低磷脅迫對水稻生長及生理特性的影響*
2020-12-21 09:27:20朱蘭保常曉梅
河北科技師范學院學報 2020年3期
丁 艷,朱蘭保,盛 蒂,常曉梅
(蚌埠學院土木與水利水電工程學院,安徽 蚌埠,233000)
水稻是重要糧食作物之一,全球約有50%以上的人口以稻米為主食[1]。水稻的生長過程中,磷肥是不可缺少的營養元素,磷可增強植株體內活力,促進養分合成和轉運,增強光合作用,延長葉的功能期,有益于水稻增產穩產[2,3]。磷素供應不足會影響水稻的正常生長發育,進而影響其產量和品質[4,5]。
有研究表明,在低磷條件下,作物會從形態和生理等方面做出一些適應性反應,以適應低磷環境[6~8]。劉文菊等[9]研究發現,不同基因型水稻在磷脅迫下其根系形態發生了明顯變化,如根長、根冠比均呈增加趨勢。在低磷條件下,水稻植株分蘗數增減少,且每株根系相互之間有橫向交織生長現象[10]。推測水稻在低磷處理下根系可能會通過橫向生長來擴大與培養液的接觸面積,從而增加對磷的吸收。此外,磷脅迫下水稻可通過提高根系APase 活性增強對磷脅迫環境的適應性[11]。綜上所述,有關水稻對低磷脅迫的適應機制的研究,大多集中在苗期根系的適應性機制,針對水稻地上部組織在形態和生理等方面對缺磷的適應性反應的報道較少。本次研究是在前期研究的基礎上,選取3種不同基因型水稻為研究對象,采用全營養液和低磷營養液培養水稻幼苗,以植株表型、根系形態、生物量及根系分泌物(APaes,H+)等為參數,分析低磷脅迫對水稻生長的影響,以及水稻對低磷的適應性反應,旨在為進一步從分子水平上闡明水稻對低磷脅迫的響應機制提供理論依據。
1 材料與方法
1.1 供試材料
選用鄭旱6號(ZH6,耐低磷基因型)、通粳981(TJ981,磷敏感基因型)和鎮稻99(ZD99,根系擴展型)3種不同基因型水稻品種為試驗材料,供試水稻種子由江蘇潤揚種業股份有限公司提供。
1.2 試驗設計
選擇籽粒飽滿的水稻種子,浸入100 mL/L H2O2溶液中消毒30 min,用去離子水反復沖洗后,放入盛有適量去離子水的培養皿(17 cm)中,催芽。待種子露白時,挑選發芽勢一致的種子放入96孔板置于塑料容器內,采用國際水稻研究所(International Rice Research Institute, IRRI)完全培養液培養[12]。當幼苗長至3葉期時,挑選長勢健壯的幼苗進行分組培養,即正常營養液培養(記為:+P)和低磷營養液培養(記為:-P),且c(+P)∶c(-P)= 25∶1。每個處理選擇4株生長健壯且長勢基本一致的幼苗,每個處理設置10個生物學重復,用泡沫板固定后置于12 L黑色塑料桶中,放置于塑料大棚中。自然光照和溫度條件下,分別用正常供磷和缺磷培養液進行全生育期培養。幼苗生長期,每3 d更換1次培養液,拔節至成熟期,每5 d更換1次培養液。
1.3 測定項目與方法
1.3.1根系形態掃描 分別挑選正常供磷和低磷處理的水稻植株各20株,用去離子水反復沖洗根部后,利用植物根系掃描與分析系統(型號:WinRHIZO,生產商:上海臨嘉科教儀器有限公司生產)掃描根系并進行分析。
1.3.2生物量、葉綠素含量及根系活力的測定 選擇低磷處理15 d的水稻植株,每個處理分別取20株長勢一致的幼苗,用去離子水反復沖洗根系和葉片表面,吸水紙吸干其表面水分,稱其地上部和根部的鮮質量,后置于烘箱中,105 ℃殺青30 min,70 ℃烘干至恒質量,再次稱量地上部和根部的質量。采用差減法計算單株相對含水率,地上部(或根部)干物質質量及根冠比等參數。采用分光光度法測定葉片中葉綠素的質量分數;TTC還原法測定根系活力。
1.3.3根系分泌酸性磷酸酶活性的測定 參考文獻[6]的方法進行,略有改動。取生長健壯的水稻幼苗2株,用去離子水沖洗干凈根系,置于含有100 mL摩爾濃度為1 mmol/L對硝基酚磷酸二鈉(p-NPP)培養液的錐形瓶中,錐形瓶用黑色薄膜包裹,正常光照培養2 h后,吸取1 mL反應液加入到含有5 mL摩爾濃度為1 mol/L NaOH的試管中,搖勻。以無酶反應為空白對照,450 nm處測定吸光度值。根系分泌酸性磷酸酶活性以單位時間單位質量鮮根水解p-NPP生成p-NP的量來表示(單位:mg·(h·g)-1)。
1.3.4根際或根表pH值的顯色觀察-瓊脂糖凝膠法 稱取1.5 g瓊脂糖分別溶于150 mL正常供磷(+P)和缺磷(-P)培養液(pH 5.4)中,于微波爐中溶解,取出后冷卻至40~50 ℃時,加入甲基紅和溴甲酚綠混合酸-堿指示劑,使混合酸-堿指示劑的終質量濃度為0.06 g/L,繼續冷卻至室溫,并分裝于25 mL玻璃試管中。分別選擇萌發后(帶胚乳)和低磷處理3 d后(去胚乳)長勢一致的幼苗,植入裝有10 g/L瓊脂糖的試管中,自然光溫條件下生長,分別在其生長24,48,72 h后觀察瓊脂糖的顏色變化[13]。
2 結果與分析
2.1 低磷對水稻植株表型和根系形態的影響
與正常供磷水稻相比,磷素供應不足導致水稻植株表現出顯著的表型差異(圖1(A))。主要表現為:地上部生長緩慢,植株矮小,葉面積減小;分蘗期延遲且分蘗數減少。此外,低磷導致根系形態發生變化:磷敏感型品種TJ981根長增加明顯(1.37倍),且增長幅度高于耐低磷品種ZH6(1.15倍)和磷高效吸收型品種ZD99(1.31倍);而ZH6和ZD99根平均直徑(1.13 倍和1.25倍)和根表面積(1.45倍和1.82倍)增加幅度均大于TJ981(1.07倍和0.83倍)(圖2)。
低磷處理的不同基因型水稻,其主根長度均較正常供磷水稻主根長明顯增長,尤其磷敏感型水稻品種TJ981主根長度增長較顯著(1.67倍),其次是ZD99(1.57倍),而耐低磷水稻品種ZH6主根長增幅較小(1.41倍)(圖3)。由此可見,磷脅迫導致水稻根系增長,且磷敏感基因型水稻TJ981根系增長幅度顯著高于ZD99和ZH6。也就是說,根系形態變化是水稻對低磷脅迫的一種適應性響應,即低磷時水稻通過改變根系形態,增加與土壤的接觸面積,從而提高其對土壤中磷的吸收和利用。
與對照相比,低磷處理使3種不同基因型水稻分蘗期葉片的長度和寬度均呈減小趨勢(圖3)。其中,低磷處理葉片與對照葉片長度比率分別為ZH6(0.899),TJ981(0.929),ZD99(0.799);葉片寬度比率的變化表現為ZH6(0.922),TJ981(0.843), ZD99(0.810)。本次研究結果可知,低磷脅迫導致水稻開花期較正常供磷水稻延遲,穗形偏小且每株穗數和每穗粒數減少(圖1(B))。這是由于磷素匱乏致使水稻植株葉片的長度和寬度均減小,光合速率下降,光合產物積累量減少,進而影響水稻的產量。
2.2 低磷對水稻幼苗相對含水率和干物質量的影響
本次研究結果顯示,缺磷水稻幼苗相對含水率下降,以ZH6植株含水率下降較大(圖4(A));根部干質量增加(圖4(B)),與TJ981和ZD99相比,ZH6根部干質量增加較多。3個不同基因型水稻植株根冠比變化由大到小依次為:ZH6,ZD99,TJ981(圖4(D))。這是由于低磷導致水稻幼苗地上部生長受阻(圖4(C)),而根系為了增加與土壤的接觸面獲取更多的營養,從而改變其形態。
2.3 低磷對水稻葉片葉綠素質量分數的影響
低磷條件下,水稻植株分蘗期葉片葉綠素質量分數較對照發生變化,其中TJ981和ZH6葉片葉綠素質量分數增加,而ZD99葉片中葉綠素質量分數呈下降趨勢(圖5(A));進入孕穗期以后,與分蘗期相比較,ZH6和TJ981植株葉片的葉綠素質量分數均下降,ZD99變化不明顯;與對照相比,3種不同基因型水稻葉綠素質量分數均表現為下降(圖5(B))。可見,隨著低磷處理時間的延長,水稻葉片中葉綠素質量分數降低。

圖1 低磷處理條件下,不同基因型水稻幼苗表型(A)分蘗期;(B)抽穗、灌漿期
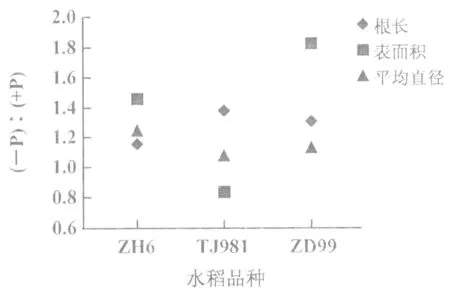
圖2 低磷處理2周后水稻根系形態指標的變化
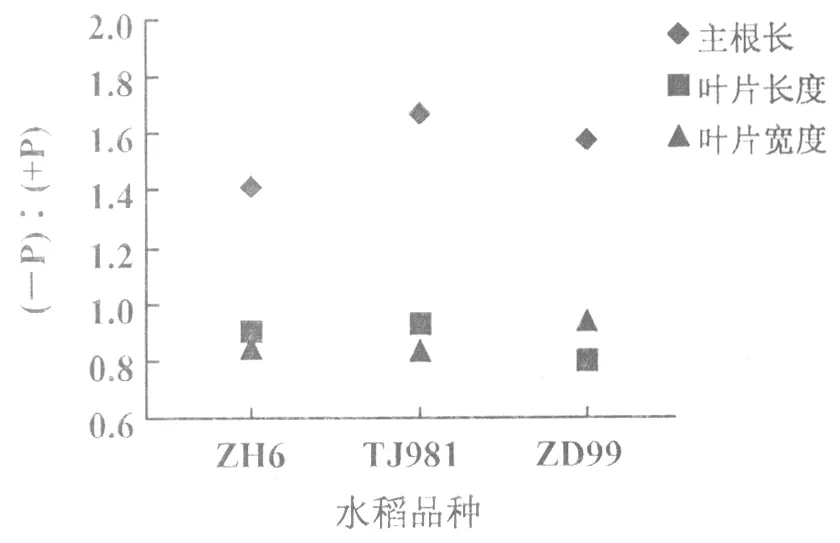
圖3 水稻分蘗期的主根長及葉片變化
2.4 低磷處理對水稻幼苗根系活力的影響
低磷處理不同基因型水稻幼苗15 d后,其根系活力均呈下降的趨勢(圖6)。即低磷脅迫下磷敏感型品種TJ981,根系擴展型ZD99和耐低磷品種ZH6根系活力有不同程度的下降,其中ZH6和ZD99根系活力呈顯著下降(p<0.05),根系活力下降率分別為42.01%和19.70%,而TJ981根系活力呈極顯著下降(p<0.01)。這表明低磷脅迫影響水稻根系的活力水平,且TJ981較其它2種基因型水稻根系活力下降幅度大(下降率為51.48%),可見磷敏感型水稻TJ981對低磷的適應能力弱。
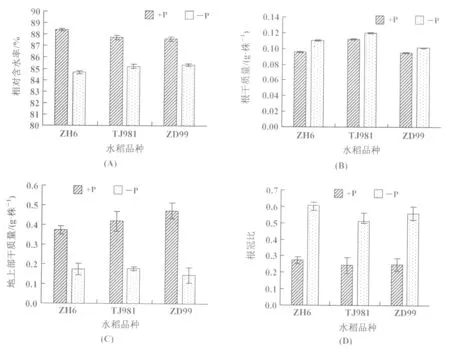
圖4 低磷處理(3周)水稻植株生物量的變化(A):相對含水率;(B):根部干質量;(C):地上部干質量;(D):根冠比
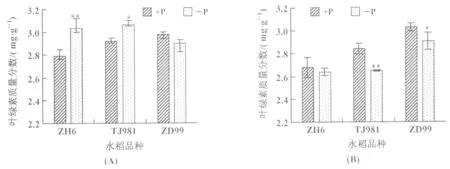
圖5 葉綠素質量分數的變化(A):分蘗期;(B):孕穗期注:*和**分別代表p<0.05和p<0.01,下同。
2.5 低磷處理對水稻幼苗根系分泌酸性磷酸酶(APase)活性的影響
低磷處理3種不同基因型水稻幼苗1,8,15 d后,其根系分泌的酸性磷酸酶活性均高于對照,但不同基因型水稻幼苗根系分泌的磷酸酶活性增長表現不一(圖7)。其中:耐低磷品種ZH6低磷處理1 d時,根系分泌的酸性磷酸酶活性較對照增強52.7%;低磷處理8 d和15 d時,其活性增長趨于穩定,增幅分別為9.1%和9.7%。ZD99根系分泌酸性磷酸酶的活性隨處理時間的延長而增強,分別為5.5%,26.0%和40.1%;而磷敏感性品種TJ981隨低磷處理時間的延長,其根系分泌酸性磷酸酶的活性增強相對穩定,增幅分別為12.5%,11.0%和11.6%(圖7)。可見低磷脅迫使不同基因型水稻根系分泌酸性磷酸酶活性增強。
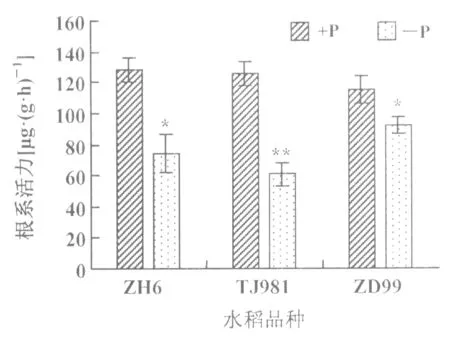
圖6 低磷條件下水稻根系活力的變化
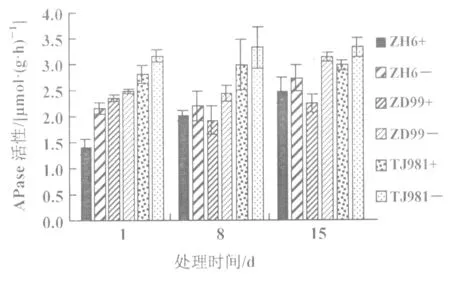
圖7 低磷條件下水稻根系APase活性的變化
2.6 低磷處理對水稻幼苗根際或根表pH的影響
本次試驗以耐低磷品種ZH6為例,采用10 g/L 瓊脂糖凝膠觀察根系分泌酸的變化。主要原理是:甲基紅-溴甲酚綠酸堿混合指示劑的變色點為 5.1,pH > 5.1 呈綠色,pH < 5.1 呈酒紅色。與對照相比,缺磷水稻幼苗根系分泌酸使瓊脂糖變紅色,而且隨著處理時間(24,48,72 h)的延長,瓊脂糖的紅色加深,這表明磷脅迫促進了根系分泌酸的增多(圖8)。同時,在幼苗移植時采用帶胚乳(圖8(A))和去胚乳(圖8(B))2種情況進行對比發現,同樣低磷處理48 h,移植時帶胚乳的幼苗根系比去胚乳的幼苗根系分泌的酸量多,即帶胚乳幼苗根系分泌酸使瓊脂糖呈較深的紅色。這表明幼苗生長前期胚乳可以提供部分營養供其生長發育所需,在沒有胚乳營養的條件下,幼苗生長力減弱,分泌酸減少。但相比于對照,低磷處理的根系分泌酸量增加,瓊脂糖紅色明顯。
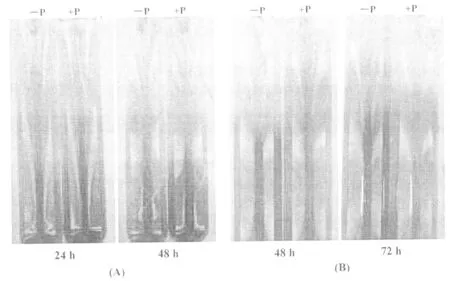
圖8 低磷對ZH6幼苗根系分泌酸的影響(A)萌發后帶有胚乳植入;(B):萌發后去胚乳植入
3 討論與結論
水稻是對磷營養較敏感的作物。低磷脅迫下,為了適應逆境條件水稻會產生一系列的適應性響應。黃榮等[14]研究表明,缺磷會影響植物的生長發育且產生相應的癥狀,如植株生長遲緩、矮小、分枝數減少。本次試驗結果表明,低磷脅迫會使水稻植株矮小、葉小、分枝、分蘗減少,這是由于低磷影響植株體內核蛋白的形成,從而抑制細胞分裂與增殖,導致植株生長發育延緩[15]。根系是最先感受養分脅迫信號并對這一逆境信號加工、處理和傳遞[4],其形態變化是水稻適應低磷脅迫的機制之一,如主根增長,側根數目增多,根表面積增大[16]。本次試驗結果顯示,低磷處理水稻幼苗2周后,耐低磷水稻品種鄭旱6號在低磷條件下其根系長度變化不明顯,但其平均直徑和根面積均呈增加趨勢;而磷敏感型品種通粳981根系增長、根平均直徑和根面積減小,根系呈細長型(圖2)。因此,由于缺磷改變根系形態并使其根質量增加,從而促進根冠比增大。此外,低磷條件下,分蘗期水稻根長顯著增長。鄭旱6號,通粳981和鎮稻99的根長增幅分別為對照的1.4,1.7和1.6倍。可見低磷脅迫時水稻可以通過增加根系長度,增大根表面積等,擴大與土壤的接觸面積,進而增加對磷素的吸收和利用效率,滿足自身生長所需的磷營養[17]。
低磷脅迫會誘導植物產生一系列的生理生化反應以調節植物的生長發育。本次試驗結果顯示,低磷處理水稻幼苗,使其葉綠素的質量分數在分蘗期和孕穗期表現不一。其中,鄭旱6號和通粳981葉片中葉綠素的質量分數先增后減,鎮稻99葉綠素的質量分數在分蘗期和孕穗期均減少。這是因為隨著低磷處理水稻時間的延長,低磷限制了葉綠素的合成使其含量降低。此外,低磷水稻抽穗期延遲,且每株穗數及穗粒數減少,這是由于低磷水稻葉片中葉綠素質量分數的減少,減弱了水稻植株的光合能力,影響光合產物的形成和積累,進一步降低了水稻的產量。
在生理學上,植物可以通過根分泌酸性磷酸酶(APase)活性的提高和小分子量有機酸的增加,從而減小根際的pH。APase是一種誘導酶,當磷是植物生長的主要限制因子時,植物根系就向胞外分泌酸性磷酸酶,增加對土壤有機磷的水解[6],其活性受植物供磷狀況的影響,低磷能誘導根系APase活性顯著升高[11]。本次試驗結果也顯示,低磷處理3種不同基因型水稻根系分泌酸性磷酸酶活性均有不同程度的提高。根際酸化是植物,特別是雙子葉植物,應對低磷脅迫的一種響應。本次研究發現,低磷促使水稻根系分泌酸增多,根系分泌的有機酸與難溶性磷酸鹽進行陰離子交換或競爭性吸附,以釋放或溶解難溶性磷酸鹽,從而提高磷的利用率。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
中國生殖健康(2019年2期)2019-08-23 08:12:08
產品可靠性報告(2017年7期)2017-09-05 09:49:12
