植物非組蛋白賴氨酸乙酰化修飾的蛋白質組學研究進展
2021-01-22 09:08:56鄭璐沈仁芳蘭平
生物技術通報 2021年1期
關鍵詞:植物
鄭璐 沈仁芳,2 蘭平,2
(1. 中國科學院南京土壤研究所 土壤與農業可持續發展國家重點實驗室,南京 210008;2. 中國科學院大學,北京 100049)
蛋白質賴氨酸乙酰化(Nε-lysine acetylation)是一種在原核生物和真核生物都廣泛存在的重要的蛋白質翻譯后修飾(Post-translational modifications,PTMs)[1-4]。賴氨酸乙酰化酶(Lysine acetyltransferases)催化乙酰基轉移到賴氨酸的ε-氨基側鏈形成蛋白質的乙酰化修飾,同時這一反應可以被賴氨酸去乙酰化酶(Lysine deacetylases)逆轉。植物線粒體中一些賴氨酸乙酰化修飾也可以通過非酶促反應發生[5]。
賴氨酸乙酰化包括組蛋白和非組蛋白的乙酰化修飾。1964年,Allfrey等[6]在牛胸腺細胞核染色體的組蛋白上首次發現了賴氨酸乙酰化修飾。真核生物染色體由核心組蛋白八聚體(H2A、H2B、H3和H4)和其纏繞的146 bp的DNA組成了基本單位核小體[7]。組蛋白賴氨酸乙酰化修飾主要發生在核心組蛋白的N末端,是影響染色體結構的重要翻譯后修飾。賴氨酸乙酰化有利于組蛋白和DNA八聚體的解離,核小體松弛,從而使各種轉錄因子能夠與DNA結合位點特異性結合,控制基因的表達水平[8]。大量研究表明,組蛋白通過乙酰化修飾可逆而快速的變化在植物生長發育和非生物/生物脅迫響應中發揮著重要的調控作用[8-10]。
1997年,Gu和Roeder[11]在 人 腫 瘤 抑 制 因子p53上首次發現了乙酰化修飾參與非組蛋白的轉錄調控。隨后,基于乙酰化肽段的免疫親和純化(Acetylpeptides immune affinity purification)和液相色譜-串聯質譜(Liquid chromatography - tandem mass spectrometry,LC-MS/MS)的乙酰化蛋白質組(Acetylproteome)技術不斷發展[12],大量非組蛋白賴氨酸乙酰化修飾被發現。2006年,Kim等[13]首次報道了HeLa細胞和小鼠肝臟線粒體的乙酰化蛋白質組,一共鑒定了195個乙酰化蛋白質,且大部分為非組蛋白。隨后,Lundby等[14]在小鼠的不同組織中一共鑒定到4541個乙酰化蛋白質(15474個修飾位點),主要參與基因表達、蛋白質代謝、三羧酸(Tricarboxylic acid,TCA)循環和細胞凋零等代謝過程。Zhang等[4]在原核生物大腸桿菌(Escherichia coli)中也鑒定到了91種乙酰化蛋白質,主要為代謝酶和調節因子。這些都表明賴氨酸乙酰化修飾在非組蛋白中大量存在,且這種翻譯后修飾的調控方式從細菌到哺乳動物在生物進化上是相對保守的。非組蛋白的乙酰化修飾通過調控酶活力[15-16]、蛋白質相互作用[17]和蛋白質穩定性[18]等方式調控各種生物學過程。目前,隨著乙酰化蛋白質組學技術的普及,植物中普遍存在的大量非組蛋白賴氨酸乙酰化修飾被發現。本文綜述了植物中非組蛋白賴氨酸乙酰化修飾的蛋白質組學研究進展,闡述其特征以及在植物生長發育和環境脅迫中的響應和作用,旨在為乙酰化修飾介導的調控機理和相關實際應用奠定理論基礎。
1 植物乙酰化蛋白質組學技術的發展
過去大量研究局限在植物組蛋白的乙酰化修飾調控。近十年來,植物的非組蛋白乙酰化修飾越來越受到研究者的關注。乙酰化蛋白質組學在多種植物中展開,主要集中在模式植物擬南芥和糧食作物水稻和小麥,還包括一些經濟作物大豆、茶樹和草莓等(表1)。隨著乙酰化蛋白質組學技術的日益成熟和發展,植物乙酰化蛋白質組學鑒定到的非組蛋白乙酰化蛋白質數量從不足一百增加到幾千,并且從定性的靜態分布研究拓展到了不同時空下的乙酰化的定量動態變化,極大加深了對植物非組蛋白賴氨酸乙酰化修飾的時空變化特征和潛在的生物學功能的認知。
目前,植物乙酰化蛋白質組研究大多采用乙酰化肽段的特異性抗體親和富集技術結合LC-MS/MS進行分析(表1)。該方法的基本流程如圖1所示。植物樣品一般采用TCA-丙酮法提取總蛋白,提取的蛋白質采用過濾輔助樣品制備法(Filter-aided sample preparation,FASP)進行胰蛋白酶酶解成肽段,然后其中的乙酰化肽段采用特異性抗體樹脂進行富集。富集的乙酰化肽段脫鹽后進行LC-MS/MS質譜分析以及蛋白質定性和定量分析。最后進行生物信息學分析,包括蛋白質功能富集和蛋白質互作網絡分析等。與常規蛋白質組學相比,乙酰化蛋白質組學還需要對乙酰化修飾位點的基序進行分析。一般對乙酰化位點上下游10個氨基酸進行肽段序列分析,解析其保守肽段特征。
早期的植物蛋白乙酰化蛋白質組學研究雖然也采用了賴氨酸乙酰化抗體的免疫親和層析富集技術,但是受質譜分辨率和精密度等因素影響[42-43],鑒定到的乙酰化蛋白質數量相對較少[19,20,24]。2011年,Finkemeier等[19]在擬南芥不同器官中只鑒定到74個乙酰化蛋白質和91個乙酰化位點。2014年,Nallamilli等[24]在水稻懸浮細胞中只鑒定到40個乙酰化蛋白質和66個乙酰化位點。但是,這些研究初步表明了乙酰化修飾不僅存在于組蛋白,而且廣泛存在于植物不同組織的非組蛋白,參與多種代謝過程。近年來,通過高效的固定化抗乙酰化賴氨酸抗體(Anti-acetyllysine antibodies)親和純化技術來富集乙酰化肽段,增加了低豐度肽段的檢出率,同時隨著質譜分辨率的提高,極大地提高了修飾肽段檢測的準確度和靈敏度[44-46]。這些技術的創新使得乙酰化蛋白質組學鑒定到的乙酰化修飾的非組蛋白及其修飾位點的數量顯著增加。目前,植物乙酰化蛋白質組學鑒定到的蛋白質數量及其對應的乙酰化位點大多可以超過600個和1000個(表1)。研究者在發育中的水稻花藥[28]、水稻幼苗葉片[32]、茶樹幼苗葉片[40]中分別鑒定到了676個、866個和1286個乙酰化蛋白質,對應有1354個、1353個和2229個乙酰化位點。這些組學研究夯實了非組蛋白賴氨酸乙酰化修飾在植物中的普遍存在性,廣泛參與各個代謝途徑。

表1 植物乙酰化蛋白質組研究
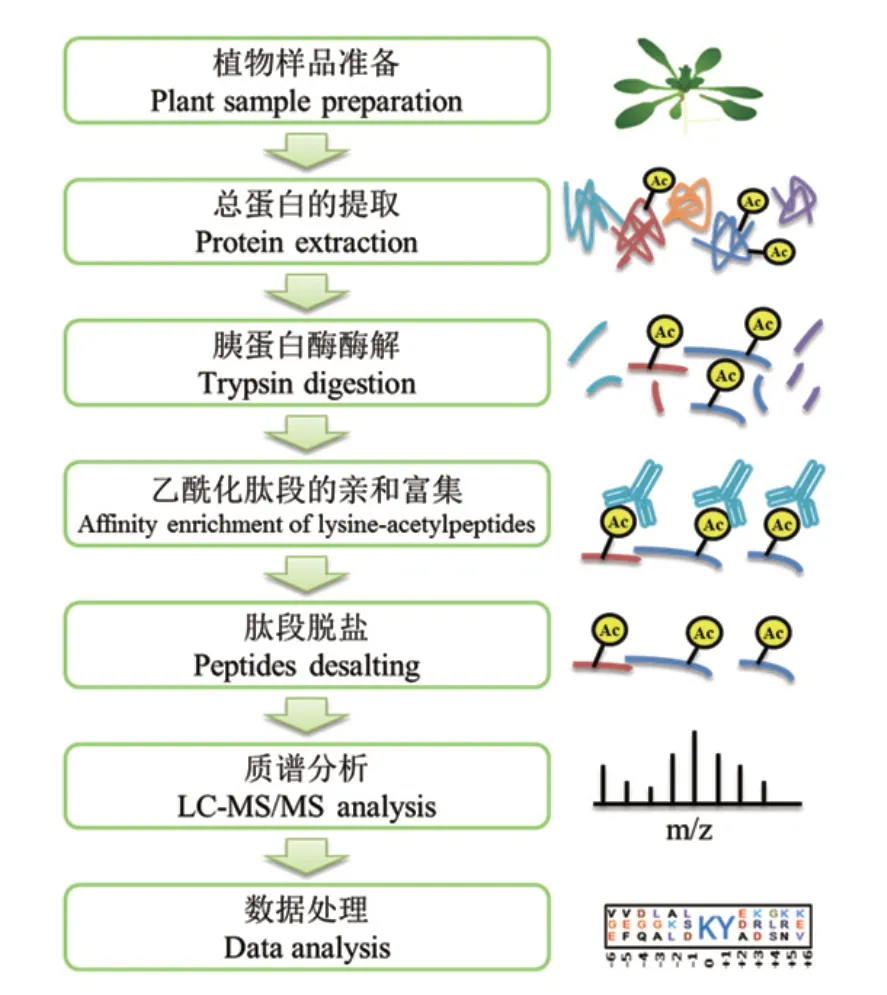
圖1 基于抗體親和富集的乙酰化蛋白質組學實驗流程
2018年,Liu等[22]對植物乙酰化蛋白質組學方法進行了改進,提出了基于氯化銫密度梯度(CsCl density gradient,CDG)離心的蛋白質分離和二甲基標記的4C定量的翻譯后修飾組學研究方法。與過去常規乙酰化蛋白質組學相比,該方法首先對提取的蛋白質樣品進行CDG離心,將蛋白質分級成上中下3層再分別進行后續分析,這有利于乙酰化修飾的膜蛋白富集。同時,不同處理的酶解肽段分別等分成2份進行輕同位素(12CH3,~28 Da)和重同位素(13CD2H,~34 Da)的二甲基標記,然后交換混合樣品后,再進行抗體親和富集和LC-MS/MS分析。最后,乙酰化蛋白質通過Mascot和SQUA-D進行定性和定量分析。該方法對擬南芥幼苗的地上部進行乙酰化修飾分析,成功鑒定到了2638個不同的乙酰化蛋白的7456個乙酰化位點,其中有4228個乙酰化位點為首次鑒定。目前為止,該研究在擬南芥中鑒定到乙酰化蛋白質數量最多,大大超過其它擬南芥或其它植物乙酰化蛋白質組分析鑒定到的600個左右的蛋白質(表1)。這些乙酰化蛋白質主要為組蛋白超家族、核糖體蛋白、熱休克蛋白以及脅迫/刺激和能量代謝相關的蛋白質。新鑒定的乙酰化蛋白質主要是一些細胞運輸蛋白、膜結合受體和受體激酶,參與油菜素類固醇、光、重力和發育信號傳導等過程。該研究也再一次擴大了非組蛋白乙酰化修飾參與的生物學過程。乙酰化蛋白質組學在蛋白質提取純化[22]、肽段富集以及質譜分析[44-46]的技術改進都提高植物非組蛋白乙酰化蛋白質定性和定量能力。今后,隨著樣品制備方法的成熟以及高通量質譜檢測靈敏度的提高[47-48],乙酰化修飾鑒定將更快速高效。未來對植物中非組蛋白賴氨酸乙酰修飾進行深度精準的的動態分析,將為后續乙酰化修飾調控機制研究提供可靠的數據支撐。
2 賴氨酸乙酰化修飾蛋白質的空間分布
非組蛋白賴氨酸乙酰化修飾廣泛存在于植物不同組織、器官和細胞器中,且空間分布具有特異性。在植物不同組織器官中,乙酰化蛋白質的種類和豐度都具有顯著差異[20,23]。通過免疫印跡法分析,擬南芥葉片、長角果、花、種子和根中的乙酰化蛋白質的豐度存在明顯的差異[20]。Uhrig等[23]通過乙酰化蛋白質組學研究也表明乙酰化蛋白質在擬南芥不同組織中的種類和豐度都存在差異,在幼苗(地上部和根系)、蓮座葉、根、花(花萼和花瓣)和長角果分別鑒定到351、199、526、146和160個乙酰化蛋白質。水稻愈傷組織、葉片、圓錐花序和根系4個典型組織的乙酰化蛋白質組分析發現,170個乙酰化蛋白質是4個組織中共有的,而97、93、27和44個乙酰化蛋白質是分別在愈傷組織、葉片、圓錐花序和根系中所特有的[26]。
植物不同的亞細胞器中乙酰化蛋白質的種類和豐度也具有顯著差異。本文對不同植物葉片中的乙酰化蛋白質的亞細胞定位的分布情況進行比較[21,31,33,35,37-38,40],結果(圖 2)顯示,90%左右的乙酰化蛋白質主要定位在葉綠體、細胞質、細胞核和線粒體。而質膜、細胞骨架、內質網和過氧化酶體等細胞器中鑒定到的乙酰化蛋白質含量極低。但是,Liu等[22]采用CDG梯度分級提取蛋白質的4C定量乙酰化蛋白質組學在擬南芥幼苗地上部鑒定到很多內質網、高爾基體和質膜上的乙酰化蛋白質,這是基于常規蛋白質提取方法的乙酰化蛋白質組學沒有發現的。因此,乙酰化蛋白質提取和富集過程中的偏好性、蛋白質降解以及脫乙酰化等都可能造成內質網和高爾基體等細胞器中鑒定到的蛋白質數量偏低[22]。
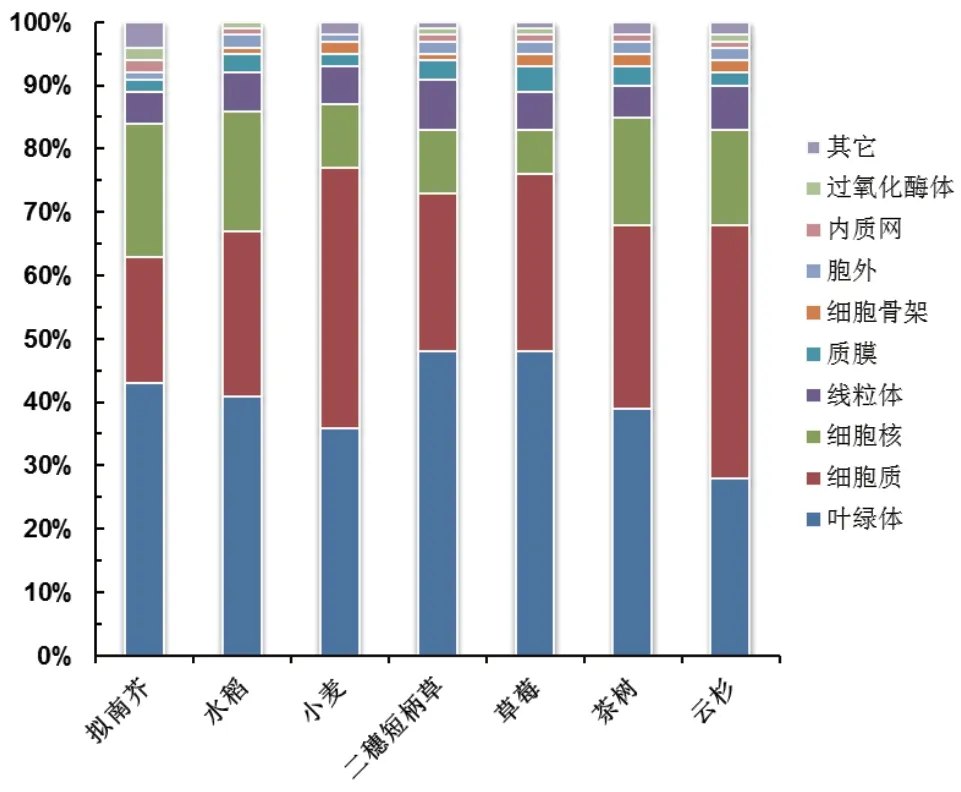
圖2 不同植物葉片乙酰化修飾蛋白質的亞細胞定位分布情況的比較
乙酰化蛋白質在植物葉片的葉綠體中比例最高,可以達到40%左右(圖 2)。葉綠體是植物光合作用發生的最主要的細胞器,這也表明乙酰化修飾可能在光合作用中發揮重要的調控作用。相一致地,乙酰化蛋白質組學在植物葉片的葉綠體中發現了大量光合作用相關的蛋白質發生了極其顯著的乙酰化修飾[19-20,33,35,37]。早期擬南芥的乙酰化蛋白質組就發現,光系統(Photosystem II,PSII)亞基和捕光色素蛋白(Light-harvesting chlorophyll a/bbinding proteins,LHC a/b)、以及卡爾文循環(Calvin cycle)中關鍵酶核酮糖-1,5-二磷酸羧化酶/加氧酶(Ribulose-1,5-bisphosphate carboxylase/oxygenase,RuBisCO)和磷酸甘油酸酯激酶(Phosphoglycerated kinase,PGK)都具有乙酰化修飾[19-20]。Zhang等[33]發現,小麥幼苗葉片捕光復合體(Light-harvesting complex,LHC)的4個 亞 基(LHCa1、LHCb3、LHCb5和LHCb6)、PS I的 兩 個 亞 基(PsaA和PsaB)、PSII的4個 亞 基(PsbO、PsbP、PsbH和Psb28)以及類囊體膜中催化PSII到PSI復合體之間的電子轉移的細胞色素復合體(Cytochrome complex,Cyt)的3個亞基(Cyt b6、Cyt f和PetD)都具有很多的乙酰化修飾位點。小麥葉片中大量卡爾文循環的代謝酶也發生了乙酰化修飾,特別是RuBisCO中含有13個乙酰化修飾位點。RuBisCO的大量乙酰化修飾在不同的植物中都普遍存在[21,31,35,37]。研究表明,擬南芥中RuBisCO的乙酰化修飾會顯著影響其三級結構形成和酶活力,進而影響光合作用的速率[19]。組蛋白脫乙酰基酶14(Histone deacetylase 14,HDA14)是葉綠體中的RPD3/HDA1類蛋白質,其大多數靶蛋白質具有光合作用[21]。HDA14功能缺失突變體的分析表明,RuBisCO的活化狀態在弱光條件下受RuBisCO活化酶(RuBisCO activase)的賴氨酸乙酰化作用控制。這些蛋白質可以通過乙酰化修飾程度來調控光合作用。研究表明,與和PSII緊密結合的LHCb相比,擬南芥中和PSII松散結合的LHCb以及游離的LHCb顯示出明顯更高的賴氨酸乙酰化水平。這表明乙酰化修飾可能在LHCb復合物在類囊體膜的分布中發揮作用[20]。乙酰化修飾是植物葉綠體中普遍存在的保守的重要翻譯后修飾,在光合作用的光反應和碳同化過程都發揮重要的調控作用。
線粒體也一直被認為是植物中乙酰化修飾參與調控的重要細胞器,因此有研究專門針對線粒體進行乙酰化蛋白質組學分析[5,39]。擬南芥[5]和豌豆[39](Pisum sativum)的線粒體乙酰化蛋白質組分析分別發現了120個和358個乙酰化蛋白質。這些蛋白質廣泛參與碳代謝、光呼吸、核酸代謝、氨基酸和蛋白質代謝以及氧化還原調節等代謝過程。特別是,碳代謝中糖酵解和TCA循環中的大量酶發生乙酰化修飾。擬南芥線粒體中TCA循環的每一步酶促反應中至少有一個酶被乙酰化修飾,包括絕大多數的丙酮酸脫氫酶復合體(Pyruvate dehydrogenase complex)等[5]。葉綠體和線粒體分別是植物光合作用和呼吸作用的細胞器,對環境變化極其敏感[49-50],研究發現參與這些代謝過程的很多蛋白質都存在翻譯后非組蛋白乙酰化修飾(圖 2),暗示乙酰化修飾在這些代謝過程和環境適應中可能發揮了重要的調控作用。
在學習角色臺詞時有很多的學習技巧,要看重語言的魅力以及潛臺詞的發掘。那么有時我悶在探究語言魅力的同時要抓住臺詞中的一些行為動詞,往往語言的魅力都會體現在動詞之中。比方說,有這樣一個經典舞臺劇,主人公在講述自己不幸遭遇的時候都會有很多臺詞,在所有的臺詞中行為動詞是最經典的,這種行為動詞是在表演是會很夸張的笑,到最后卻出現哈哈大笑。但在使用動詞時不會出現“笑著說”或“哈哈大笑著說”,不這樣安排臺詞的原因是表演的情景是主人公慘遭不幸,而所遇到的壞人卻沒有機智的頭腦,主人公才會使用這種臺詞進行抒發自己內心的情感。所以,影視表演中主人公的臺詞是由三個詞語構成,即敘述、嘲諷和贊揚。[1]
3 賴氨酸乙酰化修飾肽段的結構特征
植物不同組織中,目前鑒定到的賴氨酸乙酰化修飾蛋白質大部分只有一個乙酰化修飾位點,而一部分蛋白質則同時存在多個乙酰化修飾位點(表1)。早期的乙酰化蛋白質組[19-20,24]鑒定到的乙酰化修飾位點數量平均只有1.2個,明顯偏低。根據目前已報道的植物乙酰化蛋白質組[4,19-41],每個乙酰化修飾蛋白質的平均乙酰化位點為1.7個。最近報道的水稻種子[30]和小麥葉片[33]中都有72%的乙酰化蛋白質僅有一個乙酰化位點,而有4個或超過4個乙酰化位點的蛋白質分別只有4%(表2)。2018年,Liu等[22]鑒定的擬南芥地上部2638個蛋白質具有7456個乙酰化位點,平均每個蛋白質具有2.8個乙酰化修飾位點。只有56.6%的蛋白質僅有一個乙酰化位點,而有高達12%的蛋白質具有4個或超過4個乙酰化位點。與其它擬南芥乙酰化蛋白質組[21,23]相比,Liu等[22]從擬南芥中提取并鑒定的乙酰化蛋白質及其修飾位點數量多一倍以上,包括大量新發現的乙酰化修飾位點。該方法采用的蛋白分級提取方法不僅增加了低豐度修飾蛋白等的檢出,而且提取的蛋白質更為完整,特別是增加了膜蛋白的檢出率。因此,該乙酰化蛋白質組鑒定的乙酰化蛋白質也更為完整,得到的數據更趨向于擬南芥中真實存在的乙酰化修飾情況。這也表明了植物非組蛋白的乙酰化修飾的普遍性和重要性,在植物中仍有很多未知值得研究者發掘。
植物中乙酰化修飾位點的上下游的肽段通常具有一些保守的氨基酸序列(表2)。研究發現,草莓葉片中有近90%的乙酰化修飾位點是保守的肽段[37]。這些保守的氨基酸序列在同種植物的不同組織以及不同植物種類中也相對保守。水稻種子[30]和葉片[31]中的保守乙酰化修飾位點序列極其相似,保守序列Kac*R、Kac*K和KacH都大量存在。不同植物中,芳香族氨基酸酪氨酸(Tryosine,Y)和苯丙氨酸(Phenylalanine,F)以及帶正電荷的組氨酸(Histidine,H)和賴氨酸(Lysine,K)在保守基序中最常出現。例如,小麥葉片中乙酰化修飾的保守基序主要為KacY、KacH、KacF、LKac和FKac[33]。同時,一些植物中也有一些特有的保守基序。例如,帶正電荷的精氨酸(Arginine,R)在水稻保守基序中高頻出現[30-31],而在小麥[33]、二穗短柄草[35]、草莓[37]的葉片和云杉的體細胞胚[38]的保守基序中并沒有發現。這些保守氨基酸往往帶正電荷或具有疏水側鏈,很可能對乙酰化修飾的發生具有重要的功能。在今后的研究中,通過對乙酰化修飾保守位點的分析可以確定并優先選擇重要的關鍵氨基酸殘基進行定點突變以解析乙酰化修飾的作用方式和調控機制。
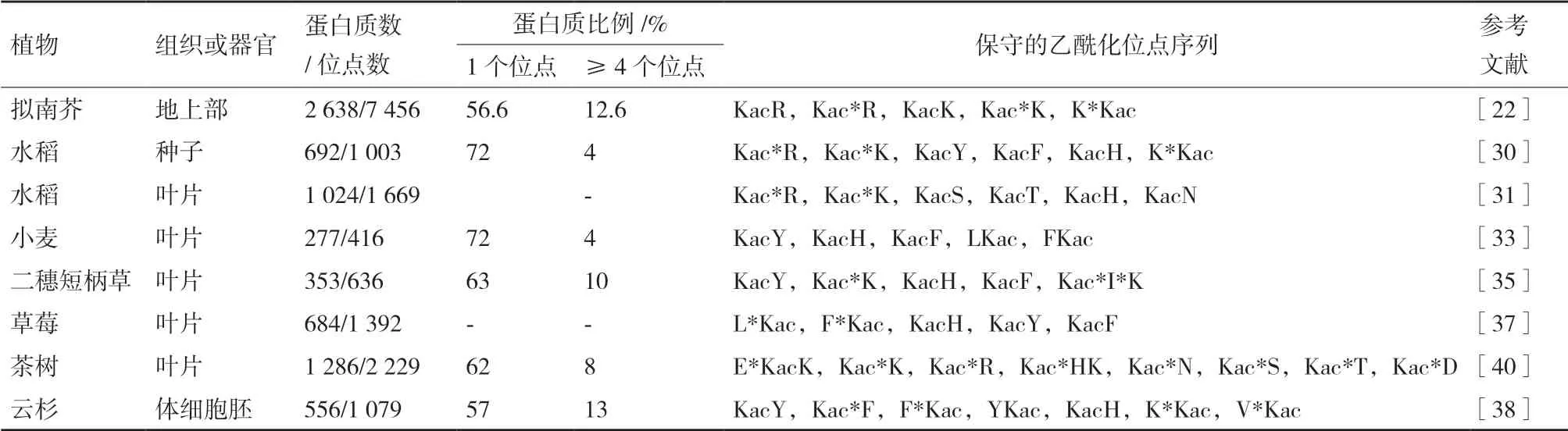
表2 不同植物乙酰化修飾位點的特征
4 非組蛋白賴氨酸乙酰化修飾在植物生長發育中的作用
植物的非組蛋白乙酰化蛋白質的分子功能大部分屬于催化活力(約45%)和結合(約40%)[30-31,33,35,37,40],這也表明植物主要通過調控酶活力和蛋白質相互作用等參與各種生物過程。其中,非組蛋白賴氨酸乙酰化修飾在植物種子萌發和生殖生長過程中的作用有著較為深入的探究。
4.1 種子萌發
非組蛋白賴氨酸乙酰化修飾在植物種子的萌發過程中的變化尤為顯著。研究發現,水稻種子在吸脹0-48 h內蛋白質的乙酰化修飾發生了顯著的變化[27]。Western blot分析顯示24 h后,蛋白質的乙酰化水平較高,乙酰化蛋白質組一共鑒定到了389個乙酰化蛋白質,包含699個乙酰化位點。這些蛋白質廣泛參與翻譯、刺激響應、葡萄糖分解代謝、糖酵解、前體代謝產物和能量、核苷酸代謝等代謝過程。這可以為后續水稻種子萌發的調控機制研究提供參考依據。
部分干燥處理可以提高針葉類植物體細胞胚的萌發能力。云杉體細胞胚的子葉和胚根的蛋白質乙酰化水平在部分干燥處理不同時間后(0、7、14和21 d)有著明顯的差異[38]。其中,處理14 d后的云杉體細胞胚的乙酰化蛋白質組分析一共鑒定到556個乙酰化蛋白質。這些乙酰化蛋白質主要參與碳代謝(糖酵解/糖異生、TCA循環和磷酸戊糖途徑)、脂肪酸途徑以及脅迫響應。蛋白質相互作用分析顯示,核糖體、蛋白酶體、剪接體和碳代謝相關的乙酰化蛋白質高度富集,它們很可能在干燥處理下的種子萌發過程中發揮主導作用。
4.2 生殖生長
非組蛋白賴氨酸乙酰化修飾在植物的生殖生長中同樣發揮著至關重要的作用。早期發育的水稻種子的乙酰化蛋白質組分析表明,非組蛋白乙酰化蛋白質占到了乙酰化蛋白質的絕大部分(948/972,97.5%)[29]。而且,在生殖生長的各個不同發育時期,花和種子都受到非組蛋白乙酰化修飾廣泛而特異性的精密調控。很多研究者對水稻花和種子在各個不同時期進行了細致的乙酰化蛋白質組研究,以剖析其中的調控機制。
植物在有性生殖過程中,減數分裂產生單倍體配子。Li等[28]通過乙酰化蛋白質組學在減數分裂時期,發育中的水稻花藥中一共鑒定到了676個乙酰化蛋白,包含1354個乙酰化位點。GO富集分析表明,染色質沉默、蛋白質折疊、脂肪酸生物合成過程以及脅迫響應相關的蛋白質大量富集。其中,超過一半的乙酰化蛋白質(357個)同時也是減數分裂早期花粉母細胞(Meiocyte)中檢測到的蛋白質[51],包括一些與水稻絨氈層和花粉發育相關的關鍵蛋白質。這些都表明乙酰化修飾廣泛參與水稻花藥發育和減數分裂。
5 非組蛋白賴氨酸乙酰化修飾在植物逆境響應中的作用
植物可以通過代謝酶的可逆乙酰化修飾來迅速感知細胞能量狀態并靈活改變反應速率或方向從而應對環境變化。近幾年,鑒于非組蛋白乙酰化修飾的廣泛調控作用,研究者通過乙酰化蛋白質組學解析植物應對逆境的響應和適應機制。
隨著水資源的匱乏和氣候變化的加劇,干旱脅迫已經成為影響農作物生產力的主要限制因子之一[52]。世界上約70%的小麥種植在干旱或半干旱地區[53],干旱脅迫已經成為限制小麥生長和產量的主要非生物脅迫因素。Zhu等[34]考察了大田實驗中干旱脅迫對發育中的小麥種子(開花后20 d)乙酰化修飾的影響。該研究一共鑒定到442個乙酰化修飾蛋白質(716個修飾位點)。其中,93個乙酰化蛋白質(106個乙酰化位點)在干旱脅迫下具有顯著變化,大部分差異蛋白與代謝(57%)和細胞(20%)過程有關。這些蛋白質參與碳代謝、淀粉生物合成、蛋白質運輸和降解、脅迫響應、轉錄等過程。干旱脅迫還導致蛋白質乙酰化和其它PTMs之間的交互作用。尤其是,淀粉生物合成的兩種關鍵酶蔗糖合酶(Sucrose synthase,SuSy)和ADP葡萄糖焦磷酸化 酶(ADP glucose pyrophosphorylase,AGPase)的乙酰化和磷酸化修飾在干旱脅迫下均發生顯著變化。干旱脅迫導致SuSy酶活力顯著增加,這表明乙酰化修飾可能在淀粉合成中發揮正調控作用。酵母分子伴侶熱休克蛋白90(Heat shock protein 90,HSP90)K294位點的乙酰化將減弱其與客戶蛋白(client protein)的相互作用,影響其與輔助伴侶蛋白的結合[17]。干旱脅迫還導致HSP90和HSP81-3去乙酰化,這可以增強其對客戶蛋白和輔助伴侶蛋白的相互作用,維持蛋白酶體的穩定性,保護細胞免受干旱脅迫的損害。
氮是植物生長發育過程中需求量最大的必需元素,缺氮嚴重抑制作物的生長發育以及最終的產量[54]。植物應對不同的氮供應有著復雜而精密的調控機制。Jiang等[40]通過乙酰化蛋白質組學解析了茶樹幼苗在氮饑餓和恢復供氮條件下葉片乙酰化修飾蛋白質的變化。恢復供氮后,光合作用、糖酵解、氨基酸代謝以及次級代謝(黃酮合成)相關的乙酰化蛋白質發生顯著變化。例如,恢復供氮后,光合系統的天線蛋白LHCa1的豐度顯著上調,而LHCa3和LHCb6的豐度顯著下調,這表明可逆的乙酰化修飾影響天線蛋白的功能,進而調控不同氮供應下葉片的光合速率。蛋白質相互分析也表明,氮供應相關的乙酰化蛋白質的多種相互作用主要涉及光合作用和核糖體。此外,還發現類黃酮合成相關的蛋白質苯丙氨酸解氨酶(Phenylalanine ammonia)、二氫黃酮還原酶(Dihydroflavonol 4-reductase)、柚皮苷3-雙加氧酶(Naringenin 3-dioxygenase)和查耳酮異構酶(Chalcone isomerase)在短期恢復供氮(3 h)和長期恢復供氮(3 d)后具有差異的修飾變化。
值得注意的是,養分脅迫下,除了碳代謝和光合作用等初級代謝過程,更多次級代謝過程的乙酰化修飾也發生特異性的響應。三角褐指藻是一種重要的產油微藻,其生長和油脂的生物合成受環境養分的影響[55]。Chen等[41]通過乙酰化蛋白質組學發現,幾乎所有參與脂肪酸合成的酶都被賴氨酸乙酰化。其中,長鏈乙酰輔酶A合成酶(Long-chain Acyl-CoA synthetase)為幾乎所有的脂肪酸代謝提供活性乙酰基。Western Blot分析顯示,該酶的乙酰化位點(K407和K425)的修飾水平受缺氮、缺磷和缺鐵脅迫影響。部分干燥處理的云杉體細胞胚處于一個缺水、缺營養和氧化脅迫的環境中。與非干燥處理相比,部分干化處理的云杉體細胞胚中的脂肪酸途徑也發生特異性富集[38]。這些次級代謝酶的快速可逆乙酰化修飾,在養分供應響應和適應的調節過程中發揮作用,有利于植物更加快速地應對不利環境。
氧化脅迫是常見的非生物脅迫方式之一。H2O2氧化脅迫下,水稻葉片中有只有12%-32%的賴氨酸乙酰化或琥珀酰化蛋白質的含量發生了變化,這表明這種調控是特異性[31]。差異蛋白質在蛋白質翻譯和折疊、光合作用和糖酵解等生物過程富集,且大部分(77%-87%)差異蛋白質的含量發生下調。嘌呤核糖體和核小體裝配相關的乙酰化位點可能特異性地參與H2O2氧化脅迫的響應。差異的乙酰化修飾蛋白在乙醛酸和二羧酸代謝、谷胱甘肽代謝以及半胱氨酸和甲硫氨酸代謝等代謝途徑富集。綜上,這些逆境脅迫下的非組蛋白乙酰化蛋白質組研究為植物應對不利環境的響應和適應的調控機制研究提供了新的參考依據。
6 賴氨酸乙酰化修飾和其他蛋白質翻譯后修飾的交互作用
蛋白質翻譯后修飾在代謝調控中具有快速、低耗和微調的特征[56-58]。除了乙酰化修飾,非組蛋白的賴氨酸還可以被其它多種類型翻譯后修飾,如琥珀酰化(Succinylation)[59]、泛素化(Ubiquitination)[60]和甲基化(Methylation)[61]等。同時,蛋白質普遍存在磷酸化、糖基化等多種PTMs,賴氨酸乙酰化修飾與其它多種翻譯后修飾之間存在交互作用(Crosstalk),形成多位點修飾,相互激活或相互抑制,協同調控各種條件下的生命過程[62-63]。
6.1 賴氨酸琥珀酰化修飾
除了賴氨酸乙酰化,琥珀酰化也是賴氨酸的一種重要的可逆酰化過程,在各種生物中廣泛存在[59]。賴氨酸琥珀酰化將琥珀酰基從琥珀輔酶A(Succinylcoenzyme A)轉化到賴氨酸的ε-氨基側鏈[64]。很多賴氨酸乙酰化的位點同時也是琥珀酰化的修飾目標,因此,研究者會同時進行賴氨酸乙酰化和琥珀酰化分析。水稻種子在吸脹24 h后,一共鑒定到389種乙酰化蛋白質(699個乙酰化位點)和261種琥珀酰化蛋白質中(665個琥珀酰化位點)[27]。其中,78個蛋白質的133個位點同時被乙酰化和琥珀酰化。這些重疊的賴氨酸修飾位點更趨向發生在極性的酸性/堿性氨基酸區域,并且暴露在蛋白質表面。這些蛋白質幾乎涵蓋所有細胞功能,其中核糖體復合物和糖酵解/糖異生相關蛋白顯著富集。二穗短柄草幼苗葉片中鑒定到了353個乙酰化蛋白質(636個乙酰化位點)和262個琥珀酰化蛋白質(605個琥珀酰化位點)[35]。其中,119個蛋白質和115個位點同時被乙酰化和琥珀酰化。這些蛋白質參與能量代謝、光合作用以及蛋白質合成等,且近一半的修飾發生在葉綠素。與乙酰化或甲基化相比,琥珀酰化增加的結構集團更大,這將導致賴氨酸結構改變更大,從而可能導致更為顯著的蛋白質結構和功能的變化。因此,賴氨酸乙酰化和琥珀酰化很可能協同調控碳代謝和光合作用等重要生物過程。
6.2 蛋白質磷酸化修飾
蛋白質可逆磷酸化是一種植物中極為普遍的蛋白質翻譯后修飾,主要發生在絲氨酸、蘇氨酸以及酪氨酸上。蛋白質磷酸化修飾幾乎參與了生命活動的所有過程,在信號傳導、細胞增殖、發育以及脅迫響應中發揮重要作用[65]。蛋白質磷酸化和賴氨酸乙酰化這兩種重要的翻譯后修飾也存在交互作用。Uhrig等[23]通過解析擬南芥不同組織(幼苗、蓮座葉、根、花和長角果)在白天結束時(the end of day,ED)和晚上結束時(the end of night,EN)乙酰化和磷酸化蛋白質豐度的變化來闡述蛋白質翻譯后修飾在調控植物的晝夜節律和光響應的作用機制。組學分析一共鑒定出909個乙酰化蛋白質和2549個磷酸化蛋白質。其中,134種參與核心植物細胞過程(捕光和光合作用、翻譯、代謝和細胞運輸)的蛋白質同時被兩種修飾,這也表明這兩種修飾在這些關鍵過程中的調控作用。Zhu等[34]在發育中的小麥種子鑒定的68個乙酰化蛋白質很可能受到磷酸化的影響。這些受磷酸化和乙酰化修飾的蛋白參與碳代謝、壓力防御和轉錄/翻譯等代謝過程。此外,計算機模擬也發現絕大多數賴氨酸乙酰化位點的定點突變都有可能影響其附近的磷酸化、甲基化和泛素化位點的修飾狀態[66]。這也表明了這些翻譯后修飾之間可能存在相互作用且相應影響,協同調控生物過程。隨著蛋白質修飾位點的種類和數量增多,PTMs之間的交互作用及其調控機制的復雜性隨之增加,檢測的難度也相應增大[67]。迄今大量研究已經證實了蛋白激酶、磷酸酶和泛素連接酶等在信號傳導過程中存在磷酸化和泛素化修飾的交互作用[62,68]。然而,有關植物中涉及非組蛋白的乙酰化修飾參與的PMTs的交互作用的系統研究仍然缺乏。今后,多種翻譯后修飾系統動態分析技術的發展將推動這一領域研究的研究。
7 問題與展望
隨著植物乙酰化蛋白質組的不斷解析,夯實了乙酰化修飾的廣泛性和重要性。賴氨酸乙酰化修飾調控不僅僅發生在細胞核的組蛋白,更多的是全面參與葉綠體和線粒體等亞細胞器以及細胞質中的多種代謝和功能的調控。非組蛋白乙酰化修飾通過多種機制影響蛋白質功能,包括調節酶活性、蛋白質穩定、亞細胞定位、調控蛋白質-蛋白質互作,蛋白質-DNA互作、與其他翻譯后修飾的共調控作用等[3]。然后,對于乙酰化修飾能否協同調節不同蛋白質參與特定生物學過程或者脅迫響應仍不清楚。今后,針對特定生物過程中乙酰化修飾的全面動態分析可能有助于了解乙酰化修飾在生長發育或應對環境變化中不同蛋白質中的協同調控過程。
毫無疑問,系統的非組蛋白賴氨酸乙酰化翻譯后修飾的研究將為調控機制和功能研究提供更多可靠的依據。目前的乙酰化蛋白質組學解析的乙酰化蛋白很可能只是細胞內乙酰化修飾的一部分。Liu等[22]建立的CDG的蛋白質分離和二甲基標記的4C定量翻譯后修飾組學解析到更多的膜蛋白翻譯后修飾,這在其它常規的植物乙酰化蛋白質組中沒有被發現。因此,該方法也為后續的植物非組蛋白乙酰化修飾的研究提供一個參考。但是,該方法步驟繁瑣,對實驗室儀器要求較高。因此,快速靈敏低成本的乙酰化蛋白質組學技術仍有待開發。高效的檢測手段將加快推進這一研究領域的進展。
蛋白質不僅可被賴氨酸乙酰化修飾,同時也受到泛素化、SUMO(Small ubiquitin modifier)、甲基化等多種蛋白質翻譯后修飾的調節[62-63,67-68]。這些翻譯后修飾過程相互影響相互作用,而且一直處于高度動態變化中。過去受檢測技術的限制,這些PTMs的交互作用沒有被系統的解析。因此,亟需提高基于質譜的檢測手段在時間和空間的分辨率來實現多種PTMs的高通量檢測。今后,多種PTMs的交互作用的解析將為我們理解植物PTMs參與植物生長發育和應對環境變化的調控機制提供全新的信息。
目前,植物應對生物或非生物逆境脅迫的賴氨酸乙酰化修飾蛋白質組學受到越來越多的關注。這些研究不僅可以揭示了植物如何快速有效應對環境脅迫,更可能為遺傳改良和育種提供了一種新的途徑。例如,植物中很多碳代謝和光合作用的代謝酶活力受到乙酰化修飾狀態的精密調控[19-21,23,32-33]。其中,Rubisco是光合作用的CO2固定的限速酶,其大亞基活性位點的乙酰化修飾狀態控制著其酶活力,通過設計非酶促乙酰化修飾的代謝物可以控制Rubisco酶活力進而提高植物的CO2固定效率[69]。因此,剖析關鍵蛋白在翻譯后修飾水平的調控機制是今后乙酰化相關研究中的重要問題之一,包括賴氨酸修飾的動態變化以及與其他PTMs的交互作用。我們希望可以通過定點突變或者CRISPR/Cas技術將賴氨酸乙酰化位點改為乙酰化或非乙酰化的類似物,進而改變代謝酶活力等,最終實現植物的生長、產量或抗逆性的提高。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
