苦蕎NAC轉錄因子FtNAC17的鑒定及表達分析
2021-01-22 09:09:04榮玉萍唐彬李鵬章潔瓊陳慶富朱麗偉鄧嬌黃娟
生物技術通報 2021年1期
榮玉萍 唐彬 李鵬 章潔瓊 陳慶富 朱麗偉 鄧嬌 黃娟
(1. 貴州師范大學蕎麥產業技術研究中心,貴陽 550001;2. 貴州省農作物技術推廣總站,貴陽 550001)
植物在生長發育過程中受環境因素的影響較大[1]。這些環境因素包括生物脅迫(如病蟲害)和非生物脅迫(如干旱、低溫、鹽脅迫、水澇、機械損傷)。轉錄因子在植物應對逆境脅迫過程中具有重要作用[2]。其中,NAC(NAM/ATAF/CUC)是植物特異的轉錄調控因子,是植物中數量最多的轉錄因子家族之一[3],其N端含有約150個氨基酸的保守結構域,由A、B、C、D、E 5個亞結構域組成,其中A、C、D高度保守,B、E較為多變;NAC轉錄因子C端為高度變異的轉錄調控區,在植物生長發育及應對生物及非生物脅迫具有重要作用[4]。根據NAC轉錄因子中保守的NAM結構,可將植物中NAC轉錄因 子 分 為ONAC4、ANAC034、SNAC、TIP、SND、NAM/CUC3六大類,同一大類的NAC基因具有相近的功能[5],其中,響應非生物脅迫的NAC基因都歸屬于SNAC大類。干旱、低溫和鹽脅迫是較為普遍的非生物脅迫。干旱對于植物生長的影響顯著,尤其葉片面積和葉片數量都會顯著減少[6]。過表達OsNAC52增強了轉基因擬南芥對干旱脅迫的耐受性[7]。低溫可引起細胞膜凍結,并通過滲透、氧化和其他脅迫抑制植物的生長,造成不可逆轉的損害,嚴重影響作物產量和質量[8-9]。鹽脅迫除了引起植物失水,還會引起植物內蛋白質和脂類破壞和水解,導致代謝紊亂,甚至引起植物體死亡。此外,鹽脅迫下活性氧的過度積累加速了植物萎蔫[10]。過表達LpNAC13的轉基因煙草具有較強的耐鹽性[11]。蕎麥屬于蓼科(Polygonaceae)蕎麥屬(Fagopyrum),是一年生草本雙子葉植物[12],其栽培種有甜蕎(F.esculentum)和苦蕎(F.tataricum)兩種。苦蕎作為一種藥食同源的作物,不僅富含蛋白質、淀粉等營養物質,還含有膳食纖維、維生素等微量物質以及黃酮類化合物、多酚類化合物等活性成分,具有降三高、抗腫瘤、增強免疫力等功效[13]。除此之外,苦蕎還具有耐瘠薄的特性[14]。因此,通過分析苦蕎NAC基因在干旱、低溫、鹽和激素脅迫下的響應情況,進而明確其響應非生物脅迫機制。這對于挖掘優異基因具有重要意義。
目前,對苦蕎的研究主要集中在營養保健和食品加工方面,在分子生物學方面的研究尚處于起步階段。在分子方面的研究主要集中在黃酮類化合物的合成及代謝途徑中關鍵酶和轉錄因子的研究[15]。隨著苦蕎基因組數據庫的構建[16],NAC家族轉錄因子在苦蕎全基因組水平上得到分析和鑒定[17-18]。且對苦蕎FtNAC15進行了系統性分析,結果表明,FtNAC15參與種子發育及響應干旱脅迫[19]。Deng等[20]通過對苦蕎的8個與脅迫相關NAC基因進行研究發現,所有的FtNAC基因對一個或多個處理都有反應,其中FtNAC4和FtNAC7在鹽、干旱、脫落酸和水楊酸處理后上調超過20倍。
雖然有部分的NAC轉錄因子在苦蕎中的研究有了初步的結果,但大部分NAC轉錄因子具體功能尚不清晰。本研究根據已有的轉錄組數據挑選并克隆了一個NAC家族基因,命名為FtNAC17,對其編碼區序列、氨基酸序列等進行生物信息學分析以及其對干旱、低溫、鹽和激素脅迫的響應進行分析,為闡明基因功能提供理論依據。
1 材料與方法
1.1 材料
采用標準發芽法將苦蕎(貴米苦11號)種子培養14 d,待長出一片真葉后,進行6種處理。選飽滿一致的種子36粒,按照6行6列的方式整齊地擺放到鋪有濾紙的發芽盒(12 cm×12 cm×6 cm)內,25℃光照培養箱中培養(光照16 h/黑暗8 h)至長出一片真葉后進行統一處理。
干旱處理:采用20%聚乙二醇(PEG-6000)溶液處理[21]。低溫處理:將培養后的幼苗在4℃處理[22]。鹽處理:用3 mg/L NaCl處理幼苗[23]。激素處理:激素處理又分為3種,分別為22.43 mg/L茉莉酸甲酯處理[24]、20 mg/L脫落酸處理[25]和150 mg/L赤霉素處理[26]。分別在處理后0、3、6、12和24 h取整株幼苗,液氮速凍后立即-80℃保存,以備提取RNA。每種處理設置2組生物學重復。
另取苦蕎幼苗用于DNA的提取,采樣地點為貴州師范大學蕎麥產業技術中心百宜基地。將采集的樣品置于液氮中,-80℃保存備用。
1.2 方法
1.2.1 DNA的提取與檢測 苦蕎總DNA的提取用改良的CTAB法。將大約0.1 g苦蕎幼苗于研缽中加入液氮充分研磨后轉入離心管中,加入1000 μL CTAB提取液混合均勻,65℃水浴30 min。離心取上清,加入800 μL氯仿/異戊醇抽提。離心取上清,加入500 μL預冷的異丙醇,混合均勻。離心去上清,加入500 μL預冷的無水乙醇清洗沉淀2次。離心后晾干乙醇溶液,溶于100 μL ddH2O中。將提取的DNA經1%的瓊脂糖凝膠電泳檢測,質量合格后用于下一步的克隆。
1.2.2 RNA提取與cDNA合成 利用天根生化科技(北京)有限公司的植物總RNA提取試劑盒提取處理樣品中總RNA。取1 μg RNA用寶日醫生物技術(北京)有限公司的反轉錄試劑盒合成cDNA第一鏈,-20℃保存備用。
1.2.3 苦蕎FtNAC17的克隆 根據苦蕎基因組數據庫中苦蕎基因組序列及預測的編碼區序列,利用Oligo7引物設計軟件設計1對特異性引物擴增苦蕎NAC17序列(FtNAC17F:5'-AAATGTCGTCGGAAGACTTGCA-3',FtNAC17R:5'-CTAGCTCAGATACATGAACATATCC-3'),于生工生物工程(上海)股份有限公司合成引物。以苦蕎DNA和cDNA為模板,FtNAC17F和FtNAC17R為引物,利用寶日醫生物技術(北京)有限公司的熱啟動型TaKaRa Ex Taq酶進行PCR擴增。PCR反應體系為DNA或cDNA 2 μL、10×PCR Buffer 5 μL、dNTP Mixture 4 μL、上下游引物各2 μL、Taq DNA聚合酶0.25 μL和ddH2O 34.75 μL。PCR反應程序為98℃ 3 min;98℃ 10 s,42℃ 30 s,72℃ 1 min,30個循環。將擴增產物進行1%瓊脂糖凝膠電泳,在凝膠成像儀上檢測,片段大小正確后利用天根生化科技(北京)有限公司的DNA膠回收試劑盒進行膠回收。回收產物連接于天根生化科技(北京)有限公司的pGM-T載體上,轉化到大腸桿菌TOP 10感受態細胞中,挑取陽性單克隆,經菌液PCR鑒定后送至生工生物工程(上海)股份有限公司進行測序。
1.2.4 苦蕎FtNAC17的生物信息學分析 將經測序驗證正確的序列進行生物信息學分析。利用GSDS(http://gsds.cbi.pku.edu.cn/)對FtNAC17的基因結構進行分析。用DNAMAN軟件分析其開放閱讀框,預測其編碼的氨基酸序列;利用SMART(http://smart.embl-heidelberg.de/)分析蛋白質保守結構域。利用EXPASY(https://web.expasy.org/)預測蛋白質的理化性質。利用PSORT Ⅱ Prediction(https://psort.hgc.jp/cgi-bin/runpsort.pl)軟件預測蛋白亞細胞定位。使用GOR IV SECONDARY STRUCTURE PREDICTION METHOD(https://npsa-prabi.ibcp.fr/cgibin/npsa_automat.pl?page=npsa_gor4.html)預測蛋白的二級結構。利用BLASTP將蛋白序列與NCBI非冗余蛋白庫(NR)進行比對,然后利用BankIt(https://www.ncbi.nlm.nih.gov/WebSub/)將FtNAC17序列提交到GenBank。利用DNAMAN將與FtNAC17同源的NAC蛋白進行氨基酸多重序列比對。利用MEGA 6.0軟件構建進化樹,使用鄰接法,設置重復次數(bootstrap)為1000次。
1.2.5 實時熒光定量PCR 利用Primer 6設計熒光定量PCR引物qNAC17-F:5'-GAATCGGTGCCACGGTGTCA-3'和qNAC17-R:5'-ACCTTGTGCAGACGGCTGATTA-3'。將1.2.2中合成的cDNA稀釋10倍為模板,以Actin作為內參基因,以qNAC17-F和qNAC17-R作為引物進行PCR擴增。利用寶日醫生物技術(北京)有限公司的TB Green Premix Ex Taq II進行熒光定量PCR[27]。反應體系為cDNA 2 μL、TB Green Premix Ex Taq II 10 μL、上下游引物各0.8 μL和ddH2O 6.4 μL。反應程序為95℃ 30 s;95℃ 5 s,55℃ 34 s,40個循環。2次生物學重復,2次技術重復。采用2-△△Ct法進行相對表達量的處理。
2 結果
2.1 苦蕎FtNAC17的克隆
以苦蕎DNA和cDNA為模板,利用特異性引物進行PCR擴增,得到大小約為1000 bp的序列(圖1)。經測序,該片段大小為903 bp。利用GSDS分析基因結構,結果顯示,在該基因中存在2個外顯子(1-438 bp和523-903 bp),其中包含一個內含子(439-522 bp)(圖2)。將該基因命名為FtNAC17,GenBank登錄號為MT641452。
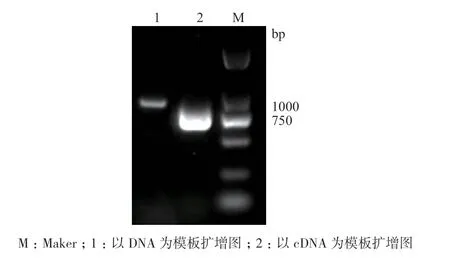
圖1 苦蕎FtNAC17的擴增
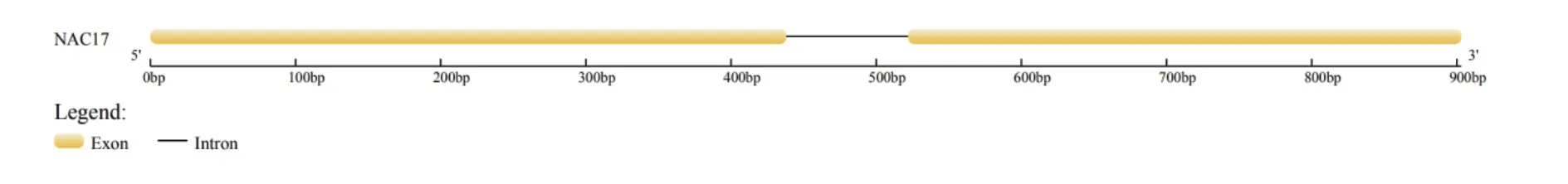
圖2 FtNAC17的結構
2.2 苦蕎FtNAC17蛋白生物信息學分析
利用DNAMAN對該序列進行分析發現,其含有819 bp的開放閱讀框,編碼272個氨基酸(圖3)。利用SMART分析蛋白質結構域,顯示在其9-132位氨基酸序列是NAC蛋白特有的NAM結構域,其E-value值為1.5e-37,說明其屬于NAC轉錄因子。利用EXPASY預測蛋白質的理化性質,結果表明,FtNAC17蛋白的分子質量為30.77 kD,等電點為7.05,疏水性平均值為-0.584,推測該蛋白可能為親水性蛋白(表1)。利用PSORT Ⅱ Prediction對FtNAC17蛋白進行亞細胞定位預測,結果表明FtNAC17蛋白有45%的可能性在細胞質中(表2)。綜上所述,FtNAC17蛋白很可能存在于細胞質中。

圖3 FtNAC17的核酸及其編碼的氨基酸序列

表1 FtNAC17蛋白的理化性質

表2 FtNAC17蛋白亞細胞定位
利用GOR IV SECONDARY STRUCTURE PRE-DI CTION METHOD對FtNAC17蛋白進行二級結構預測,結果表明FtNAC17蛋白的二級結構由α-螺旋、β-折疊和無規則卷曲構成。二級結構中α-螺旋有32個氨基酸,占序列的11.76%;β-折疊有70個氨基酸,占序列的25.74%;無規則卷曲有170個氨基酸,占序列的62.5%(圖4)。上述數據表明FtNAC17蛋白的二級結構以無規則卷曲為主,β-折疊其次,α-螺旋數量最少。
2.3 FtNAC17蛋白同源進化分析
將FtNAC17序列與擬南芥TAIR11蛋白庫比對發現,該基因與擬南芥中SNAC類中ANAC002最為相近。為推測FtNAC17的功能,從植物NAC轉錄因子家族6大類中挑選具代表性的蛋白,分別為ONAC4、ANAC034、ANAC002(SNAC)、ANAC078(TIP)、ANAC012(SND)、NAM/CUC3(ANAC018、ANAC031)。將FtNAC17與這7條蛋白NAM保守結構域序列進行多重序列比對,結果表明,NAC蛋白N端的氨基酸序列較為保守,由A、B、C、D、E 5個亞結構域組成(圖5)。將以上氨基酸序列構建系統進化樹,結果表明,FtNAC17與擬南芥ANAC002(ATAF1)親緣關系最近(圖6)。
2.4 FtNAC17的表達分析
為揭示FtNAC17響應干旱、低溫、鹽和激素脅迫情況,采用熒光定量PCR方法來探究FtNAC17在干旱、低溫、鹽、茉莉酸甲酯、脫落酸和赤霉素處理苦蕎幼苗0、3、6、12和24 h的表達模式(圖7)。結果表明,6種非生物脅迫均有顯著影響(P<0.05)。尤其是赤霉素處理下,FtNAC17的表達水平最高(達10.27倍)。

圖4 FtNAC17蛋白二級結構預測

圖5 NAM保守結構域氨基酸多重序列比對
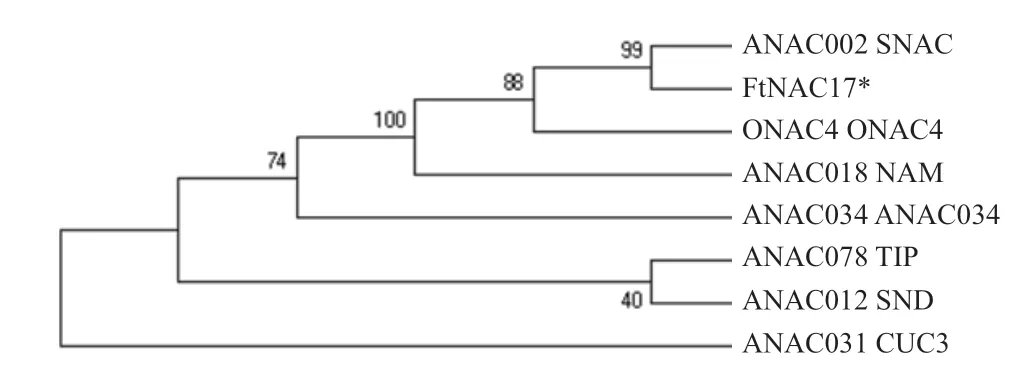
圖6 FtNAC17與6類NAC蛋白的系統進化分析
干旱處理后,FtNAC17的表達量呈波浪型變化。推測在響應干旱脅迫時,FtNAC17可能與多個基因協同響應該脅迫。低溫處理后,FtNAC17的表達量上調。鹽脅迫處理后,FtNAC17顯著上調表達。茉莉酸甲酯處理后,FtNAC17呈上調表達。脫落酸處理后,FtNAC17呈顯著上調表達。赤霉素處理后,FtNAC17呈顯著上調表達。
3 討論
轉錄因子在植物生長過程中具有重要調控的作用,分析鑒定相關的轉錄因子,并揭示其分子機制一直是分子生物學領域的焦點。NAC轉錄因子是植物特異的轉錄因子,也是植物中數量最大的轉錄因子,所以研究其家族成員的結構與功能具有重要意義。本研究鑒定了苦蕎FtNAC17,并對其進行了生物信息學分析。通過對FtNAC17編碼的蛋白分析可知,該蛋白為親水蛋白,與王洋等[28]結果一致。蛋白質的二級結構是其整個空間結構的基礎,在蛋白質折疊過程的早期形成,為蛋白質三級結構的形成奠定基礎,對亞細胞定位有一定作用。預測FtNAC17蛋白的二級結構發現,無規卷曲在蛋白質二級結構中占據最大比例,使蛋白質構型多樣化,這一點與黎幫勇等[29]研究結果相一致。根據NAC轉錄因子保守NAM結構域,可將植物中NAC轉錄因子分為6個大類,同一大類的NAC基因具有相近的功能,其中響應非生物脅迫的NAC基因都歸屬于SNAC大類。本文克隆的FtNAC17,經與擬南芥TAIR蛋白庫比對,與擬南芥中SNAC類中ANAC002(ATAF1)最為相近。過表達ANAC002(ATAF1)的植物敏感性增強,表明ANAC002(ATAF1)響應病原和非生物脅迫有關[30]。因此,推測親緣關系較近的FtNAC17參與響應非生物脅迫。通過研究干旱、低溫、鹽和激素等非生物脅迫下FtNAC17的表達情況發現,該基因對干旱、低溫、鹽、茉莉酸甲酯、脫落酸和赤霉素脅迫處理均有不同程度的響應,這表明FtNAC17確實參與響應非生物脅迫。干旱處理時,FtNAC17的表達量呈先到達峰值后下降后再上升的一個趨勢,推測該基因在響應這些脅迫時并非獨自發揮作用,很可能與其他基因共同發揮作用。目前,本研究僅就單一脅迫影響FtNAC17表達進行研究,未在多脅迫層面上研究該基因的響應情況。因此,后續可以將FtNAC17響應多脅迫情況進行量化,更深層次的挖掘FtNAC17的功能。
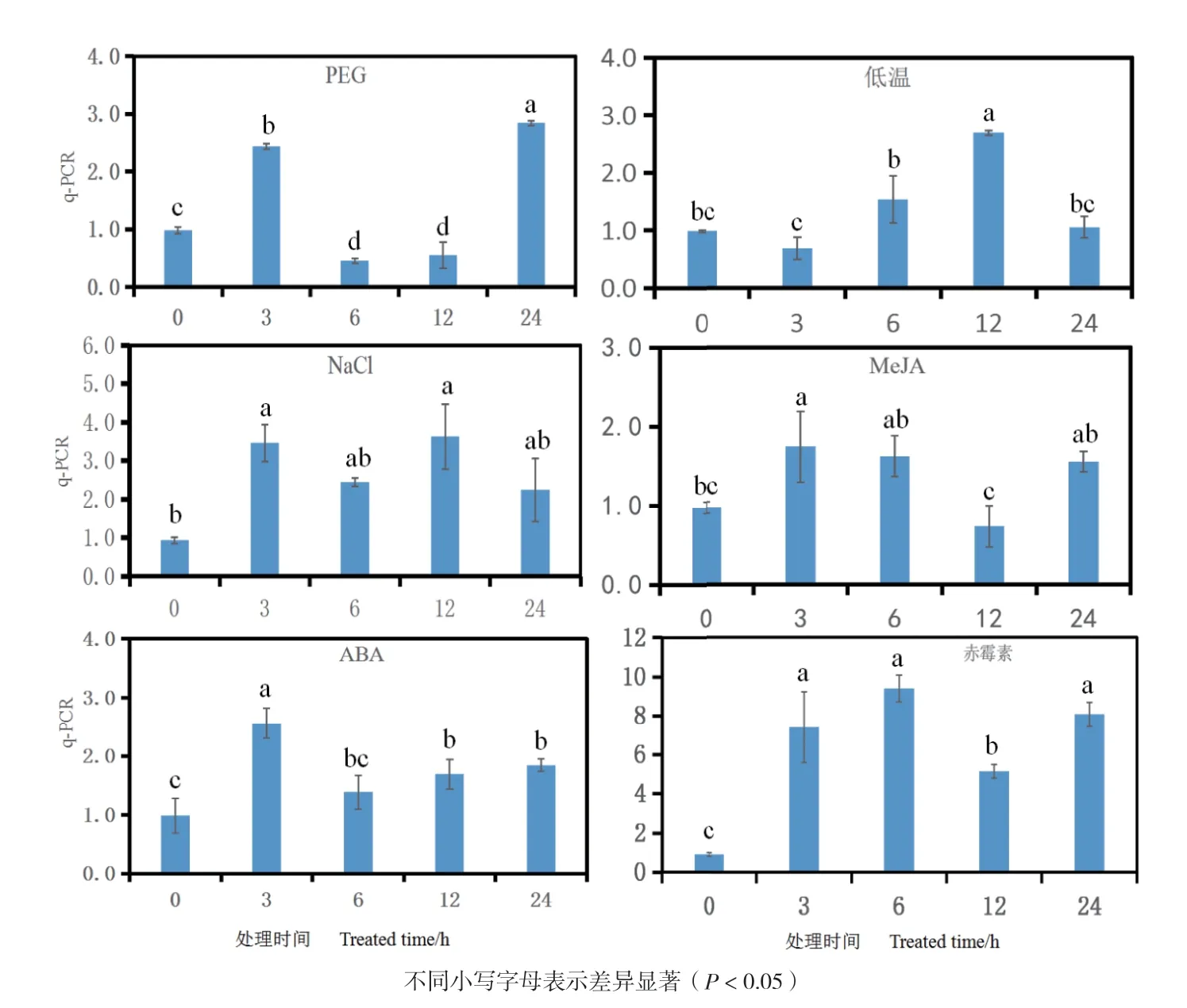
圖7 FtNAC17在不同脅迫下的表達情況
4 結論
初步明確FtNAC17參與應對部分非生物脅迫反應。并且FtNAC17可能通過ABA和MeJA信號通路響應非生物脅迫。
猜你喜歡
中等數學(2022年2期)2022-06-05 07:10:50
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
青年歌聲(2018年5期)2018-10-29 03:18:40
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
廣東飼料(2016年3期)2016-12-01 03:43:12
林業與生態(2016年2期)2016-02-27 14:24:01
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
