煙草中異源表達譖茄HQT基因對多酚物質合成的影響
2021-01-18 06:22:01丁香玉王鵬偉儲昭輝趙翔宇丁新華李洋
中國煙草科學 2021年2期
關鍵詞:煙草
丁香玉 王鵬偉 儲昭輝 趙翔宇 丁新華 李洋
摘要:羥基肉桂酰輔酶A:奎尼酸羥基肉桂酰轉移酶Iydroxycinnamoyl-COA:quinate hydroxycinnamoyltransferase,HQn)是催化茄科植物咖啡酰奎尼酸生物合成最后一步的關鍵酶。為培育富含多酚物質的煙草,提高煙草品質,通過克隆番茄FQT基因,構建P35:SHOT超量表達載體,在野生型三生煙草(SS)中異源表達,利用高效液相色譜技術(HPLC)及液質聯用(ILC-MSMS)技術對轉基因煙草葉片中一咖啡酰奎尼酸及黃酮醇含量進行檢測,探究SHQT基因對三生煙草一咖啡酰奎尼酸及黃酮醇物質合成的影響。結果顯示,轉SQT煙草葉片中一咖啡酰奎尼酸總含量最高可提高14.98倍,蘆丁和山奈酚蕓香糖苷含量最高可分別提高1.47和9.32倍,且生長表型與野生型煙草沒有明顯差異;轉SIHOR基因煙草葉片中有4種(1-CQA,3-CQA,4-CQA,5-CQA)一咖啡酰奎尼酸,野生型煙草葉片中只檢測到3種(3-CQA,4-CQA,5-CQA)SIHOT基因在煙草中的異源表達促進了1-CQA的生物合成且提高了一咖啡酰奎尼酸總含量,同時增加了黃酮醇的生物合成。
關鍵詞:煙草;番茄OT基因;一咖啡酰奎尼酸;1-CQA;黃酮醇
酚類物質是煙草次生代謝的主要產物,與煙草品質緊密相關,對煙草的生長發育、抗逆性、色澤、香氣質等方面有重要影響。其中,一咖啡酰奎尼酸和黃酮醇是煙草中重要的多酚物質,二者含量可達總酚量的80%以上。
一咖啡酰奎尼酸是煙草、番茄、馬鈴薯、蘋果等植物中重要的苯丙酸,能夠幫助植物細胞抵御低溫、紫外線等各種非生物脅迫以及病原、害蟲侵害5。根據咖啡酰基在奎尼酸上的結合位置不同可分為4種異構體:1CQA、3-CQA、4-CQA和5-CQA(CGA,綠原酸)。在植物生長發育過程中,CGA能夠通過增強植物清除活性氧的能力,幫助植物抵御逆境。黃酮醇作為植物重要的苯丙烷類物質,廣泛存在于煙草、番茄、馬鈴薯等茄科作物中,黃酮醇類物質參與植物生長發育的許多方面,如植物與微生物互作、紫外線防護、花粉生長和抵御害蟲侵害等。
羥基肉桂酰輔酶A:奎尼酸羥基肉桂酰轉移酶HQT,是多種作物咖啡酰奎尼酸合成的最后一步關鍵酶,將咖啡酰輔酶A和奎寧酸催化形成綠原酸。在蒲公英中超量表達Tahor可使葉片CGA水平升高82.49%,而RNA干擾使葉片CGA水平降低51.48%;超量表達LMHOT使忍冬中CGA含量提高2-3倍川;RAJA在馬鈴薯中通過RNAi對HOT的抑制導致CGA下降909%以上;RICARDAL剛將HOT在番茄中超量表達使CGA含量提高10%,RNAi沉默HQT后番茄葉片中CGA含量降低98%;結果均表明HOT基因是CGA合成的關鍵基因且正向調控綠原酸的生物合成。SHOT基因在番茄中調控綠原酸的合成,但對蘆丁和山奈酚蕓香糖苷的合成調控功能尚不清晰。因此,本研究以番茄中CGA合成的關鍵酶SOT為對象,分析番茄SIHOT基因在煙草中異源表達對煙草咖啡酰奎尼酸和黃酮醇生物合成的影響,為增加煙草中次生代謝物質含量,提高煙草品質及抗逆能力提供參考。
1材料與方法
1.1材料
番茄植株于山東農業大學溫室大棚種植(2019年5-8月),生長條件為:白天25~28℃,夜晚15~18℃,土壤相對濕度為(7010)%。生長至果實成熟期,摘取果實,取果皮液氮急速冷凍,-80℃超低溫冰箱保存備用。
三生煙草(SS)于山東農業大學作物生物學國家重點實驗室植物培養室種植(T0代轉基因株系及野生型種植于2019年10-12月,T1代轉基因株系及野生型種植于2020年2-4月),生長條件為:25℃,16h8h光暗周期,相對濕度為(70:10)%。番茄品種Micro om種子,三生煙草(SS)種子,大腸桿菌EscherichiacoliDH5a,農桿菌LBA4404和真核表達載體PCXSN均由本實驗室保存提供。
1.2方法
1.2.1SOT基因克隆和表達載體構建采用
Plant RNA Kit(OMEGA,美國)提取番茄果皮總RNA,根據反轉錄試劑盒(TOYOBO,日本),以番茄果皮總RNA為模板反轉錄獲得CDNA,置于20℃保存。
以上述反應得到的cDNA為模板,根據番茄HOT基因序列(Genebank AJ582652),設計引物,序列為SOTF:5-ATGGGAAGTGAAAAAATGA-3,SIHOT-R:5-AATTTGTGTTGTACATTCTTGAT-3'。以cDNA為模板,利用高保真KOD酶(TOYOBO,日本)進行片段擴增,擴增到目的條帶后切膠回收。回收純化后SHOT基因片段和實驗室保存的表達載體PCXSN,用XcmI(NEB,美國)分別進行酶切,純化后將載體和目的基因片段用T4DNA連接酶(ThermoFisherScientific,美國)進行連接,構建遺傳轉化載體P35S:SHQ7。重組載體熱激轉入大腸桿菌DHSa,以35s啟動子內部序列設計的引物35Ss-F:5-ACGCACAAICCCACTATCCTT-3'和基因內部序列設計的引物SIHOT-R5-AATGTGTGTACATCTGA-3,挑取單克隆進行菌落PCR檢測,選擇陽性克隆過夜擴繁,提取質粒送至華大基因股份有限公司(深圳)測序。經測序無誤后,轉化農桿菌LBA4404。
1.2.2農桿菌介導的煙草遺傳轉化及陽性檢測選取煙草葉片作為外植體,采用農杄菌介導的葉盤轉化法叫,將重組載體P35S:ST轉化三生煙草受體材料。采用CTABI]法提取轉基因植株葉片基因組DNA,進行PCR陽性檢測。待生根培養基中煙草根系發育良好時,移栽到營養缽中于培養室生長。在T0代陽性植株中隨機選擇3個株系,編號為SOT-1,SHQT-2和SHOT-3,收取種子后進行T1代種植。每個株系取3棵生物學重復,進行半定量檢測,驗證遺傳穩定性。
1.2.3一咖啡酰奎尼酸和黃酮醇物質提取和含量測定待煙草葉片生長至8葉期,取中部3片真葉進行檢測。參照文獻[1-18],對轉基因和非轉基因煙草葉片進行含量檢測和質譜分析,取10L過濾液用于HPLC檢測,5uL用于LC-MSMS分析標準品為蘆丁(Sigma),一咖啡酰奎尼酸(Sigma),山奈酚蕓香糖苷(Extrasynthese)。T0代煙草每株重復檢測3次;T1代煙草每個株系3個生物學重復。
2結果
2.1ST基因克隆和基因表達載體構建
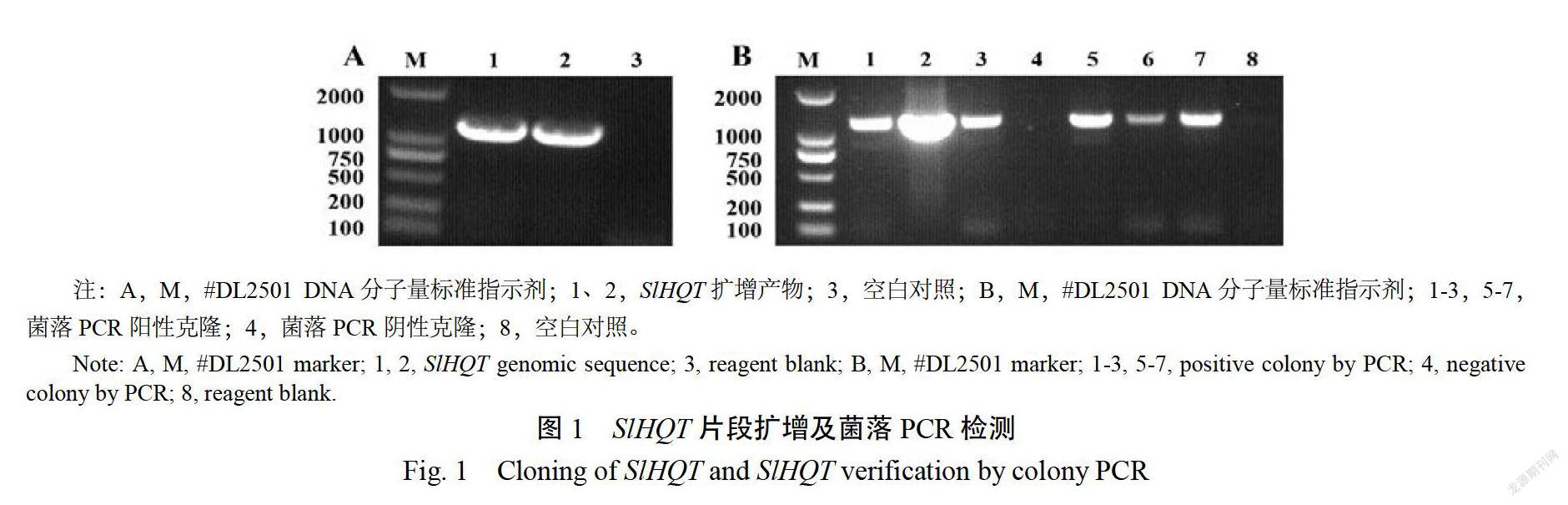
按照番茄中SOT的已知序列設計引物,以番茄果皮cDNA為模板,通過PCR擴增獲得長度為1293bp的清晰目標序列(圖1A);構建P35:SHOT,挑取單克隆利用35-F、SIHOT-R引物進行PCR陽性驗證(圖1B),選取3個陽性克隆過夜擴繁后提質粒測序。
將測序結果與目標序列Sihor(GenebankAJ582652)比對,二者序列比對一致,確認成功克隆SHOT的完整基因序列,并成功構建P3S:SHOT載體。
2.2SQT基因遺傳轉化和轉基因煙草的獲得
對轉基因煙草進行陽性檢測(圖2),在獲得的9株轉基因材料中,有7株擴增到了目標基因序列,確認為陽性轉基因植株,轉化率為77.7%。
2.3煙草葉片一咖啡酰奎尼酸和黃酮醇物質含量
HIPLC檢測結果顯示(表1、圖3),轉SIHT基因的煙草葉片中一咖啡酰奎尼酸、蘆丁、山奈酚蕓香糖苷的含量都有不同程度的提高。其中,一咖啡酰奎尼酸的含量提高了797~14.98倍,最高含量達4598.17ugg。蘆丁和山奈酚蕓香糖苷含量最高分別可達265.08和175ugg,分別是相同條件下野生型三生煙草(SS)葉片中含量的9.58倍和8.71倍。
對T1代植株進行半定量檢測,結果表明(圖4)轉基因煙草中SFQT基因表達量較高,而野生型中沒有基因表達,說明轉基因陽性的真實性和遺傳穩定性。并且轉基因煙草T1代植株3種物質含量都有不同程度的提高,說明轉SHOT基因煙草富含一咖啡跣奎尼酸及黃酮醇的優勢可以穩定遺傳(圖5、表2)。
2.4煙草葉片中一咖啡酰奎尼酸質譜分析
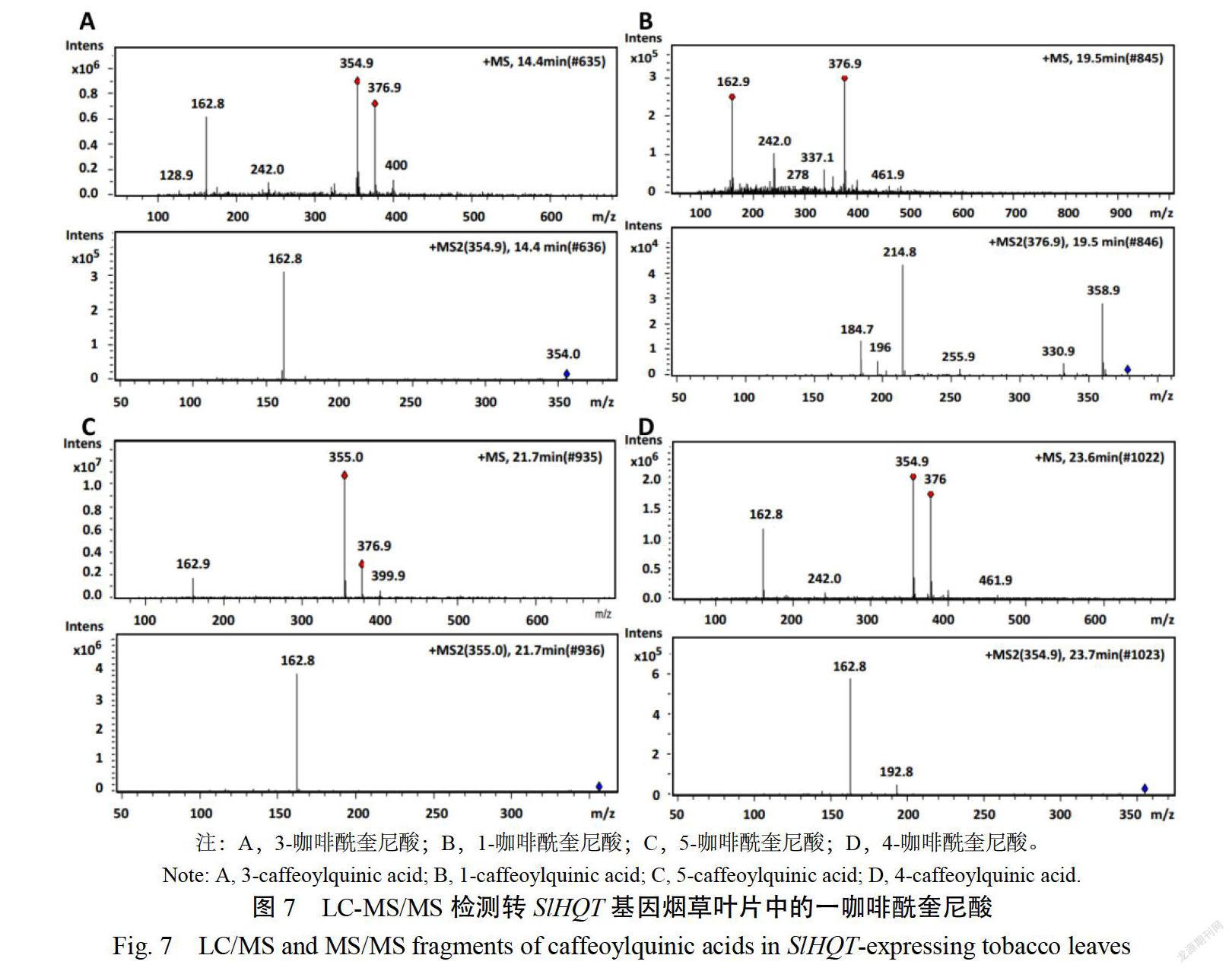
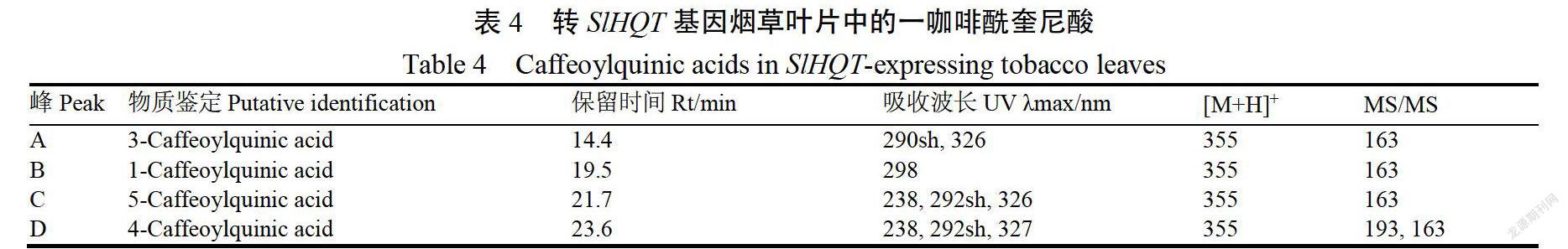
質譜分析結果顯示(圖6,表3),在野生型煙草葉片中,檢測到3-CQA,4-CQA,5-CQA三種咖啡酰奎尼酸,而在轉SIHOT基因煙草葉片中檢測到1-CQA,3-CQA,4CQA,5-CQA四種一咖啡酰奎尼酸(圖7,表4)。
3討論
本研究將番茄中的QT基因連接35s啟動子構建真核表達載體,在野生煙草中異源表達,煙草葉片中一咖啡酰奎尼酸含量明顯提高,說明番茄中的HT基因能在煙草中正向調控一咖啡酰奎尼酸的生物合成,提高一咖啡酰奎尼酸含量,這與RICARDA1、ANDREA等的研究結果基本一致。番茄SHQT與煙草MHT編碼的蛋白質序列相似性分別高達91%,在植物體內行使相同的功能,都是將咖啡酰輔嗨A和奎寧酸催化形成綠原酸,正向調控綠原酸的生物合成。
另外,轉SIOT基因煙草與野生型相比,多檢測到一種一咖啡酰奎尼酸(1CQA),在番茄中存在4種一咖啡酰奎尼酸,而在野生型煙草中僅檢測到3種,可能與SHQT和MHOT編碼的蛋白序列差異有關,后者在225-229位置缺少蘇氨酸(T)、絲氨酸(S)、脯氨酸(P)、賴氨酸(K)、脯氨酸(P)5個氨基酸,導致咖啡酰基在奎尼酸的1號位不能結合,不能形成1-CQA;也可能是因為野生型煙草葉片中1-CQA含量極低,未達到檢測線,而轉基因煙草葉片中,SIHOT基因的異源表達促進一咖啡酰奎尼酸的大量合成,1-CQA含量升高,達到檢測線。而本研究中高效液相色譜技術未分離出4種咖啡酰奎尼酸的單峰,可能與流動相配比和酸濃度有關。
SHHOT基因在番茄中調控綠原酸的合成,并且隨著果實成熟,表達量提高,但對蘆丁和山奈酚蕓香糖苷的合成調控功能還未有明確報道。此次試驗將番茄中HOT基因在三生煙草上表達分析,對T0代和T1代轉基因煙草葉片進行一咖啡酰奎尼酸和黃酮醇含量檢測,發現轉基因煙草代謝物質含量增多的表型能夠穩定遺傳,但是含量差異較大,猜測煙草的生長狀態對次生代謝物質積累具有較大影響。除一咖啡酰奎尼酸外,煙草中蘆丁和山奈酚蕓香糖苷含量也有明顯提高。這幾種多酚物質合成路徑共享多種中間酶和前期次生代謝物,都是以苯丙氨酸為合成底物,經過PAIL(L-苯丙氨酸解氨酶)C4H(肉桂酸羥化酶)、4CL(p-香豆酰CoA合成酶)催化合成p-香豆酸輔酶A。然后以p-香豆酰輔酶A為底物,通過多種酶分別催化合成一咖啡酰奎尼酸,蘆丁和山奈酚蕓香糖苷等。結合試驗結果,推測HT基因的超量表達可能激活一咖啡酰奎尼酸合成路徑中上游的PAL、C4H、4CL酶,增加前期代謝物質萃丙氨酸和p-香豆酸輔酶A的積累,這些合成相關酶及代謝物質的增多,同時促進下游多種黃酮醇合成酶的反應,從而更強地激活蘆丁和山奈酚蕓香糖苷合成路徑,而這些黃酮醇合成途徑中關鍵酶基因的表達變化情況有待進一步研究。
4結論
將SOT在煙草中異源表達發現,HQT基因正向調控煙草中一咖啡酰奎尼酸的合成,也促進煙草中蘆丁和山奈酚蕓香糖苷的積累。轉SHOT基因煙草葉片中一咖啡酰奎尼酸種類比野生型煙草葉片中多一種(1-CQA),且總含量最高可提高14.98倍,達4598.17ugg,蘆丁和山奈酚蕓香糖苷含量最高可分別提高11.47和9.32倍。轉SHQT基因沒有影響植株的正常生長發育。這幾種多酚物質含量提高有助于提高煙草抵抗脅迫能力,增加香氣量,提高煙草品質,為煙草品質育種和多用途開發利用提供研究基礎。
參考文獻
[1]朱小茜,徐曉燕,黃義德,等.多酚類物質對煙草品質的影響安徽農業科學,2005,33(10):132-133
[2]楊虹琦,周冀衡,邵巖,等.不同產地烤煙葉中綠原酸和蕓香苷的含量分析.天然產物研究與開發,2006,18(4):670-673
[3] CLIFFORD M N. Chlorogenic acids and other cinnamates-nature, occurrence, dietary burden, absorption and metabolism [J] Journal ofthe Science of Food &Agriculture, 2000, 80(7): 1033-1043
[4] UPADHYAY R, MOHAN R L J An outlook on chlorogenic acids occurrence, chemistry, technology and biological activities [J].Critical Reviews in Food Science and Nutrition. 2013. 53(9968-984.
[5]王玲娜,姚佳歡,馬超美.綠原酸的研究進展[J].食品與生物技術學報,2017,36(11):1121-1130
[6] CLE C, HILL L M, NIGGEWEG R, et al. Modulation of chlorogenic acid biosynthesis in Solanum lycopersicum: consequences forphenolic accumulation and UV tolerance [J]. Phytochemistry, 200869(11):2149-56
[7] LI Y, TANG W Z, CHEN J, et al. Development of marker-free transgenic potato tubers enriched in caffeoyl quinic acids andflavonols [J]. Journal of Agricultural and Food Chemistry, 2016,64(14):2932-2940
[8] GRACE S C, LOGAN B A. Energy dissipation and radical scavengng by the plant phenylpropanoid pathway[J]. Biological sclences,2000,355(1402):1499-1510
[9] WANG S L, CHU Z H, JIA R, et al. SIMYB12 regulates flavonol synthesis in three different cherry tomato varieties [J]. Scientifi Reports,2018,8(1):1582
[10] LIU Q, LIU Y, XU Y, et al. Overexpression of and RNA interference with hydroxycinnamoyl-coa quinate hydroxycinnamoyl transferaseaffect the chlorogenic acid metabolic pathway andenhance saaittolerance in Taraxacum antumngense Kitag J]. Phytochemistry Letters2018,(28):116-123
[11]CHEN Z, LIU G, LIU Y, et al. Overexpression of the IMHO/ gene increases chlorogenic acid production in Lonicera macranthoides Hand-mazz[J]. Acta Physiologiae Plantarum, 2017, 39(1): 27
[12] RAJA S P, ROSHANI S, VENKATESAN G S, et al. Synthesis and regulation of chlorogenic acid in potato: Rerouting phenylpropanoidflux in Hqt-silenced lines[J]. Plant Biotechnology Journal, 201513(4):551-564.
[13] NIGGEWEG R, MICHAEL A J, MARTIN C Engineering plants with increased levels of the antioxidant chlorogenic acid [J]. Nature Biotechnology, 2004, 22(6): 746-754
[14趙莉,鐘鳴,馬慧,等,農桿菌介導的煙草高效遺傳轉化體系的建立.江蘇農業科學,2011,39(3):67-68
[15] POREBSKI S, BAILEY L G, BAUM B R Modification of CTAB DNA extraction protocol for plants containing high polysaccharideand polyphenol components [J]. Plant Molecular Biology Reporter1997,15(1):8-15
[16] LI Y, CHEN M, WANG S L, et al. ATMYBLL regulates caffeoylquinic acid and flavonol synthesis in tomato and tobacco [J]Plant Cell Tissue and Organ Culture, 2015, 122(2): 309-319
[17] LUO J, BUTELLI E, HILL L, et al. ATMYB12 regulates caffeoylquinic acid and flavonol synthesis in tomato: expression irfruit results in very high levels of both types of polyphenol [J].The Plant Joumal,2008,56(2):316-326
[18] CLIFFORD M N, JOHNSTON K L, KNIGHT S, et al. Hierarchical scheme for LC-MSN identification of chlorogenic acids [J] Journal of Agricultural and Food Chemistry. 2003.51: 2900-2911
[19] MOGLIA A, LANTERI S, COMINO C, et al. Dual catalytic activity of hydroxycinnamoyl-coenzyme a quinate transferase from tomatoallows it to moonlight in the synthesis of both mono-anddicaffeoylquinic acids [J]. Plant Physiology, 2014, 166(4177-1787
[20] MISRA P, PANDEY A, TIWARI M, et al. Modulation of transcriptome and metabolome of tobacco by Arabidopsistranscription factor, ATMYB12, leads to insect resistance [J]. Plant Physiology,2010,152(4):2258-2268
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中國煙草學報(2019年5期)2019-11-14 07:54:12
首都公共衛生(2019年5期)2019-05-21 01:08:34
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
新聞傳播(2016年3期)2016-07-12 12:55:34
當代化工研究(2016年9期)2016-03-20 16:22:08
自動化博覽(2014年6期)2014-02-28 22:32:15
聲屏世界(2014年6期)2014-02-28 15:18:09
西南學林(2013年2期)2013-11-12 12:58:54
中國煙草學報(2012年5期)2012-04-12 06:21:18
