2 mm,下同;B. SlMYB33在轉基因擬南芥中的GUS組織化學分析
2.2 SlMYB33過表達載體的構建
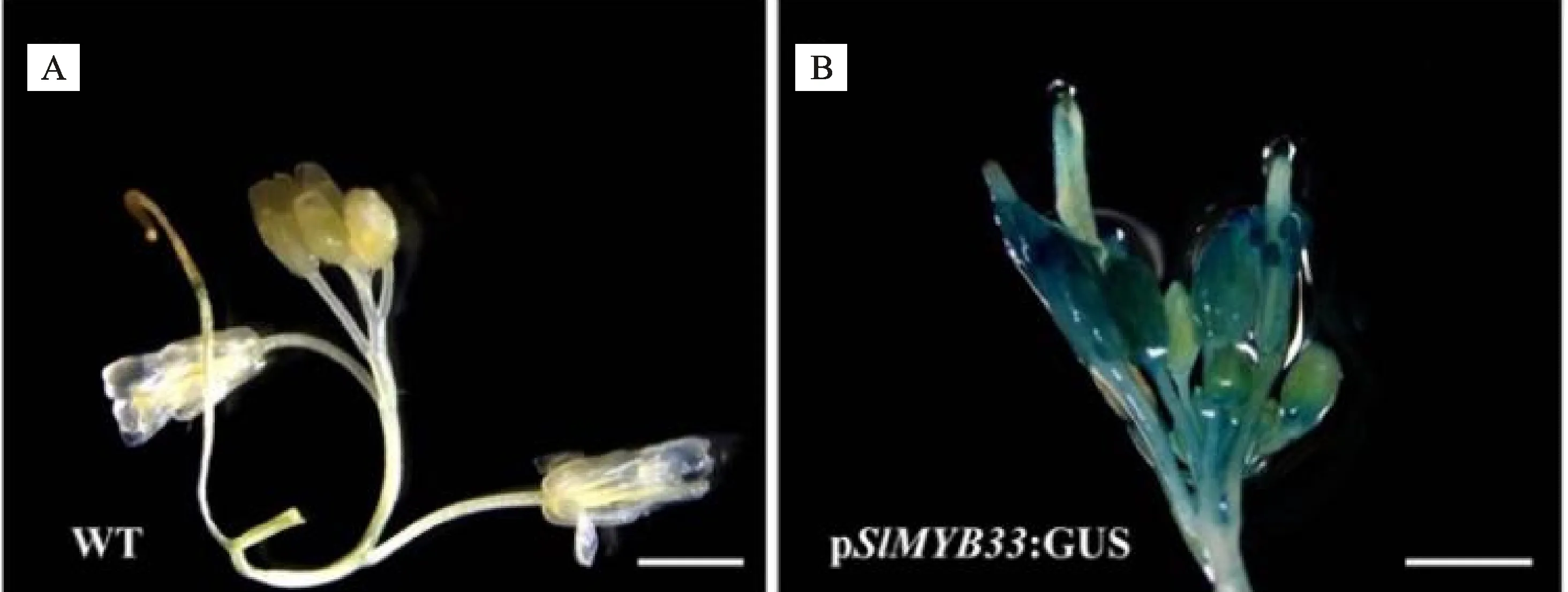
提取番茄花的RNA,反轉錄成cDNA后作為模板,用設計好的引物對其進行PCR擴增,PCR產物經回收、連接、轉化后進行菌落PCR檢測和雙酶切驗證。菌液1~5的PCR擴增條帶與陽性對照一致,均為1 500 bp左右(圖2-A);雙酶切結果顯示,重組質粒可被酶切成1 500 bp和 6 000 bp的條帶,其中1 500 bp為目的基因長度(圖2-B)。最后根據測序結果確定SlMYB33過表達載體構建成功。
A. SlMYB33過表達載體的菌液PCR鑒定;M.DNA 分子標記,下同;pCAMBIA2307- SlMYB33質粒為陽性對照 (+);野生型DNA做陰性對照 (-);泳道1~5.重組質粒菌落PCR;B. SlMYB33過表達載體的雙酶切驗證;CK.pCAMBIA2307- SlMYB33質粒;1~2.雙酶切驗證
2.3 SlMYB33過表達番茄植株的鑒定
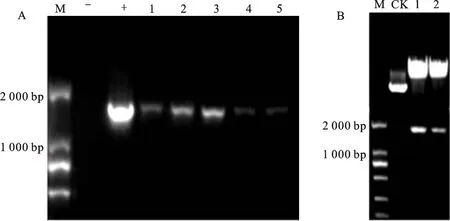
利用農桿菌介導法將SlMYB33過表達載體轉入番茄中,通過PCR鑒定篩選陽性植株。如圖3-A所示,鑒定出11個轉基因株系,進一步對轉基因株系中的SlMYB33表達進行檢測。qPCR結果顯示,轉基因株系OE3、OE4、OE5、OE6、OE7中的SlMYB33表達顯著上調(圖3- B),其中在OE4、OE5、OE7中的表達量均為WT的 10倍以上,因此選擇這3個株系進行下一步 分析。
A. SlMYB33過表達株系的PCR鑒定;M.DNA 分子標記;野生型DNA做陰性對照(-);泳道1~11.植株PCR鑒定結果;B.轉基因植株中的 SlMYB33的表達分析;星號表示過表達株系和野生型之間的顯著差異(* P<0.05,** P<0.01;Student’s t-test), 下圖同
2.4 SlMYB33過量表達對番茄開花時間的 影響
鑒于其他作物GAMYB對開花的影響,筆者統計了SlMYB33過表達T1代植株的開花時間,發現OE4、OE5、OE7株系的始花節位為6~9,顯著大于野生型的4.7(圖4-B),表明SlMYB33過量表達導致番茄開花延遲;與之對應的是,SlMYB33過表達株系中第一朵花開放時間同樣推遲,如OE4和OE7播種后52~61 d第一朵花開放,明顯長于野生型的43.7 d (圖4-A,C)。
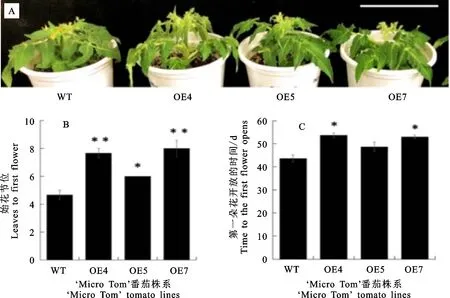
A.野生型和 SlMYB33過表達株系T1代。標尺=10 cm;B.野生型和 SlMYB33過表達株系T1代的始花節位統計。試驗設置6個生物學重復,下圖同;C.野生型和 SlMYB33過表達株系T1代中第一朵花開放時間
2.5 SlMYB33過量表達對番茄果實發育的影響
為進一步了解SlMYB33是否影響番茄果實發育,對SlMYB33過表達株系中的果實表型進行分析(圖5-A)。結果發現,OE4、OE5、OE7株系的成熟果實橫徑為 23.16~26.20 mm,縱徑為20.54~22.03 mm,顯著大于野生型的橫徑 16.17 mm、縱徑15 mm(圖5-B),表明SlMYB33的上調可促使番茄果實增大。
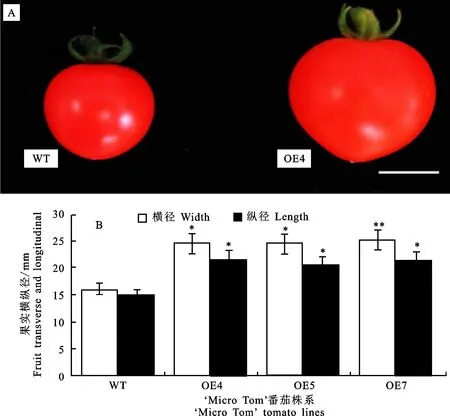
A. SlMYB33-OE系果實顯著大于野生型。標尺=1 cm;B.果實橫縱徑
3 討論與結論
GAMYB是GA信號傳導途徑的關鍵組成部分,其在植物生長發育過程中起著重要作用。目前,關于GAMYB的研究主要集中在種子萌發[1,40]、開花誘導[28-29,37,41]和花器官發育[22-23,30,35-36]等方面。本研究初步鑒定了番茄中GAMYB同源基因SlMYB33的功能,發現其可以調控番茄的開花時間(圖4),說明GAMYB在不同作物間具有保守功能[28-29,37,41-43]。此外,本試驗還揭示了SlMYB33對番茄果實大小的影響(圖5),為進一步探討GAMYB在植物果實發育方面的作用奠定了基礎。
GAMYB是植物開花誘導的促進因子[24],而SlMYB33在番茄中的過量表達卻推遲開花(圖4),推測SlMYB33對開花時間的影響存在劑量效應,即SlMYB33的過量會起到反效果。這種生物學現象在其他物種中同樣出現,例如,大麥HvGAMYB的超量表達抑制了雄蕊發育,類似于GA過量對雄蕊的影響[6,23,44]。
前人研究表明,在GAMYB調控植物開花過程中,LFY是其下游目標基因,即GAMYB通過激活LFY的轉錄誘導植物開花[28,45-46]。番茄中Falsiflora(FA)是LFY同源基因,其功能缺失突變體表現為開花延遲表型[46]。因此推測,番茄FA有可能是SlMYB33的下游靶基因。總之,盡管SlMYB33對番茄開花具有非常重要的作用,但其調控機制尚不清楚,有待進一步研究與驗證。
此外,前人對于GAMYB調控植物開花和花發育的作用機制有了較為深入的研究,但在果實發育調控方面涉及較少。而本研究初步證實了SlMYB33在番茄果實大小調控中的作用,但目前對其調控機制的探討有待進一步挖掘。番茄果實大小受到一系列基因的調控,如FW2.2、FW11.3、OVATE、SUN、FASCIATED(FAS)和LOCULENUMBER(LC)等[47-56],而SlMYB33與這些基因是否存在調控關系以及如何調控將是揭示SlMYB33調控番茄果實大小分子機制的關鍵。
綜上,本研究初步分析了番茄中GA信號轉導因子SlMYB33的表達模式和生物學功能,證明其在番茄開花和果實發育中的作用。下一步將通過定點基因編輯技術獲得SlMYB33的功能缺失突變體,對該基因的功能在花和果實發育等方面進行深入研究,同時篩選鑒定其上下游調控基因,對SlMYB33調控番茄生長發育的分子機理展開研究。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
