單細胞轉錄組測序技術在心臟發育、疾病以及醫學中的應用
2021-01-22 09:09:02朱慶元李天晴
生物技術通報 2021年1期
關鍵詞:小鼠
朱慶元 李天晴
(昆明理工大學生命科學與技術學院,昆明 650093)
相對于Bulk RNA-seq測量的是測序組織所有細胞同一基因混在一起平均化后的表達水平,scRNAseq技術以其高分辨率和測序深度對每個細胞的轉錄組進行測序,對心臟研究領域產生了深遠影響。在胚胎發育過程中,心臟迅速地完成了高度精細復雜的心臟發生過程[1-2],在妊娠大約21 d就開始跳動,是第1個發揮生理功能的實質臟器[3]。目前通過單細胞測序,在小鼠模型上,已經對心臟發生在時空層面已經有了更好的理解[4],同時人鼠心臟發育具有巨大物種差異,對人類胎心的單細胞測序也在心臟發育的時間層面上以及細胞類型之間發現了人鼠特異基因的表達[5],這些對體外心肌分化研究也有非常重要的意義[6]。本文概述了scRNA-seq技術在心臟發育、疾病以及醫學中的應用及作出相應展望。
1 scRNA-seq測序技術的進展
2009年,Tang等[7]首次報道了3'偏倚的全長單細胞轉錄組測序方法。2011年,Islam等[8]建立了STRT-seq(Single-cell tagged reverse transcription sequencing),能夠在逆轉錄過程中給每個細胞加上條形碼,從而能夠同時對更多量的各種混合細胞樣本 測 序。2012年,Smart-seq(Switching mechanism at 5'end of the RNA transcript)出現,不僅能夠檢測mRNA的全長,而且有更高的轉錄本序列覆蓋度[9]。同 年,Hashimshony等[10]開 發 出CEL-seq(Cell expression by linear amplification and sequencing),進行雙端深度測序能夠準確檢測兩條鏈的序列,并且給每個細胞加上條形碼,實現了混樣單細胞的多重分析和研究。2013年,Picelli等[11]開發出 Smartseq2技術,相比Smart-seq技術,產物無需純化,產量提升。
同年,第一臺商業化單細胞轉錄組制備系統Fluidigm C1面世,能夠自動捕獲單個細胞并完成Smart-seq操作。2014年,Jaitin等[12]建立了MARSseq(Massively parallel single cell RNA-seq)技術,可分析上千個單細胞在體轉錄狀態。2015年,單細胞轉錄組測序技術中引入微流控技術,利用微流體裝置將帶有條形碼的微珠和細胞一起包裹進油滴,隨后條形碼連接到反轉錄后的cDNA上進行標記。其中Marc領導開發的inDrop只有約15萬個條形碼,處理的細胞數少,但捕獲效率高,適用于細胞數量少珍貴的樣本[13]。而Steven等開發的Drop-seq有1600萬個條形碼,可以處理大量細胞[14]。2017年,商業化的10×Genomics技術出現,其在微流控技術基礎上引入16個堿基總計400萬個細胞身份條形碼,進一步降低了技術門檻,使得更多實驗室可以進行大規模單細胞測序的研究[15]。
在技術實現高通量后,針對scRNA-seq成本高昂的問題,降低測序成本的相關技術得到發展。2017年,Gierahn等[16]開發Seq-Well技術,利用每個納米孔中捕獲1個細胞和1個帶條形碼微珠,使每個細胞的成本低于1美元。隨后2018年,Han等[17]的Microwell-seq測序平臺利用瓊脂糖材料,使得單細胞建庫測序的成本進一步降低。同年,使用SPLit-seq技術可以無需對單個細胞進行分離,而是形成單獨的RNA“隔離室”,為每個細胞的RNA引入一個獨特的組合條碼,可以特異性地標記百萬數量級的細胞[18]。
此外,空間轉錄組迅速發展,目前Slide-Seq技術可以將RNA從組織樣本轉移到覆蓋有已知位置的DNA條形碼珠子的表面,從而允許通過測序確定RNA的空間位置[19]。這種方法將轉錄組映射的空間分辨率提高到10 μm,以接近單細胞的分辨率為scRNA-seq技術提供了空間維度。隨著scRNAseq技術門檻及測序成本降低(表1),小鼠和人的心臟單細胞測序陸續地發表出來,讓更多實驗室可從心臟單細胞水平研究心臟發育、疾病以及醫學中的應用。
2 scRNA-seq測序技術探究胚胎心臟發育

表1 主要scRNA-seq測序技術
目前運用到心臟研究中的主流的scRNA- seq技術 有Smart-seq2、10×Genomics和Smart-seq/C1等(圖1)。2016年,Li和DeLaughter等分別獨立采用Smart-seq/C1技術發表了世界上前兩篇小鼠心臟單細胞測序文章,通過對不同發育階段的胚胎心臟的特定幾個解剖區域進行單細胞測序,第一次得到了主要心臟細胞類型的單細胞圖譜[23-24]。Li等[23]使用隨機森林算法分析了胚胎第8.5-10.5天的小鼠心臟的轉錄圖譜,并以超過91%的準確率成功預測心肌單細胞在發育過程中的解剖位置,并以譜系示蹤技術證實了Isl1標記的細胞主要分布在流出道和右心室處。DeLaughter等[24]則進行了胚胎第9.5天到出生后第21天的心臟細胞的測序,闡釋了胎心成熟過程中心肌細胞轉錄譜的變化。此外這兩項研究都闡釋了胚胎發育過程中心肌細胞類型的異質性并發現Nkx2.5的缺陷會導致心肌細胞成熟障礙。
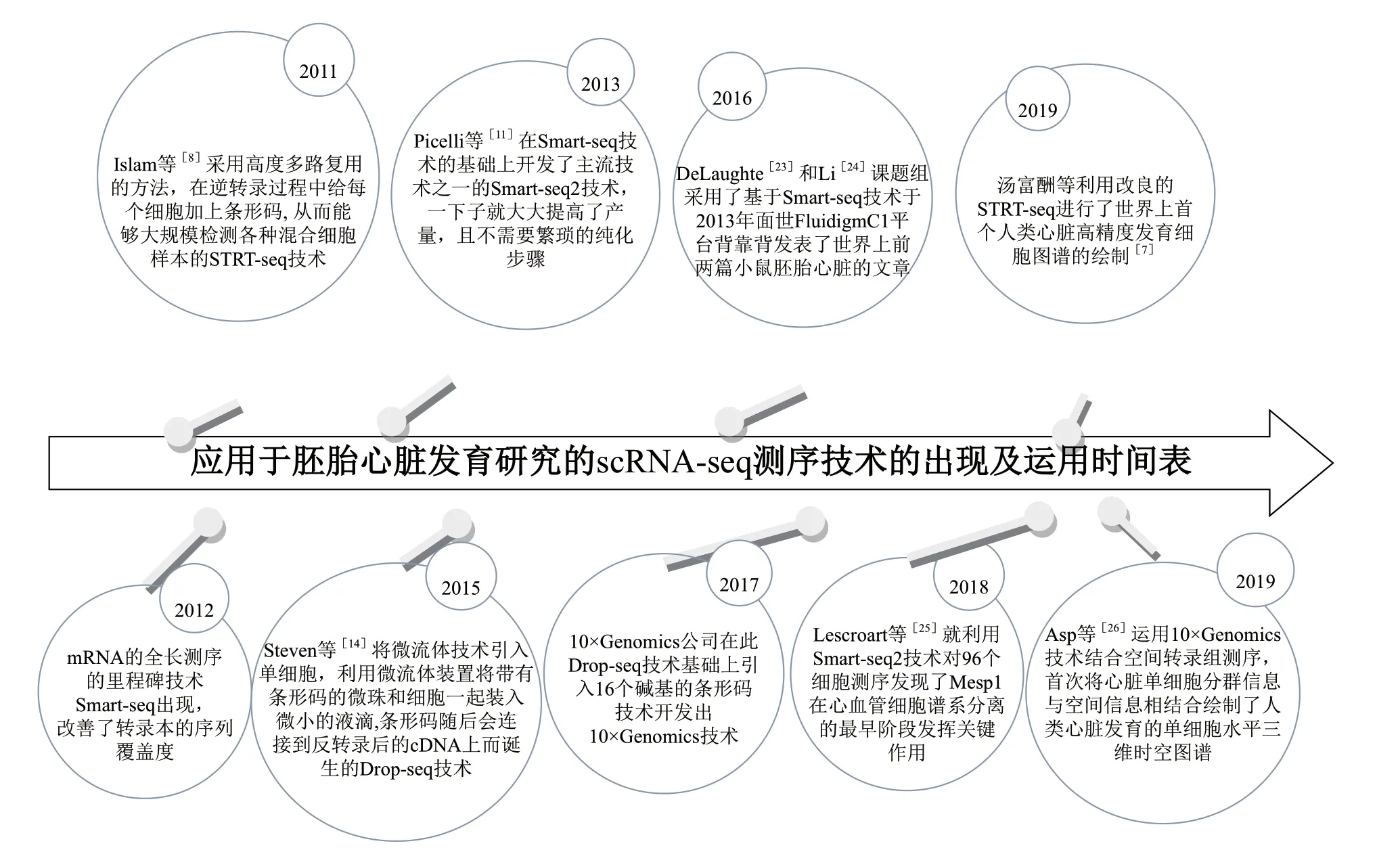
圖1 胚胎心臟研究中用到的scRNA-seq技術的開發和運用時間表
2018年,Lescroart 等[25]利用Smart-seq2技術對96個細胞測序發現了Mesp1在心血管細胞譜系分離的最早階段發揮關鍵作用,通過對比小鼠野生型的和Mesp1陰性的心血管祖細胞的單細胞轉錄譜,發現Mesp1是祖細胞退出多能性和在早期原腸形成過程中誘導心臟基因表達所必需的,并且不同的Mesp1前體細胞群對應于不同的細胞譜系和不同解剖區域的心臟祖細胞。
2019年,Cui等[5]利用改良的STRT-seq進行了首個人類心臟高精度發育細胞圖譜的繪制。該研究通過對來自18個人類胚胎(孕期從5周到25周)的大約4000個解剖學定義的心臟細胞單細胞測序,鑒定出心肌細胞、內皮細胞、成纖維細胞和瓣膜間質細胞4種主要的心臟細胞類型,并且發現心肌細胞和成纖維細胞在發育過程中經歷了基因表達的逐步變化。通過人和小鼠心臟單細胞數據的比較分析,發現了人類心臟發育的幾個獨特特征,這些差異在整體組織轉錄組測序中不明顯,但在單細胞轉錄組水平上十分明顯。4種細胞類型中,人和小鼠的心肌細胞在轉錄組上最相似。而在發育階段上,小鼠胚胎10.5 d的心肌和第7周的人的心肌發育最好的同步,而小鼠胚胎10.5 d的心臟內皮和人第6周的心臟內皮發育最同步,小鼠胚胎9.5 d的成纖維細胞和人第5周的最同步,表明小鼠和人在發育過程中心臟細胞類型的分化和成熟具有各自的同步時間線。此外,RNASE1在人內皮細胞中特異表達,THY1在人成纖維細胞中特異表達,CFB和ITLN1在人心外膜細胞中特異表達,并且人的心肌更多地表達細胞外基質基因COL1A1、COL6A3、DCN和LUM,而這些基因很少或不在在小鼠對應細胞類型上表達,相比之下,Icam2在小鼠內皮細胞中特異表達,Rnf213在小鼠心外膜細胞中特異表達。這些突顯了心臟不同細胞類型的轉錄組在物種間的相似性和差異性,便于研究者在了解人鼠不同細胞類型基因表達差異的基礎上,選擇細胞類型相近的,可以直接用小鼠研究,相差較遠的要注意小鼠模型得出結論的可靠性。另外在發育階段上,要根據研究重點選擇人鼠細胞類型發育階段匹配的時期研究。
2019年12 月Asp等[26]運 用10×Genomics技術結合空間轉錄組測序,首次將心臟單細胞分群信息與空間信息相結合繪制了懷孕前3個月發育中的人類心臟發育的單細胞水平三維時空圖譜。10×Genomics技術測得的單細胞轉錄組對應空間轉錄組技術,對整個心肌切片測序,恢復了單細胞測序完全丟失的空間信息,高精度和高準確度繪制了心臟細胞轉錄譜總體相似屬于同一細胞類型但空間分布不同的圖譜,發現神經嵴細胞和施旺前體細胞都定位在縱隔間質和流出道,并且施旺前體細胞還發現于房室外膜下間質,神經嵴細胞只在早期階段出現,施旺細胞在晚期出現,證實了神經嵴細胞對流出道分隔成主動脈和肺動脈是必須的。
3 scRNA-seq測序技術探究人心臟細胞的異質性
由scRNA-seq產生的細胞圖譜表明,心臟及其周圍血管系統由多種細胞類型組成,包括心肌細胞、成纖維細胞、內皮細胞、血管平滑肌細胞、瓣膜間質細胞和常駐免疫細胞,每種細胞類型都可以進一步劃分為亞型(表2)。
心肌目前可以分為5種亞型,其中新發現了特異表達MYOZ2和FABP3的myoz2心肌[5,26]。心房肌 高 表 達NR2F1、FOS、HEY1、EGR2、CREB3L2和HAND2;而HAND1、HEY2、IRX3和NFIA則特異性表達在心室肌,此外跟心房肌相比,心室肌更高的表達細胞外基質基因DCN和FBN2。從轉錄因子來看,轉錄因子IRX3和HAND1在左心室高表達對左心室發育至關重要,房性心律失常相關基因PITX2則在左心房高表達。此外心肌又可以分為致密化心肌和小梁肌,其中小梁肌表達NPPA和GJA5等標志基因[23]。致密化心肌和小梁肌又都可以進一步通過標志基因區分是位于心房還是心室。致密化心室肌特異表達MYH7,S100A4和LBH,而致密化心房肌則特異表達MYH6,MYL7和ULK4;ITGA6和RELN特異富集在心房的小梁肌,CRABP2和SLIT2則特異富集在心室的小梁肌。通過單細胞測序獲得各種心肌細胞亞型的標記基因,在此基礎上對特定細胞亞型做進一步研究。
內皮細胞可以分為4種亞型,其中對于心內膜細胞,Cui等[5]和Asp等[26]可能因為缺乏相關心臟方面的背景知識,把它當成了毛細血管內皮。心內膜細胞是一種內皮細胞,表達CDH5、PECAM1等內皮標記的同時,特異表達NPR3,可以作為心內膜區別冠狀動脈內皮等其他內皮的特異標記基因。瓣膜內皮則更高的表達NTRK2和NFATC1。冠狀動脈內皮特異表達FABP4和CD36,與血管發育和促進心肌脂肪酸的使用相關,且它們對脂質的作用與成體后發生冠狀動脈粥樣硬化有很大關系。
單細胞測序數據結合空間轉錄組顯示[5,26],胚胎第5周心外膜還未出現時,心外膜前體細胞既表達UPK3B、ALDH1A2、WT1和TBX18等心外膜標記,但其基因表達模式又和心外膜不同,其增殖細胞率是非免疫細胞中最高的,特異表達NOTCH信號通路的兩個靶基因HEY1和HEY2,可能其命運被NOTCH信號通路所控制。而心外膜細胞更加成熟,表達補體成分基因CFB、C3、C1R和C1S以及細胞外基質基因ELN和DPT。空間轉錄組顯示,心外膜前體細胞表達TCF21主要位于房室心外膜下間質。
結合空間轉錄組區分位置,單細胞轉錄組提示亞型功能,心臟中分出5種成纖維細胞[26]。第1種成纖維樣細胞主要集中在流出道根部和瓣膜,第2種成纖維樣細胞主要在流出道和形態發生相關,第3種成纖維樣細胞和心外膜前體細胞一樣都定位在房室心外膜下間充質,可能和冠脈形成有關。心外膜下第4種成纖維樣細胞和結締組織發育以及血管發生有關。第5種成纖維樣細胞定位更朝向流出道,與動脈和主動脈形態發生相關,調節內皮細胞增殖。
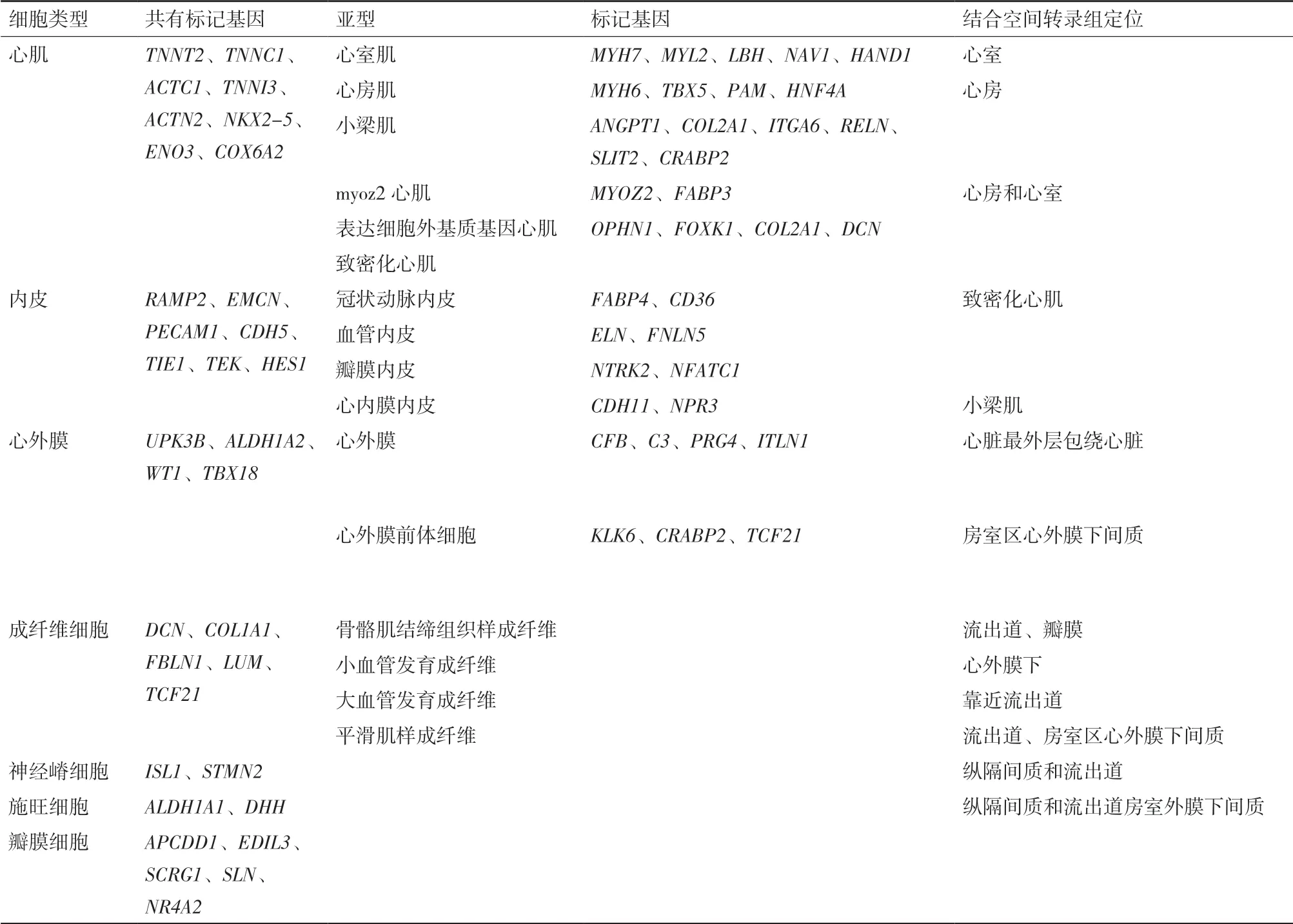
表2 心臟細胞主要類型及其基因表達特點和空間定位
4 scRNA-seq測序技術在心臟健康大動脈和動脈粥樣硬化動脈方面的研究
將scRNA-seq與譜系示蹤轉基因小鼠相結合,揭示了冠狀動脈是由發育過程中靜脈內皮細胞轉變的特定動脈內皮前體細胞群所分化而來[27]。這類單細胞群的轉錄譜由靜脈表型逐漸轉變為動脈表型,其中靜脈的特異性轉錄因子Couptf2為這種命運轉換的核心轉錄因子,為進一步研究靜脈衍生動脈的機制奠定了分子基礎。人的單細胞轉錄組顯示人的冠狀動脈內皮特異表達FABP4和CD36,與血管發育和促進心肌脂肪酸的使用相關,且它們對脂質的作用與成體后發生冠狀動脈粥樣硬化有很大關系[26],對冠狀動脈這兩個特異基因的研究有可能找到預測發生冠狀動脈粥樣硬化的風險指標,以及研發藥物預防冠狀動脈粥樣硬化及繼發的心肌梗塞。通過健康成年小鼠的主動脈單細胞圖譜確定了主動脈的所有細胞類型[28],進一步比較了8周和18個月小鼠的主動脈,發現年輕和老年小鼠主動脈內皮細胞種群在轉錄水平上存在顯著差異[29]。
scRNA-seq還被用來研究主要血管疾病的細胞狀態和命運決定。動脈粥樣硬化后由多種細胞類型組成,包括內皮細胞、血管平滑肌細胞和免疫細胞,其中血管平滑肌細胞是具有高度可塑性的終末分化的細胞類型,單細胞分析的將其定義為一群特異的成纖維細胞樣細胞,并進一步鑒定出引導血管平滑肌細胞的分化狀態的特定的組蛋白變體[30-31]。動脈粥樣硬化的典型特征還包括大量的免疫細胞,但由于細胞標記物的缺乏,巨噬細胞亞群的功能和表型都沒有很好的定義,為此,將健康和動脈粥樣硬化主動脈中巨噬細胞的scRNA-seq數據進行對比,鑒定出病變主動脈中富集的髓系亞群[32]。這些亞群包括單核細胞、單核細胞來源的樹突狀細胞和兩個巨噬細胞亞群。這些巨噬細胞群僅見于動脈粥樣硬化后的主動脈,Trem2和Il1β等炎癥分子高表達。類似地,將單細胞測序與質譜分析相結合,在小鼠主動脈粥樣硬化斑塊中鑒定出11種不同的白細胞類型,這些主動脈白細胞的組成可以用來預測動脈粥樣硬化患者的臨床事件,體現出單細胞測序臨床轉換潛力[33]。
5 scRNA-seq測序技術與多能干細胞分化心血管的研究
人多能干細胞誘導分化的細胞模型已經被證明在特定患者的疾病建模和再生治療中是有用的,但多能干細胞來源的心血管細胞還是存在成熟度不夠和異質性較大的問題。因此,許多研究已經使用scRNA-seq來確定參與分化轉錄調控的信號通路,識別由分化引起的所有細胞類型,并優化和修改方案以產生成熟和均質性較高的心臟血管細胞亞型。Friedman等[34]在誘導多能干細胞干細胞分化心肌(Induced pluripotent stem cells derived cardiomyocytes,iPSC-CMs)分化的不同階段對超過40000個細胞進行了scRNA-seq測序,發現HOPX的調節失調導致了iPSC-CMs持續的未成熟狀態。在另一項單獨的研究中,取了iPSC-CMs分化過程中大約10000個細胞鑒定心臟轉錄因子發現表達NR2F2和TBX5的iPSC-CMs具有更不成熟和心房樣的特征,而HEY2、IRX4和MYL2富集表達在更成熟的心室肌。通過基因編輯NR2F2和HEY2,證實了這兩個轉錄因子分別產生心房樣和心室樣iPSC-CMS的調控作用[35]。總之,這些研究結果突出了iPSC-CMS的異質性,并揭示了控制細胞成熟和腔室特化的特定心臟轉錄因子。人誘導多能干細胞分化來源的內皮細 胞(iPSC-derived endothelial cells,iPSC-ECs)與iPSC-CMS一樣受到細胞不成熟和異質性的限制。在iPSC-ECs的分化過程中鑒定出4個主要的iPSC-ECs亞群,CLDN5+簇代表代謝活性的iPSC-ECs,GJA5+簇代表動脈樣的iPSC-ECs,APLNR+簇代表炎癥反應性iPSC-ECs,ESM1+亞群代表活化的細胞[36]。McCracken等[37]對兩個分化方案來源的人胚胎干細胞的內皮細胞(esc-ec)進行了scRNA-seq分析。擬時間分析顯示分化第6天開始分化為內皮細胞和間充質細胞類型,這兩種細胞類型在分化第8天開始成熟,以間充質細胞群中SNAI2表達減少,內皮細胞中ANGPT2、ESM1和GNG11表達增加為顯著特征。盡管ESC-ECs在最初的內皮命運決定后進一步成熟,但沒有觀察到器官特異性內皮標記基因表達。未來的研究應該集中在進一步優化分化方案以產生組織特異的成熟ECs。
6 scRNA-seq測序技術與心肌梗死與再生
Gladka等[38]對小鼠心梗缺血-再灌注模型進行心臟單細胞測序,發現在各種細胞類型中出現了以前未知的細胞亞群,尤其是鑒定出一群Ckap4表達增加的成纖維細胞群。冷凍損傷斑馬魚心臟交界區心肌細胞的scRNA-seq顯示,損傷后交界區心肌細胞的單細胞轉錄組與胚胎心肌細胞相似,這解釋了在斑馬魚心臟再生過程中觀察到的小梁心肌細胞向皮質心肌細胞的轉換,從而產生了小梁細胞和皮質細胞[39]。為了研究心肌梗死后新生血管內皮細胞的異質性,從內皮特異性譜系追蹤的梗死7 d后小鼠心臟中分離的內皮細胞進行scRNA-seq,鑒定出的10個內皮細胞簇中,其中有5個簇在心肌梗死后組顯著富集,這些簇中與心臟重塑、內皮細胞-細胞外基質相互作用、增殖和細胞周期調節相關的基因表達上調。其中Plvap在心肌梗死小鼠模型和人類心臟模型的這5個簇中的3個中高表達,且僅局限于梗死邊緣區的內皮細胞[39],與梗死后促進內源性心臟組織修復的生長因子的定位相同[40]。類似的,心肌梗死后7 d的每個非心肌細胞群體都有自己的中間過渡狀態和獨特的轉錄信號,越來越多的研究已經使用scRNA-seq來闡明心肌梗死后各種非心肌細胞群體的作用,促進了人們對缺血組織重建過程中微環境變化的理解。未來的研究將受益于對這些細胞類型和過渡狀態的特定功能的更深入的機制研究,特別是在損傷后的不同階段,如早期急性炎癥(梗死后1 d)和晚期重塑(梗死后30 d)。這樣的研究將允許更好地描述病理性心臟纖維化期間內皮細胞到間充質細胞轉變或間質細胞到內皮細胞轉變過程中的動力學特征。目前,非心肌細胞和心肌細胞之間在組織修復和重塑過程中的細胞間通訊的轉錄研究和隨后的驗證還沒有得到充分的研究,填補這些知識的空白對于提高人們對心臟損傷和修復不同階段所涉及的細胞反應的理解至關重要。
7 scRNA-seq測序技術與先天性心臟畸形以及心衰
Nkx2.5是心臟早期發生重要的轉錄因子[41],其顯性突變會導致房間隔缺損[42],DeLaughter等[24]利用Nkx2.5+/-單倍型不足的小鼠心肌單細胞測序發現心肌和內皮的成熟都推遲了,推測心肌對心內膜的分化至關重要。而Li等[23]發現Nkx2.5-/-胚鼠心肌直接丟失了心室的轉錄譜,轉錄異常,沒有心室的表型,只能表現出左房心肌的表型。致密化心室肌特異表達MYH7,而MYH7突變在心肌致密化不全中最常見[43],可能為心肌致密化不全好發于左心室游離壁提供了一定的解釋[5]。此外,發現人第7周的心內膜高表達配體NRG1,他們的受體ERBB2和ERBB4高表達在小梁肌,但很少有細胞表達BMP10,暗示這一時期心內膜通過NOTCH信號通路調節心肌致密化主要是是通過ERBBs信號來促進分化,而不是通過BMP10誘導的增殖[44],這與先前從小鼠研究中發現基因Bmp10的突變與胚胎發生過程中心肌致密化不全的發展密不可分提出了不同見解[45],當然這也不排除是物種間的差異。GATA6突變導致永存動脈干、法洛四聯癥等動脈畸形[46-47],而GATA6調控晚期心肌發育[5],這之間也尚待研究。此外房性心律失常相關基因PITX2則在左心房高表達[5],是否暗示房性心律失常異位起搏點多發生左心房,還需進一步研究。有研究報道MYH6是病竇綜合征[48]的易感基因,而MYH6在致密化心房肌中特異表達,研究致密化心房肌對臨近的竇房結的信號通路可能是找出其中原因的一個方向。Gollob等[49]發現GJA5是房顫的重要致病基因,而GJA5特異表達在小梁肌。此外,細胞外基質重構異常與心衰相關,目前單細胞測序發現細胞外基質不僅主要來源成纖維細胞,心肌和其他類型心臟細胞也會分泌細胞外基質,且心肌隨著細胞外基質基因表達增加而不斷成熟,對細胞外基質的研究可能是闡明心衰相關疾病機制的一個方向。Nomura等[50]在單細胞水平鑒定出肥厚型心肌病心肌特異轉錄譜。其中值得注意的是,基于擬時分析,在DNA氧化損傷積累后,P53依賴的Mef2Nrf2信號軸在促進肥厚心肌細胞致病基因激活方面發揮了關鍵作用,這些發現進一步被來自心力衰竭患者的心肌細胞scRNAseq獲得的數據所證實[50]。
8 結語與展望
本文概述了scRNA-seq技術在胚胎心臟發育、心臟細胞的異質性以及在心血管方面、多能干細胞分化心血管和心臟疾病方面的進展。
在胚胎心臟細胞的異質性方面,至今也還沒有分出淋巴內皮和起搏整個心臟的竇房結細胞群。在中樞神經系統有報道說淋巴內皮具有引導血管內皮再生功能,這對心梗之后心臟再生血管重建可能也有重要的意義[51]。而分化出人的竇房結起搏細胞代替心臟起搏器有很大的臨床轉化價值,但竇房結區域小,缺乏明確形態標記,異質性高,有大比例的心房細胞和非起搏細胞,從而需要設計出更巧妙的實驗捕獲到足量純化的竇房結細胞進行單細胞測序。
在心臟再生方面,人類成體心肌再生能力有限,而在心肌有再生能力的動物模型上,梗死區強烈的炎癥反應是原位的單細胞測序到的免疫細胞[5],還是通過淋巴管,血管趨化而來還需進一步研究。此后心肌細胞增殖顯著上升,成纖維細胞參與了心肌梗死后的纖維疤痕過程[52],可以借助scRNA-seq技術鑒別哪種亞型的成纖維細胞參與了這一過程。此外,前期研究認為TBX18陽性的心外膜可以作為心臟前體細胞產生心肌細胞[53],單細胞測序結果也顯示其作為心臟前體細胞有心臟發育的潛能[5,26]。目前,成體干細胞,誘導多能干細胞,細胞重編程,心臟組織工程[54]等心肌再生的方法都已經在實驗中,但仍沒有突破性成果,希望單細胞測序分析進一步了解心肌再生機制,找到最適于再生的心肌細胞亞型及其核心轉錄因子,分化或者原位將成纖維細胞重編程成成熟有功能的心肌細胞。
除了心臟再生,先天性心臟病也亟待單細胞測序進一步研究,如已知神經嵴細胞與很多先天性心臟病相關,需要進一步通過Smart-seq2等scRNA-seq技術進一步測序研究。此外,還可以對先天性心臟病流產胎兒心臟組織單細胞測序,然后與現有正常流產胎兒數據作比較,發現可能致病機制。
最后考慮到不同患者在心血管疾病藥物治療反應上的顯著差異,通過普通轉錄組也只能發現iPSCCMs對常見心血管藥物轉錄組譜整體上存在人際差異,卻無法鑒別出靶細胞群之間藥物敏感性差異。在治療前基于scRNA-seq技術得到的細胞異質性的評估,可以為醫生確定最有效的、個性化的治療方案提供依據。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
