小飛蓬捕光葉綠素結合蛋白基因CcLhca-J9克隆及表達分析
2021-01-14 03:03:18李祖任羅丁峰柏浩東徐晶晶韓進財徐強王若仲柏連陽
中國農業科學 2021年1期
關鍵詞:除草劑
李祖任,羅丁峰,柏浩東,徐晶晶,韓進財,徐強,王若仲,柏連陽,?
1湖南省農業科學院雜草生物學及安全防控生物學湖南省重點實驗室,長沙 410125;2湖南農業大學植物激素與生長發育湖南省重點實驗室,長沙 410128
0 引言
【研究意義】小飛蓬(Conyza canadensis)為菊科(Asteraceae)飛蓬屬(Conyza)的一種具較強繁殖和環境適應能力的入侵雜草,主要分布于果、桑、茶園、草坪和荒地[1]。小飛蓬已對草甘膦、草銨膦、莠去津等多種化學除草劑產生不同程度的抗藥性,例如浙江寧波的兩個抗草甘膦種群抗性指數分別達8.28和7.92[2-3]。據報道,當小飛蓬1 m2發生數為72株時,油菜角果數、角果籽粒數顯著下降,產量下降達35.93%,可見惡性雜草小飛蓬給農業生產帶來較大的經濟損失[4]。植物化感物質的挖掘和利用是開發植物源除草劑的有效途徑,是治理小飛蓬等抗藥性雜草的熱點方向之一[5]。發掘新穎生物源化合物結構、除草作用機制和靶標,闡明活性化合物與雜草相互作用機制,利用生物除草關鍵基因資源,可最終克服生物除草劑產品發展的成本與環境制約因素[6]。【前人研究進展】目前,以天然產物為基礎研發的生物除草劑的作用機制主要有五大類,但是許多天然產物的抑草作用靶點和分子機制仍然是未知的[7]。光合系統是除草劑作用的重要靶標之一,其中光系統I和光系統II是吡啶類和三嗪酮類除草劑的靶標[8]。鏈霉菌(Streptomycesspp.)產生的多肽L-phosphinothricin抑制谷氨酰胺合成酶,鏈格孢菌(Alternari alternata)產生的細交鏈孢菌酮酸(tenuazonic acid,TeA)阻斷光合系統 II[9-10]。上述抑草靶標的發現,為生物除草劑的開發提供了科學依據。羊脂酸分子式為C8H16O2,油狀液體不溶于水,屬于有機酸類化合物,主要用于染料、防腐劑、殺菌劑[11]。筆者所在課題組前期從椰子中分離提取出羊脂酸,并驗證其為一種具有廣譜、高效抑草活性的植物源化合物,具有開發成為植物源除草劑的潛力[12]。采用透射電鏡技術觀察到羊脂酸處理小飛蓬葉片后,葉綠體和線粒體結構隨著作用時間的延長出現了空腔、裂解的現象[13]。采用 Lable-free蛋白質組學技術分析羊脂酸處理小飛蓬葉片,發現LHC蛋白(UniProt ID:A0A103Y9J9)響應了羊脂酸處理,并與光合指標測定結果相一致,但該蛋白是否為羊脂酸抑制小飛蓬的靶標蛋白仍需進一步研究[14]。捕光葉綠素a/b結合蛋白(light harvesting chlorophyll a/b-binding protein,LHC)是光系統 I和光系統 II中獲取光能并將其傳送至光反應中心的色素蛋白復合體[15]。在高等植物體內,編碼LHC(LHCI、LHCII)蛋白是一個多基因家族,定位于細胞核內;LHCI蛋白由Lhcal、Lhca2、Lhca3和Lhca4基因編碼,LHCII蛋白由Lhcbl、Lhcb2、Lhcb3、Lhcb4、Lhcb5和Lhcb6基因編碼[16]。隨著分子克隆和基因測序技術的發展,部分植物的LHC蛋白基因已被克隆出來,例如擬南芥(Arabidopsis thaliana)、橡膠樹(Hevea brasiliensis)、秈稻9311(Oryza sativa)[17-19]。【本研究切入點】目前的研究大部分集中在各植物 LHC蛋白及其基因的表達差異、功能解析等,對于LHC蛋白與抑草活性脅迫相關的功能解析研究較少[20]。【擬解決的關鍵問題】通過分子克隆和生物信息學等技術全面闡釋小飛蓬捕光葉綠素a/b結合蛋白基因(CcLhca-J9)的分子特征,解析羊脂酸處理下該基因表達的特殊性,預測羊脂酸作用LHC蛋白的關鍵位點,為后續基因功能驗證打下基礎,同時為植物源羊脂酸產品研發提供科學依據。
1 材料與方法
1.1 供試材料
小飛蓬種子2018年9月采集于湖南省長沙市芙蓉區湖南省農業科學院院內荒地并保存于種子超干低濕存儲柜(購于北京明日百傲生物科技有限公司),選出飽滿種子用0.4%赤霉素溶液催芽后,挑選吐白的種子移入塑料盆缽(內徑20 cm),置于光照培養箱(8 h黑暗、溫度 18℃/16 h光照、光強 100—120 μmol·m-2·s-1、溫度 20℃)培養至三葉期待用。羊脂酸(純度99%)采用蒸餾萃取法自制,具體制備方法參考文獻[12]。
1.2 CcLhca-J9的克隆
從NCBI中選擇保守性高的Lhca基因同源序列設計簡并引物CcJ9-1(表1),進行PCR擴增獲取目的基因片段。PCR反應程序:94℃預變性5 min;94℃變性30 s,55℃退火30 s,70℃延伸30 s,循環30次;70℃延伸7 min。凝膠電泳檢測PCR產物,切膠回收純化目的片段,連接到pCambia2301載體,連接轉化大腸桿菌 DH5α(購于南京諾唯贊生物科技有限公司),挑取陽性克隆送上海生工生物工程有限公司測序[17]。根據已知目的片段設計5′和3′ RACE特異引物(表1),采用TaKaRa公司Clonetech SMARTer RACE 5′/3′ Kit擴增試劑盒說明擴增。獲得Lhca的 5′ RACE序列、核心序列及3′ RACE序列,根據三者拼接后序列設計引物(表1)確定CcLhca-J9全長。
1.3 生物信息學分析
采用DNAMAN分析核酸序列并檢測編碼氨基酸序列,ExPaSy在線預測分子量和等電點(molecular weight/isoelectric point,MW/pI),在 NCBI中搜索LHC的高相似度氨基酸序列,選取19個典型的植物蛋白,先用ClustalX軟件多序列比對,再用MEGA 4.1軟件的鄰接法(neighbor-joining,NJ)構建進化樹[21]。利用 SWISS-MODEL在線搜索與選擇模板,再通過Swiss-PdbViewer軟件優化模型結構[22]。
1.4 分子對接
采用上一步 LHC同源建模結果作為模型來尋找羊脂酸與LHCI-J9蛋白的對接方式。在AutoDock 4.2軟件上運行,電荷計算采用 RESP(Restrained electrostatic potential)法;活性位點采用AutoGrid格式點搜索,網格大小為60?×40?×60?,格點間隔為1.0?。中心設定為配體在LHC中的質心;采用LGA(Lamarckian genetic algorithm)法計算配體與受體間的自由能,其余參數均為缺省值[23]。在最終得到的10個最優構象中,選取能量值最低的構象作為進一步研究對象。
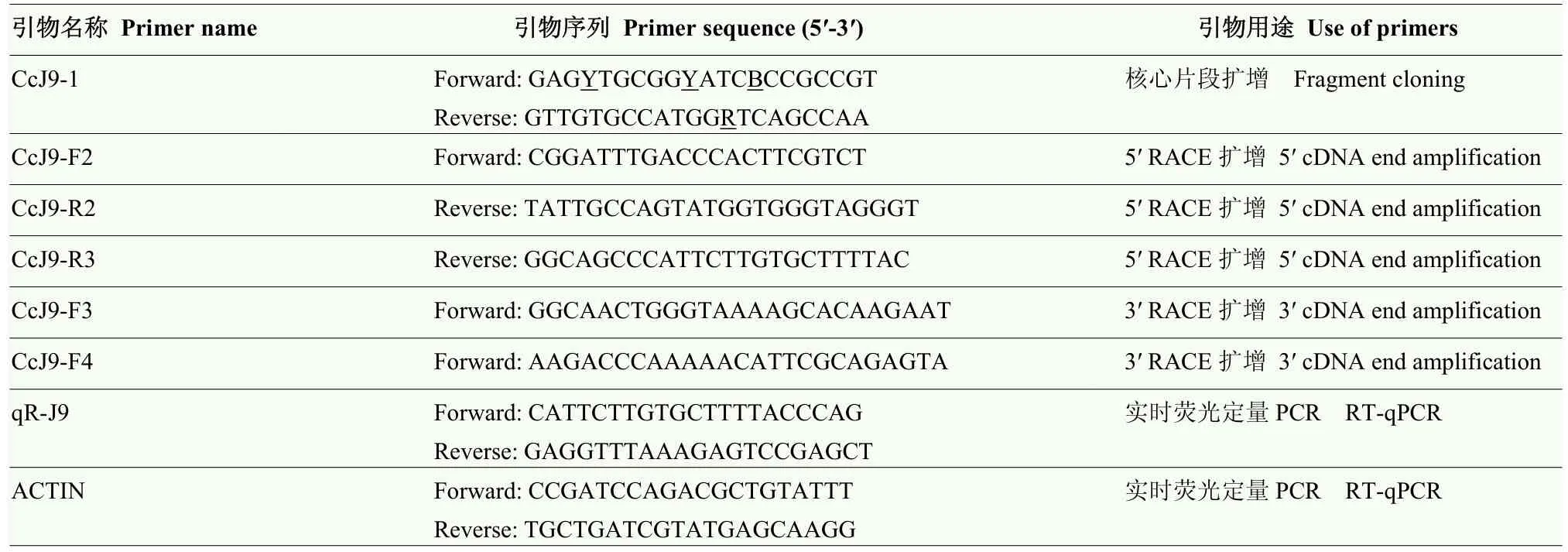
表1 CcLhca-J9克隆及實時熒光定量PCR表達引物Table 1 The primers used for CcLhca-J9 cloning and RT-qPCR
1.5 CcLhca-J9響應羊脂酸的表達分析
用羊脂酸(625 μmol·L-1)、阿魏酸(625 μmol·L-1)和清水對照各10 μL,兌水10 mL,噴霧處理小飛蓬葉片0、0.5、1、2、4、8 h后,收集新鮮葉片立即置于液氮。參照 Trizol改良法提取小飛蓬葉片總RNA,反轉錄合成第一鏈cDNA。RT-qPCR反應體系 20 μL:SYBR Premix ExTaqTM10 μL、引物(R和 F)0.5 μL、cDNA 5 μL、ddH2O 4 μL。反應程序:95℃ 30 s,58℃ 20 s,循環40 次。所用引物為目的基因特異性引物qR-J9和內參基因Actin引物(表1)。數據處理運用 2-ΔΔCT法,比較目的基因的表達水平[24]。利用SPSS軟件中Duncan氏新復極差法進行差異顯著性分析。
2 結果
2.1 CcLhcIa-J9序列的獲得
以反轉錄未施藥的小飛蓬葉片cDNA為模板,用簡并引物CcJ9-1進行PCR擴增、回收和測序,獲得CcLhca-J9核心序列483 bp(圖1-A)。用引物Cc-F2、Cc-R2、Cc-R3、Cc-F3和Cc-F4進行三輪PCR擴增,產物經回收和測序,獲得cDNA 5′端序列428 bp和3′端序列253 bp(圖1-B)。將5′ RACE序列、核心序列及 3′ RACE序列用 DNAMAN 軟件拼接,獲得CcLhca-J9cDNA序列全長(圖2)。
2.2 CcLhcIa-J9生物信息學分析
經同源克隆和 RACE克隆,獲得了CcLhca-J9的全長cDNA序列,該基因閱讀框長744 bp,編碼247個氨基酸(圖2)。編碼蛋白分子質量為26.766 kD,理論等電點為6.43;該蛋白中25個丙氨酸,占總數的 10.1%;其次是脯氨酸和甘氨酸,分別為 24和23個,各占9.7%和9.3%。CcLhca-J9蛋白保守區預測顯示,該蛋白屬于Chloroa_b-bind家族,具有典型的捕光葉綠素a/b結合蛋白功能(圖3)。系統進化結果表明,不同物種來源的Lhca蛋白在分屬不同分支,CcLhca-J9蛋白與除蟲菊(GEW73959.1,Tanacetum cinerariifolium)和黃花蒿(PWA35049.1,Artemisia annua)Lhca蛋白進化程度最為接近,同處于菊科這一分支(圖4)。預測二級結構發現,蛋白具有螺旋(Hh)97個,占39.27%;β轉角(Tt)7個,占2.83%;延伸鏈(Ee)9個,占3.64%;無規則卷曲(Cc)134個,占54.25%(圖5)。LHC-J9蛋白以4y28.1.O(2.80?)為模板進行同源建模,序列相似度為 89.9%,高度同源結構,GEQE=0.75,QMEAN=-2.20,可見所獲得的模型質量是較為可靠的。CcLhca-J9蛋白是單分子物體,具有6個葉綠體a配體,是一個典型的捕光復合物I葉綠素a/b結合蛋白(圖6)。
2.3 分子對接
分子對接的計算結果顯示(圖 7),羊脂酸與CcLhca-J9蛋白形成氫鍵和p-π的作用力共同影響了兩者之間的結合,其結合能量為-20.9 kJ·mol-1。羊脂酸的羰基氧原子與 Gly68氨基酸殘基形成氫鍵,鍵長為2.9?;Phe67和Phe69中氮原子與羊脂酸的碳氧雙鍵形成p-π,距離均為3.1?;羊脂酸的羥基氫原子與Arg197氨基酸殘基形成氫鍵,鍵長為 2.4?;羊脂酸的羥基氧原子與Arg197氨基酸殘基形成p-π,鍵長為3.3?。
2.4 羊脂酸處理對CcLhca-J9 表達的影響
RT-qPCR結果表明,在羊脂酸處理小飛蓬葉片后0—8 h,CcLhca-J9表達量明顯下降。羊脂酸處理后0.5—1 h,CcLhca-J9的表達量與清水對照相比下降了66.01%;羊脂酸處理后2 h,CcLhca-J9的表達量緩慢下降,與對照相比下降了 82.72%;4—8 h 后,CcLhca-J9的表達量趨于穩定,與對照相比下降 92%—93%(圖8)。羊脂酸處理后,CcLhca-J9表達量與對照相比呈下調趨勢,此結果與前期蛋白質組學結果一致。清水對照和另一有機酸類化合物阿魏酸處理小飛蓬葉片,CcLhca-J9表達量總體未出現顯著變化。
3 討論
植物LHC蛋白家族基因是細胞核內的光合系統基因,其編碼的蛋白與色素形成的復合物能捕獲光能并傳遞能量至光化學反應中心[16]。本研究采用RACE技術克隆獲得小飛蓬葉片LHC蛋白家族基因成員CcLhca-J9cDNA序列,其OFR全長為744 bp,編碼247個氨基酸,分子量為26.766 kD,理論等電點為6.43;二級結構具有螺旋、β轉角、延伸鏈、無規則卷曲;以4y28.1.O(2.80?)為模板進行同源建模,三級結構是單分子物體,具有6個葉綠體a配體,是一個典型的捕光復合物I葉綠素a/b結合蛋白。系統進化分析表明,與除蟲菊和黃花蒿Lhca蛋白進化程度最為接近,同處于菊科這一分支,一致性超過85%,表明該基因家族保守性較強。比對已報道的植物LHC蛋白,發現它們均無信號肽,存在跨膜區,含有結構功能域,為親水性非分泌蛋白[25-26]。
植物LHC表達與抗逆性密切相關。YANG等[27]研究表明,與野生型相比,稀脈浮萍(Lemna perpusilla)的細胞色素b6f突變體在低光強下可維持LHC II在一個恒定的低水平上;李真等[28]研究發現,東南景天(Sedum alfredii)的SaLhcb2在鎘、銅、鉛處理后,根、莖、葉中表達量相比對照均發生顯著上調或者降低;YANG等[29]研究發現,菠菜(Spinacia oleracea)N-端LHCII可識別蛋白降解酶,通過降解LHCII的含量可達到響應不同光強。本研究qPCR結果顯示,羊脂酸脅迫處理小飛蓬葉片后,CcLhca-J9的表達量在處理后0—8 h隨時間延長表現出下降的趨勢。與對照清水和阿魏酸處理相比,羊脂酸抑制CcLhca-J9的表達存在著一定的特異性,預示著LHC可作為抑草靶標用來開發除草劑。
植物LHC是一類潛在的植物源除草劑作用靶標。光合作用抑制劑的開發一直是農藥學家創制新型除草劑的熱門方向,但是建立在充分研究受體蛋白結構與抑制劑相互作用的前提下[30]。去草凈(terburyn)是一種典型三嗪類 PSII抑制劑,與 D1蛋白氨基酸殘基Ile224和Ser233形成氫鍵,與Glu212、Val220和Ile229也有反應[31]。敵草隆(diuron,DCUM)的芳香取代基與 Phe255形成氫鍵,提高了其抑制活性[32]。TeA是一種從鏈格孢菌里分離提取出來的抑制PSII的除草化合物,其作用機制也是與靶蛋白D1的相互作用有關[33]。本研究采用分子對接的手段,發現羊脂酸與植物LHC中氨基酸殘基Gly68、Phe67、Phe69和Arg197形成氫鍵和p-π鍵,其結合能量為-20.9 kJ·mol-1,兩者存在較強的作用力,羊脂酸具有開發成光合作用抑制劑類除草劑的潛力。
4 結論
CcLhca-J9是一個典型的捕光復合物I葉綠素a/b結合蛋白,與羊脂酸結合形成氫鍵和 p-π鍵,同時CcLhca-J9表達量在羊脂酸處理小飛蓬葉片后存在明顯的特異性。因而,推測CcLhca-J9在羊脂酸抑制小飛蓬葉片生長過程中起了較為關鍵的作用,但是該基因是否為植物源羊脂酸的靶標基因有待更進一步的功能驗證。
猜你喜歡
世界農藥(2019年3期)2019-09-10 07:04:10
今日農業(2019年15期)2019-01-03 12:11:33
現代園藝(2017年19期)2018-01-19 02:50:21
長江蔬菜(2016年10期)2016-12-01 03:05:27
獸醫導刊(2016年12期)2016-05-17 03:51:29
現代農業(2016年5期)2016-02-28 18:42:36
雜草學報(2015年2期)2016-01-04 14:58:05
種業導刊(2016年9期)2016-01-03 01:27:14
營銷界(2015年23期)2015-02-28 22:06:18
營銷界(2015年22期)2015-02-28 22:05:11
