基于COI和Cytb基因的黃腰響蜜系統發育分析和分歧時間估算
2021-01-13 04:01:20元青苗段玉寶
野生動物學報 2021年1期
元青苗 段玉寶*
(1.西南林業大學,云南省高校極小種群野生動物保育重點實驗室,昆明,650224;2.西南林業大學生物多樣性保護學院,昆明,650224)
隨著分子生物學的發展,更多的鳥類(Aves)mtDNA基因全序列被測定[1],線粒體基因組作為一種研究手段在揭示鳥類的起源、系統發生[2-3]、分歧時間的研究中發揮著重要作用[4-5]。在mtDNA的13個蛋白編碼基因中,COI和Cytb基因常用于動物類群的分類和系統發生關系的研究中[6-7],具有良好的解析作用。
1 材料與方法
1.1 材料數據來源
用于研究的樣品采集于云南省西北部的瀘水縣高黎貢山,在此區域中獲得1只黃腰響蜜個體;取0.5 g左右的肌肉組織,采用無水乙醇進行浸泡,保存于西南林業大學生物多樣性保護學院。其余19個物種的COI和Cytb基因序列均來自于NCBI(National Center for Biotechnology Information)數據庫。其中,將犀鳥目(Bucerotiformes)作為外群[11],共下載2條序列。每條COI和Cytb序列的長度分別大于650 bp和861 bp。物種信息詳見表1。
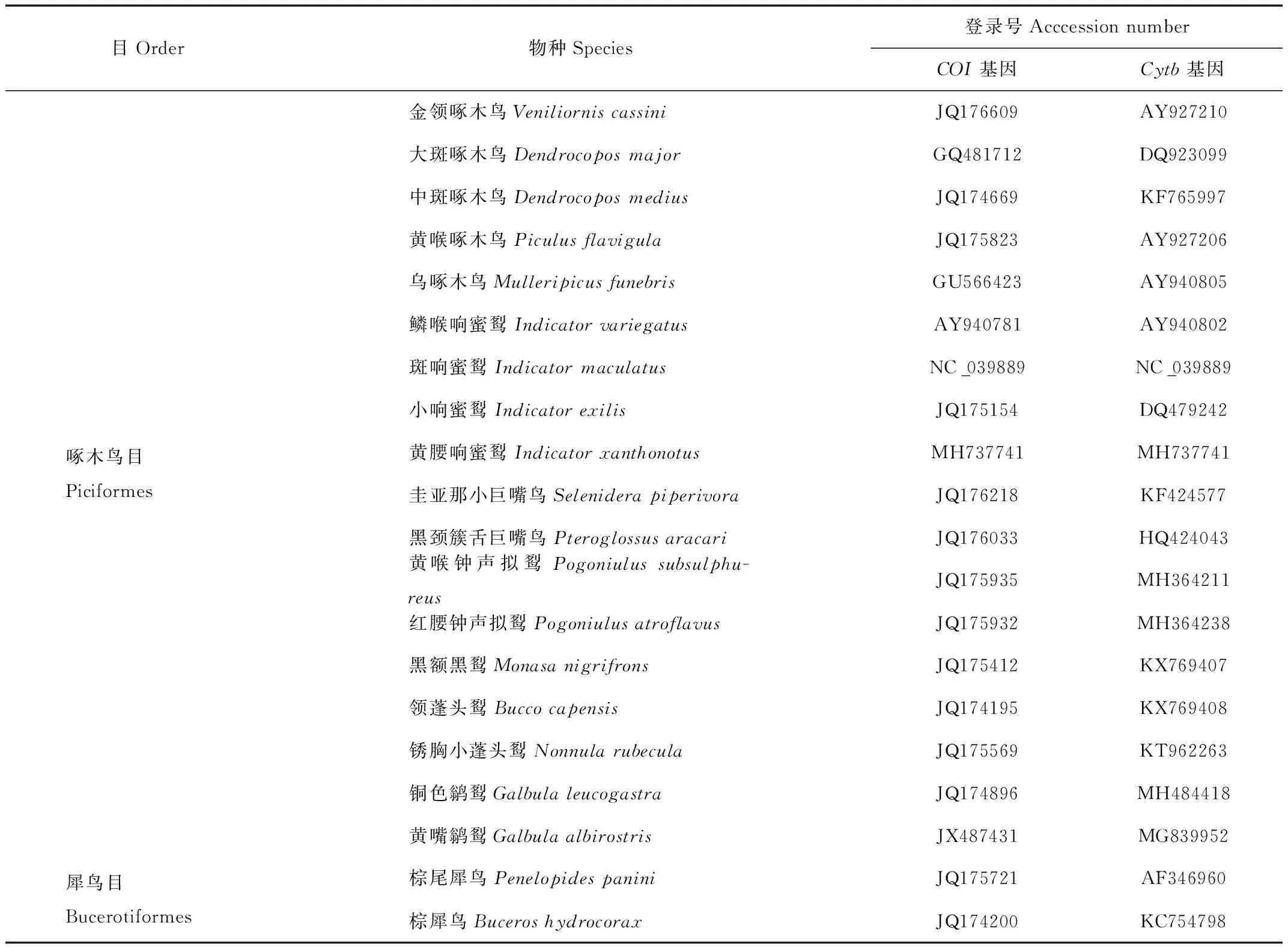
表1 本研究所需的物種基因序列信息Tab.1 List and gene information of species in this study
1.2 DNA的提取、擴增及測序
從采集的肌肉組織樣品中提取總線粒體DNA,DNA提取完成后加入100 μL ddH2O進行溶解,保存于-20℃的冰箱中。
線粒體基因組的引物和反應體系參照Sorenson等[12]的研究。DNA提取和PCR初步擴增完成后,經過1%的瓊脂糖凝膠電泳檢測,在紫外燈下觀察,然后將擴增成功的產物送往上海派森諾生物科技有限公司進行測序。黃腰響蜜的線粒體全基因組信息也已提交NCBI數據庫,登錄號為MH737741。
2 數據處理
2.1 序列處理
在MEGA 7軟件[13]中導入目標物種的序列,20條序列進行多重比對,比對好后對序列進行剪切,每條COI基因序列的最終長度均為650 bp,Cytb序列為861 bp。然后將比對剪切好的COI和Cytb序列分別整理成Fasta和Mega格式進行保存。
2.2 線粒體COI和Cytb基因的飽和性檢驗
運用DAMBE 5.2.63軟件[14]分析所有物種的序列飽和性。如果Iss與Iss.c滿足Iss 在MEGA7中,選用Kimura 2-parameter模型[15]計算響蜜科響蜜屬4種鳥類的種間遺傳距離。 使用貝葉斯法(bayesian inference,BI)[16]和最大似然法(maximum likelihood,ML)[16]建樹。在ClustalX 1.83軟件中轉換整理好的序列文件的格式,保留為Nexus和Phylip格式。 構建樹算法可以采用馬爾可夫鏈蒙特卡羅(MCMC)算法提供所有樹的驗證頻率。最高的后驗概率樹是最優解,也是最優選擇的樹[17]。在jModelTest 0.1.1軟件[18]中算出核苷酸替代類型數(nst)、位點間速率分布(rates)等參數以及分析出研究系統發育關系最優的模型為GTR+I+G。MrBayes軟件是構建BI樹的常用軟件[19],該軟件只支持貝葉斯法建樹,所以本文運用MrBayes v3.2.1構建BI樹。建樹時使用4個馬爾可夫鏈,外群則選擇棕犀鳥(Buceroshydrocorax)。運行后,共進行2次run,二者相互獨立,將起始的25%作為burn-in刪除。運行20萬代后,給出的標準誤差(sd)小于0.01,則運算結束。最后,在Figtree v1.2.2中查看生成的文件。 最大似然法建樹則是通過概率的計算進行的,ML系統發育樹可以利用MEGA7進行構建。 使用BEAST v2.5.2軟件[20-21]估算黃腰響蜜的分歧時間,此方法根據Jarvis等[11]依據鳥類化石記錄對多個目的分歧時間估算的結果作為校正點,啄木鳥目和佛法僧目(Coraciiformes)的分化時間約為43 Ma,犀鳥目的分化時間發生約為58 Ma。將 “*.nxs”格式的文件用于分析,采用松弛的分子鐘(relaxed clock log normal)模型,樹的先驗值計算選擇Yule Model模型,運行1 000萬代。使用Tracer v1.5評估結果的穩定性,之后再進一步分析,最終得到一個含有物種分歧時間的樹文件。可在Figtree v1.2.2中查看含有黃腰響蜜分化時間的文件。 通過DAMBE 5.2.63軟件分析20個物種的核苷酸序列飽和性,導入Fasta格式的序列文件,COI序列分析結果顯示Iss=0.191 3 此外,為了更加直觀地看出核苷酸序列的飽和程度,做飽和度分析的散點圖(圖1A和圖1B),橫坐標為F84遺傳距離;縱坐標為轉換Ts和顛換Tv數。圖1A顯示,當F84距離為0.330 8時,Ts與Tv并未相等,未出現飽和現象,則本研究所獲得的COI序列可以構建系統發育樹。同樣地,當F84距離為0.441 8時,Ts與Tv未相等,未出現飽和現象(圖1B),則本研究所獲得的Cytb序列可以構建系統發育樹。 表2 基于COI基因的響蜜科鳥類遺傳距離Tab.2 Genetic distance between species of Indicatoridae based on COI 表2 基于COI基因的響蜜科鳥類遺傳距離Tab.2 Genetic distance between species of Indicatoridae based on COI 序號No.種名Species12341鱗喉響蜜Indicator variegatus2斑響蜜Indicator maculatus0.0213小響蜜Indicator exilis0.1120.0934黃腰響蜜Indicator xanthono-tus0.1200.1200.154 表3 基于Cytb基因的響蜜科鳥類遺傳距離Tab.3 Genetic distance between species of Indicatoridae based on Cytb 表3 基于Cytb基因的響蜜科鳥類遺傳距離Tab.3 Genetic distance between species of Indicatoridae based on Cytb 序號No.種名Species12341鱗喉響蜜Indicator variegatus2斑響蜜Indicator maculatus0.0323小響蜜Indicator exilis0.1320.1264黃腰響蜜Indicator xanthono-tus0.1770.1780.194 利用COI(圖2A)和Cytb(圖2B)基因重建的BI和ML系統發育樹結構顯示出兩種基因構建的系統樹拓撲結構基本一致,具有相似的系統發育關系,以BI樹的拓撲結構作為展示。據圖顯示,黃腰響蜜與響蜜科響蜜屬聚為一大支(A:PP=0.99,BP=82;B:PP=1.00,BP=100),與啄木鳥科互為姐妹群,并有較高的支持率(A:PP=0.97,BP=75;B:PP=0.96,BP=74)。在響蜜科內部,黃腰響蜜構成單系;在4種鳥類中,最早分化出來的類群是黃腰響蜜,其次是小響蜜(Indicatorexilis)。鱗喉響蜜(Indicatorvariegatus)與斑響蜜(Indicatormaculatus)聚為一支(A:PP=0.98,BP=91;B:PP=1.00,BP=100)。 Tracer v1.5軟件中顯示Ess值大于200,說明在BEAUti中設置的參數是合理的,最后得到含有物種分歧時間的樹文件。響蜜科鳥類的分化時間詳見圖3,可知響蜜科鳥類起源于新第三紀中新世時期,距今約20.44(15.88,25.77)Ma,而黃腰響蜜也在此時間段內分化出來(20.44 Ma)。此外,在漸新世和中新世階段,啄木鳥目鳥類的多樣性不斷擴大,物種多樣性越來越豐富。 COI和Cytb基因片段序列被廣泛地應用于解決鳥類的分子系統發育問題[22]。本文的結果顯示,黃腰響蜜聚在響蜜科響蜜屬物種所在分支里,并且有著很高的支持率。黃腰響蜜所在的響蜜科與啄木鳥科有著姐妹群的關系,這與Prum等[23]的研究結果一致。而在響蜜科內部,黃腰響蜜是最早分化出來的物種,其次是小響蜜;且黃腰響蜜無姐妹群。 本研究分子定時的結果顯示啄木鳥科分化于14.95(14.02,15.89)Ma,與Prum等[23]的研究結果(Prum等研究表明啄木鳥科的分化時間約為15 Ma)相近,表明了分歧時間推斷的合理性和準確性,進而為評估黃腰響蜜的分歧時間提供支持。響蜜科鳥類的分化可追溯到新生代新第三紀中,黃腰響蜜在新第三紀中新世階段分化,時間節點為20.44(15.88,25.77)Ma。在這一時期,青藏高原發生了廣泛的隆起(22—20 Ma),并隨著多個隆升事件[24-25],直到3.4 Ma到1.6 Ma,高原抬升4 000 m以上,這被認為顯著影響了其形貌和鳥類物種的多樣化[26]。所以黃腰響蜜的形成可能與青藏高原隆起有關,其歷史和地理模式受到了青藏高原及周邊區域隆升的強烈影響。2.3 響蜜科鳥類種間遺傳距離計算
2.4 系統發育樹的構建
2.5 分歧年代評估
3 結果與分析
3.1 基因序列飽和性分析
3.2 響蜜科鳥類種間遺傳距離分析
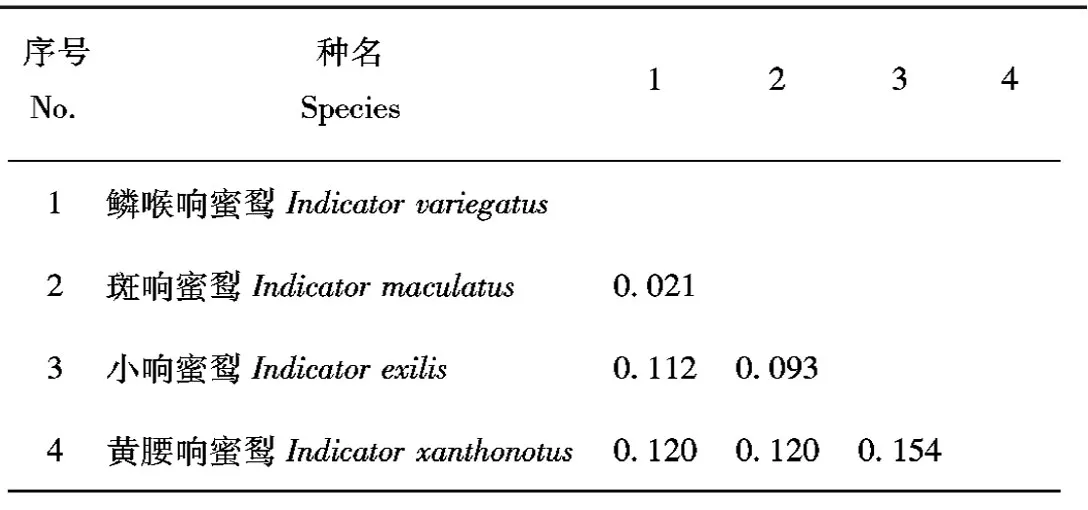
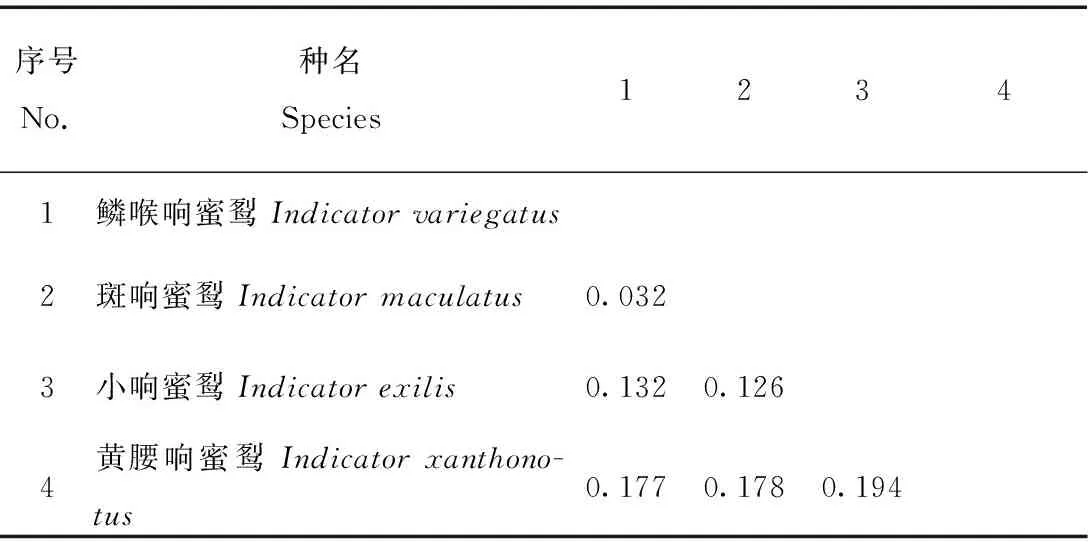
3.3 系統發育樹分析
3.4 分歧時間分析
4 討論
4.1 黃腰響蜜的系統發育分析
4.2 黃腰響蜜的起源分化
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00英語世界(2023年10期)2023-11-17 09:18:18民用飛機設計與研究(2020年4期)2021-01-21 09:15:02科學大眾(中學)(2019年3期)2019-05-17 10:04:30電子制作(2018年18期)2018-11-14 01:48:24汽車觀察(2018年10期)2018-11-06 07:05:26山東工業技術(2016年15期)2016-12-01 05:31:22少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
