花椰菜BraERF023a的克隆及在響應鹽和干旱脅迫中的功能
2021-01-14 03:04:08李慧韓占品賀麗霞楊亞苓尤書燕鄧琳王春國
中國農業科學 2021年1期
關鍵詞:植物
李慧,韓占品,賀麗霞,楊亞苓,尤書燕,鄧琳,王春國
1天津農學院園藝園林學院,天津 300384;2南開大學生命科學學院,天津 300071
0 引言
【研究意義】ERF轉錄因子隸屬于AP2/ERF轉錄因子超家族,在植物響應逆境脅迫中發揮重要作用。花椰菜是重要的蔬菜作物,土壤鹽漬及干旱是造成花椰菜種植減產的主要環境災害。但對花椰菜響應鹽及干旱脅迫的分子基礎認識十分有限,這已成為采用分子育種等手段創制花椰菜抗逆新種質及培育新品種的重要瓶頸。因此,克隆ERF等逆境應答相關轉錄因子或基因并闡釋其功能,對開展抗逆花椰菜分子育種及基因靶向育種具有重要意義。【前人研究進展】植物在生長發育過程中需要不斷應對持續變化的不利環境,如動物啃食、病原體侵染等生物脅迫和干旱、高溫、土壤鹽漬等非生物脅迫。這其中干旱、鹽害及溫度脅迫是影響植物地理分布,造成農作物減產甚至絕收的主要環境災害[1]。因此,闡釋植物響應逆境脅迫的分子基礎及調控機制,進而在作物中開展抗逆分子育種一直是植物學研究的前沿及熱點領域。已有研究表明,WRKY、MYB、bZIP、NAC和AP2/ERF等轉錄因子家族成員在植物生長發育及響應各類逆境脅迫中發揮重要作用[2-3],其中 AP2/ERF轉錄因子家族為植物所特有。該家族成員具有參與 DNA結合的AP2保守結構域,該結構域約由60個氨基酸殘基按照3個β折疊和1個α螺旋的方式構成一個典型三維結構[4-6]。根據 AP2結構域的數量和識別序列的不同,AP2/ERF轉錄因子家族可以分為 AP2、ERF、DREB、RAV和Soloist 5個亞家族[7-8]。其中ERF亞家族只含有1個AP2結構域,被證實主要通過識別并特異性結合各類脅迫響應相關基因順式作用元件,而在植物抵抗各種逆境脅迫過程中發揮重要的調控作用[6,9]。水稻中ERF亞家族成員SNORKEL1(SK1)和SNORKEL2(SK2)可以促進水稻節間伸長,從而使植株可以在深水環境中存活[10]。在擬南芥中過表達菊花CmERF053的研究表明,CmERF053可能參與細胞分裂相關的側枝形成調控途徑,同時在植物抵御干旱脅迫中發揮作用[11]。大豆中GmERF6能夠被干旱誘導表達,將GmERF6在擬南芥中過表達可顯著提高轉化株的抗旱性[12]。在擬南芥中過表達甜薯IbRAP2-12和蕪菁BrERF4同樣也可提高轉化株對鹽和干旱脅迫的耐受性[13-14]。PARK等[15]研究發現擬南芥AtERF71/HRE2功能缺失可使突變株表現為鹽敏感,而將AtERF71/HRE2轉入突變體中,植株表現出耐鹽性,可成功抵抗滲透脅迫,且提高轉化株對低氧脅迫的耐受能力。此外,研究證實,辣椒的CaERFLP1[16]和CaPF1[17],番茄的TERF1[18]和JERF1[19]以及小麥中TaERF1[20]的過表達均可顯著提高轉化株對鹽及低溫等逆境的適應能力。綜上所述,ERF轉錄因子已被證實在植物響應逆境脅迫,特別是非生物逆境脅迫中發揮重要作用。【本研究切入點】雖然在不同植物中已有多個ERF轉錄因子被克隆并證實其在逆境應答中發揮重要作用,但對ERF轉錄因子在花椰菜逆境應答中的功能及調控機制仍知之甚少。筆者實驗室前期對花椰菜中的AP2/ERF家族成員采用高通量轉錄組測序方法進行了分析,從中篩選、鑒定到146個ERF轉錄因子,并證實包括BraERF025a等在內的多個ERF亞家族成員在鹽及干旱脅迫前后呈現顯著差異表達特征[21]。進一步的分子進化樹分析結果表明BraERF023a是與BraERF025a同屬于一個進化分支的ERF亞家族成員,二者具有較近的親緣關系[21]。但BraERF023a是否也響應鹽及干旱脅迫,且其是否在花椰菜抵抗鹽及干旱脅迫中發揮作用仍未知。【擬解決的關鍵問題】對花椰菜中的BraERF023a進行克隆,揭示其序列特征,以及在鹽及干旱脅迫條件下的表達特征,并通過過表達分析闡釋BraERF023a在鹽及干旱脅迫應答中的功能。
1 材料與方法
試驗于 2017—2019年在南開大學生命科學學院植物細胞及分子遺傳學實驗室進行。
1.1 試驗材料
供試花椰菜‘津品 70’種子由天津市科潤蔬菜研究所孫德嶺研究員惠贈。擬南芥為哥倫比亞生態型。克隆載體pEASY-T1購自北京全式金生物技術有限公司。植物表達載體pCAMBIA3301、大腸桿菌DH5α、農桿菌菌株LBA4404由筆者實驗室保存。
1.2 DNA提取
野生型和過表達BraERF023a轉基因擬南芥、花椰菜幼苗新鮮組織,于研缽中加入液氮快速研磨成粉末狀,采用經典CTAB法提取DNA。ddH2O于4℃過夜溶解DNA沉淀。在溶解液中RNase A于37℃金屬浴中反應 2 h除去 RNA。NanoDrop?ND-1000檢測DNA濃度和質量。高質量的DNA于-20℃保存備用。
1.3 RNA提取及反轉錄
參照植物總 RNA提取試劑盒的操作說明提取花椰菜及擬南芥葉片總RNA。NanoDrop?ND-1000檢測提取RNA的濃度,2%瓊脂糖凝膠電泳檢測RNA完整性。采用M-MLV反轉錄酶合成cDNA第一鏈(具體操作參照Promega公司的M-MLV反轉錄說明書進行)。
1.4 花椰菜不同逆境脅迫處理
待花椰菜幼苗長至四片真葉時,將幼苗從營養土中取出,自來水緩緩沖洗根部,注意不要傷害到根系,隨后將幼苗分為兩組,每組至少16個單株。將其中一組幼苗根部直接浸入含有 200 mmol·L-1NaCl的水溶液進行鹽脅迫處理;另外一組幼苗根部浸入含 20% PEG6000的水溶液中進行模擬干旱處理。分別在0、4、8、12和24 h時摘取葉片并進行總RNA提取。提取的RNA反轉錄合成cDNA第一鏈,-80℃保存備用。
1.5 不同脅迫處理條件下BraERF023a表達特征的定量表達分析
采用 qRT-PCR方法對花椰菜不同脅迫處理下BraERF023a的表達特征進行分析。反應體系為20 μL,其中含有10 μL 2×SYBR Green反應mix(羅氏公司)、1 μLBraERF023a定量引物(表 1)。以花椰菜BraActin為內參,采用2-ΔΔCT法進行定量數據分析。
1.6 BraERF023a植物過表達載體構建
以花椰菜cDNA為模板擴增具有NcoI和BstE II酶切位點的BraERF023a編碼區cDNA。將BraERF023a的PCR擴增產物與pEASY-T1克隆載體連接(參照說明書),采用熱激法將重組載體轉化大腸桿菌DH5α。利用NcoI-BraERF023a-F/BstE II-BraERF023a-R引物組合對陽性克隆進行鑒定(表 1),并進一步對陽性克隆進行測序分析。對含有BraERF023a正確序列的陽性克隆及pCAMBIA3301表達載體分別進行NcoI和BstE II雙酶切,回收酶切片段,利用T4 DNA連接酶將BraERF023a連接至 pCAMBIA3301載體的CaMV 35S啟動子下游,篩選獲得 35S::BraERF023a重組表達載體。采用熱激法轉化農桿菌LBA4404,利用BraERF023a特異引物和組合引物(35S-F/BraERF023a-R)對農桿菌陽性轉化菌株進行分子鑒定(表1)。將含有35S::BraERF023a重組質粒的農桿菌采用浸花法遺傳轉化擬南芥。

表1 BraERF023a克隆、載體構建和定量RT-PCR分析引物序列Table 1 Primers used for BraERF023a cloning, vector construction and qRT-PCR assay
1.7 農桿菌介導BraERF023a過表達載體遺傳轉化擬南芥
于 250 mL液體 YEB培養基中,擴大培養含有35S::BraERF023a重組質粒的陽性農桿菌,使其OD600值約為1.2—1.6,5 500 r/min離心10 min收集菌體,用500 mL 5%的蔗糖溶液(含終濃度為0.05%的silwet L-77)重懸菌體,配成轉化液。于盛花期將擬南芥花序浸入轉化液中浸染30 s,用保鮮膜包好植株,平置于托盤中避光培養24 h,7 d后再次轉化,兩次轉化完成后將擬南芥置于培養室中繼續培養至種子成熟,收集種子,記為T0代。
1.8 BraERF023a過表達擬南芥的抗性篩選及鑒定
將春化后的BraERF023a轉基因T0代擬南芥種子種于營養土中,待其長出兩片真葉后,噴灑稀釋10 000倍的 Basta溶液,對轉化株進行抗性篩選。對篩選獲得的T0、T1、T2和T3代抗性轉化株進一步提取其DNA,利用特異引物(BraERF023a-F/BraERF023a-R)和組合引物(35S-F/BraERF023a-R)進行 PCR擴增(表1),檢測BraERF023a是否已整合到擬南芥基因組中。進一步提取 T3代純合抗性轉化株RNA,以qBraERF023a-F/qBraERF023a-R為引物,采用qRT-PCR方法檢測BraERF023a在擬南芥轉化株中的表達特征,收獲純合的T3代轉化株種子用于后續分析。
1.9 BraERF023a過表達擬南芥的表型及抗性分析
將 T3代過表達BraERF023a擬南芥轉化株系和野生型對照種子同時播種,在相同的環境條件下進行培養,并對其株高、莖粗、抽薹時間、分枝數目、果莢長短等表型進行觀察。此外,待過表達BraERF023a擬南芥轉化株系和野生型對照長至3周時,在一次性澆足水后7 d,改為澆灌200 mmol·L-1NaCl溶液進行為期3周的鹽脅迫處理,3周后觀察二者在鹽脅迫下的表型變化。對長至3周的過表達BraERF023a擬南芥轉化株系和野生型對照在一次性澆足水后,停止澆水并進行為期20 d的干旱脅迫處理,之后復水,觀察復水10 d后二者的干旱脅迫表型并統計其存活率。
2 結果
2.1 BraERF023a的克隆及序列分析
分別以花椰菜DNA、cDNA為模板,利用特異性引物組合BraERF023a-F/BraERF023a-R擴增BraERF023a的全長編碼區(表 1)。擴增產物的電泳檢測結果表明,無論是以DNA還是cDNA為模板均能擴增出一條約600 bp的特異條帶,其長度與前期轉錄組測序獲得的BraERF023a序列長度一致,證實花椰菜BraERF023a序列中無內含子。進一步對擴增產物進行TA克隆及測序,測序結果顯示,BraERF023a編碼區全長597 bp,預期編碼一個含有198個氨基酸的蛋白質(圖1)。序列分析發現,BraERF023a包含一個AP2保守結構域,該結構域包含3個β折疊和1個α螺旋,且保守結構域中第14和19位氨基酸分別為丙氨酸和天冬氨酸,均符合ERF轉錄因子的典型結構特征(圖1)。進一步利用ProtFun 2.2 Server(http://www.cbs.dtu.dk/services/ProtFun/)對BraERF023a的蛋白功能進行預測分析。結果顯示,BraERF023a可能主要在植物生長發育及轉錄調控兩個過程中發揮作用。此外,序列比對及聚類分析結果顯示,BraERF023a與B.napus、B. oleracea、B. rapa等十字花科蕓薹屬其他植物中的ERF023序列具有較高相似性(大于90%),而與水稻、玉米、高粱等禾本科植物中的ERF023相似性較低,表明ERF023在十字花科蕓薹屬作物中具有較高保守性,其可能具有相似的功能。
2.2 BraERF023a在鹽脅迫和模擬干旱處理條件下的表達特征
200 mmol·L-1NaCl鹽處理條件下,BraERF023a的表達量隨著處理時間的延長顯著升高,表明鹽脅迫處理可以顯著誘導BraERF023a的表達(圖2-a)。與其相似,在 20%的 PEG6000模擬干旱脅迫處理條件下,花椰菜中的BraERF023a在脅迫處理后不同時間段的表達量均顯著高于處理前(圖 2-b),表明BraERF023a不但正向響應鹽脅迫,也受干旱脅迫的誘導表達。
2.3 BraERF023a表達載體構建及鑒定
分別采用基因特異引物(BraERF023a-F/BraERF023a-R)和組合引物(35S-F/BraERF023a-R)進行菌液PCR,鑒定重組載體是否已成功轉入農桿菌中。隨機挑取6個陽性克隆進行檢測,PCR產物的電泳結果顯示在 6個陽性克隆中均可檢測到長度約為 600 bp和1 100 bp的特異性條帶(圖3-a),條帶大小均符合預期,表明 35S::BraERF023a過表達載體已成功轉入農桿菌中。
2.4 BraERF023a過表達擬南芥轉化株系篩選及分子鑒定
野生型擬南芥于盛花期經農桿菌浸花法侵染后培養至種子成熟,記為T0代,T0代種子萌發后進行Basta抗性篩選。篩選結果顯示,絕大部分幼苗葉片逐漸黃化,26個植株生長發育狀況正常,初步鑒定為抗性轉化株系。隨機選取其中 10個抗性轉化株系分別進行DNA水平及轉錄水平的分子鑒定。以DNA為模板,采用BraERF023a特異引物及BraERF023a與載體序列組合引物分別進行PCR檢測,在其中5個株系中可以分別擴增出與預期大小一致的約600 bp和1 100 bp的特異性條帶,表明BraERF023a已成功整合到這些株系的基因組中(圖3-b)。進一步采用qRT-PCR方法對BraERF023a在這5個陽性株系中的表達特征進行分析。結果顯示,BraERF023a在這 5個株系中的表達水平顯著高于野生型對照(圖 3-c),證實BraERF023a不但成功整合到轉化株基因組內,并且實現在轉化株的高表達。將這5個T0代植株進行多代自交純合,T3代株系用于后續分析。
2.5 BraERF023a過表達轉基因擬南芥表型分析
為揭示BraERF023a在擬南芥中過表達是否影響轉化株的生長發育,對過表達BraERF023a轉基因擬南芥在正常生長條件下不同生長階段的表型特征進行觀察、分析。結果顯示,幼苗生長初期(3周以前),過表達BraERF023a轉基因擬南芥株系與對照相比無明顯差異,但在幼苗生長3周后,過表達BraERF023a轉化株系的長勢明顯優于對照(圖4)。待植株進入成熟期,對其株高、分枝數目等表型特征進行統計分析。結果顯示,過表達BraERF023a轉化株系相對于對照植株表現為植株更高、分枝數目更多、花莖更粗,整體生物量增加。但二者花序大小及果莢長度無明顯差異。在抽薹時間方面,過表達BraERF023a轉化株系比對照滯后約一周(表2)。上述結果表明,在擬南芥中過表達BraERF023a對轉化株的生長發育有一定影響,其可以促進轉化株的生長發育,延長其抽薹期,但對轉化株育性、結種量無顯著影響。
2.6 BraERF023a過表達轉基因擬南芥鹽脅迫分析
BraERF023a過表達轉基因擬南芥和野生型對照經鹽脅迫后生長均受到一定抑制。但對照組擬南芥葉片萎蔫黃化嚴重甚至全部枯萎,植株存活率低;而過表達BraERF023a轉化株系葉片雖也發生黃化,但程度顯著低于對照(圖5),且盡管受到鹽脅迫,BraERF023a過表達轉化株植株最終的存活率(86%)顯著高于對照組(41%)。表明在擬南芥中過表達BraERF023a可以提高轉化株抵御鹽脅迫的能力。
2.7 BraERF023a過表達轉基因擬南芥干旱脅迫分析
干旱脅迫處理20 d后,BraERF023a過表達轉化株系和野生型植株均萎蔫嚴重,但野生型植株基本已全部萎蔫黃化至死亡,而BraERF023a過表達轉化株系萎蔫黃化程度相對野生型較輕,部分轉化株系葉片仍保持綠色(圖6)。對所有干旱處理植株進行復水,復水10 d后,其中2個過表達BraERF023a轉化株系中(L8、L9)的大部分植株恢復生長,而對照組絕大部分已枯萎(圖 6)。且過表達BraERF023a轉化株系的存活率顯著高于對照組(圖 7)。表明在擬南芥中過表達BraERF023a可以顯著提高轉化株的耐旱性。
3 討論
3.1 BraERF023a在植物中的分子進化特征分析
AP2/ERF轉錄因子最初在植物中被發現。1994年 JOFUKU等[4]從模式植物擬南芥中分離到第一個AP2轉錄因子APETALA2,該轉錄因子與花發育和種子萌發有關。隨后AP2/ERF轉錄因子家族成員陸續從不同植物中被鑒定、克隆,并證實其為植物所特有。基因結構分析表明AP2/ERF家族成員均含有保守的AP2結構域[5]。根據AP2結構域的數目及序列特征,AP2/ERF家族中的ERF亞族和DREB亞族均含有單個的AP2結構域[6],但是AP2結構域第14位和第19位氨基酸殘基在兩者中不同。ERF亞族成員AP2結構域的第14位和19位分別為丙氨酸和天冬氨酸,而DREB亞族成員該結構域第14位和19位分別為纈氨酸和谷氨酸[7]。通過對BraERF023a的序列分析發現,其具有1個AP2結構域,并且保守結構域第14位和19位分別為丙氨酸和天冬氨酸,符合ERF亞族成員的典型結構特征,證實其為ERF轉錄因子。已有研究證實ERF轉錄因子可與GCC盒(AGCCGCC)等下游靶基因的順式作用元件結合,從而在基因轉錄調控中發揮作用[9,22]。進一步的序列比對發現,BraERF023a與雙子葉植物,特別是與花椰菜同屬于十字花科蕓薹屬的其他植物中的ERF023在進化上具有較高保守性,而與單子葉植物中該轉錄因子的序列同源性較低,暗示BraERF023a在祖先植物進化形成單子葉和雙子葉植物時,序列已發生分化,但其功能是否也發生了亞功能化,有待進一步探究。在克隆并揭示BraERF023a序列及表達特征基礎上,通過遺傳轉化策略將其在花椰菜中進行過表達,并分析過表達BraERF023a花椰菜轉化株系生長發育及在逆境脅迫條件下的表型及生理特征,對于闡釋BraERF023a的功能具有重要價值。但花椰菜遺傳轉化效率受材料基因型、遺傳背景等因素影響很大,不同基因型材料遺傳轉化效率普遍不高,很難在短期內獲得轉化株系,即使獲得了轉化株系,很多轉化株系也不能正常結籽。因此,轉化效率低及結籽困難一直是花椰菜基礎研究進展緩慢的一個重要瓶頸。基于此,本研究為探究花椰菜BraERF023a的功能,首先將其轉入到易于轉化的模式植物擬南芥中進行功能分析。BraERF023a在花椰菜中過表達及采用CRISPR/Cas9基因編輯技術敲除的工作正在進行中。
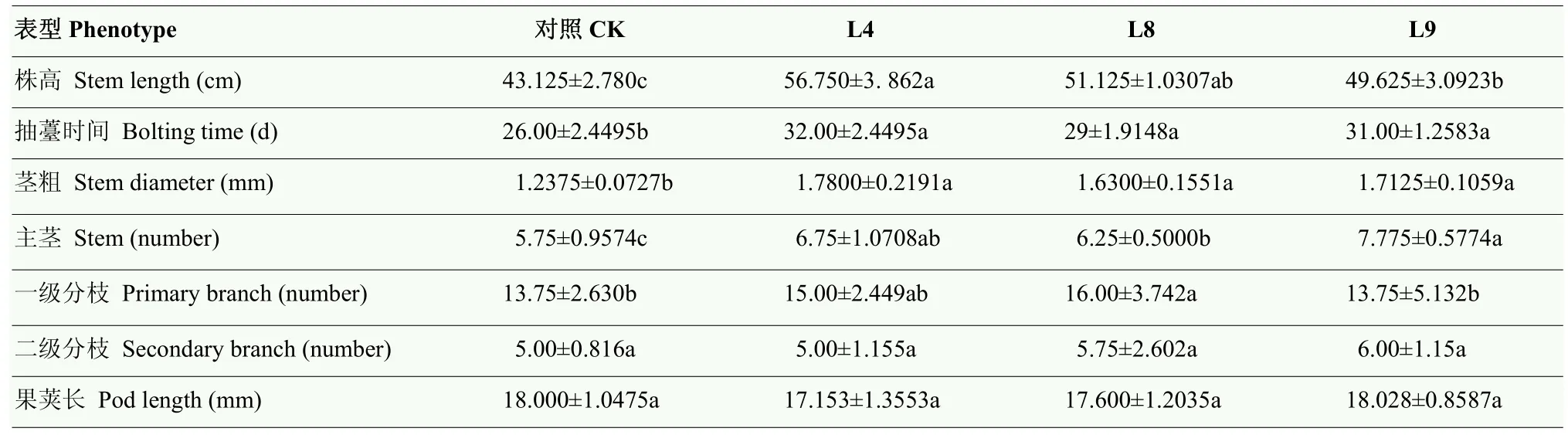
表2 過表達BraERF023a轉基因擬南芥表型分析Table 2 Phenotypic analysis of overexpression BraERF023a transgenic Arabidopsis
3.2 BraERF023a響應鹽和干旱脅迫分析
已有研究表明,ERF轉錄因子亞家族成員主要在植物逆境響應中發揮作用。但不同植物來源的ERF成員或同一植物中不同的ERF成員對逆境脅迫的響應特征不盡相同。JERF1在煙草中過表達可增強轉化株對低溫及高鹽的耐受[23]。擬南芥AtERF1的表達受ABA負調控,其過表達導致ABA積累和脯氨酸增加,進而增強轉化株對干旱、高鹽和熱脅迫的耐受性[24]。過量表達MdERF72的蘋果愈傷組織在高鹽和低溫脅迫處理下,生長勢明顯強于野生型對照,表明MdERF72在響應高鹽、低溫脅迫過程中發揮著重要的正調控作用[25]。苜蓿中 ERF轉錄因子MfERF1過表達可促進轉化株多胺的合成與轉化,提高植株抗氧化能力和脯氨酸積累,進而提高轉化株對低溫的耐受[26]。BOLT等[27]研究發現ERF105在擬南芥對凍害的耐受性中發揮重要作用,是擬南芥冷馴化中所必需的轉錄調控因子。ERF102和ERF103也被證實在擬南芥冷馴化中扮演重要角色[28]。有關花椰菜中ERF轉錄因子的報道很少。筆者課題組前期的研究表明,在花椰菜中至少存在146個ERF成員,這其中絕大部分成員的功能未知[21]。本研究結果表明,鹽及干旱逆境脅迫可顯著誘導花椰菜中BraERF023a的表達,暗示BraERF023a是一個鹽及干旱正向響應應答因子。進一步的過表達功能分析表明,在擬南芥中過表達BraERF023a可顯著提高轉化株對鹽及干旱脅迫的耐受。該結果進一步證實,BraERF023a在植物耐鹽及抗旱應答中發揮重要的正向調控作用。此外,前期研究結果表明花椰菜中除BraERF023a外,還具有一個BraERF023a的等位變異,命名為BraERF023b,其編碼區長度為567 bp,比BraERF023a少30 bp[21]。在花椰菜中BraERF023b是功能性冗余還是具有其他功能有待進一步探究。目前,有關不同植物中ERF023的研究很少。在杏桃干旱脅迫響應的研究中發現,干旱脅迫可誘導其內PpERF023的表達,暗示PpERF023在干旱脅迫應答中發揮重要作用[29]。杏桃ERF023在擬南芥中的同源基因AtHARDY被證實在提高植株的水分利用效率,提高對干旱脅迫的適應能力過程中發揮重要作用[30]。這暗示不同植物來源的ERF023在干旱脅迫應答中可能發揮重要作用,但其是否也都參與鹽脅迫應答并發揮作用有待進一步探究。在葡萄中的研究表明,ERF023可能參與葡萄對病原菌的防御,其過表達在提高葡萄對灰霉病的抗性方面有一定作用[31]。暗示ERF023不但在非生物逆境脅迫中發揮作用,也響應生物脅迫。因此,進一步克隆不同植物中的ERF023,并揭示其對不同逆境脅迫響應的分子機制,對于闡釋ERF類轉錄因子的功能具有重要意義。
3.3 BraERF023a在生長發育中的功能分析
盡管大部分研究表明ERF類轉錄因子主要在植物響應逆境脅迫中發揮作用。但也有研究表明一些ERF轉錄因子參與植物生長發育調控。如擬南芥中ERF轉錄因子家族成員TINY可抑制植物激素BR調控的生長發育過程,同時也調控植株對干旱的脅迫反應。TINY過表達可導致植物生長發育遲緩[32]。在水稻中ERF轉錄因子OsEATB可通過下調赤霉素生物合成基因限制節間伸長,導致植株矮化[33]。NAKANO等[34]研究表明,番茄SlERF52在花梗離區特異表達,其在花梗脫落中發揮重要作用。過量表達擬南芥AtERF019不但可以提高植物的耐旱性,還可以延遲植株生長和衰老[35]。在黃瓜中鑒定出 138個ERF家族成員,其中多個成員被認為在雌花發育過程中發揮調控作用[36]。本研究結果表明,正常生長條件下,在擬南芥中過表達BraERF023a可使轉化株表現出顯著不同于野生型對照的生長發育特征,如相對于野生型對照,過表達BraERF023a轉基因擬南芥植株更高,主莖加粗,蓮座和一級分枝數增多、生物量提高等,并且過表達轉化株的抽薹時間延長,致使其生長期延長(表2、圖4)。該結果表明在擬南芥中過表達BraERF023a可以促進轉化株的生長發育,暗示BraERF023a不但是一個鹽及干旱脅迫響應的正向調控因子,其在植物生長發育調控過程中也可能發揮重要作用,有待對其調控機制進行深入探究。
4 結論
花椰菜BraERF023a含有一個保守的AP2結構域,其與十字花科蕓薹屬植物中的ERF023具有較高序列同源性。轉錄表達特征分析證實鹽及干旱脅迫可顯著誘導BraERF023a的表達。在擬南芥中過表達BraERF023a可促進轉化株的生長發育,并顯著提高轉化株對鹽及干旱脅迫的耐受力。結果證實,BraERF023a在植物響應鹽及干旱脅迫中發揮重要的正向調控作用,是一個優質耐鹽抗旱基因,在花椰菜抗逆分子育種中具有重要應用價值。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
