葉綠素結合蛋白CP24介導光照響應基因StRSM 1調控葉綠素積累
2021-01-22 09:09:08撒世娟伍涵宇張曉萍鄭蕊姚新靈
生物技術通報 2021年1期
撒世娟 伍涵宇 張曉萍 鄭蕊,2 姚新靈,2
(1. 寧夏優勢特色作物現代分子育種重點實驗室 寧夏大學,銀川 750021;2. 西部特色生物資源保護及利用教育部重點實驗室 寧夏大學,銀川 750021)
擬南芥基因組注釋中最早識別了RSM(RADIALIS-like SANT/MYB)的存在,RSM僅含有單個MYB結構域,這是其不同于其它MYB轉錄因子的組成特征[1],在已測序的植物基因組中均有RSM基因;在比較擬南芥和水稻MYB轉錄因子進化關系研究中,就RSMR是否屬于MYB基因家族存在不同觀點[1-2]。
早期有研究將擬南芥中含有單個MYB結構域的4個RSM蛋白(RSM 1、RSM 2、RSM 3和RSM 4)歸為單MYB轉錄因子家族;研究顯示,RSM 1(AT4G39250)過量表達幼苗生長頂端彎鉤缺失,下胚軸變短,早期光形態建成時對紅光過敏;推測RSM 1參與早期光形態建成調控[3];RSM 1參與了光照誘導頂端分生組織的細胞分裂調控和形態建成。
上調水稻(Oryza sativa)OsRL3(RADIALIS-LIKE3)表達,加速葉片黑暗下衰老;osrl3突變體黑暗下葉片葉綠素滯留,光合能力增強,葉綠素降解相關基因表達水平低于野生型[4]。水稻OsRL3反向調控葉綠素代謝,但調控機理未知。
研究已明確,一定數量的葉綠素等色素分子與葉綠素結合蛋白24(CP24)結合,保持其穩定性和活性;弱光下更多葉綠素與CP24結合,行使吸收光子功能,強光下更多胡蘿卜素與CP24結合,發揮光保護作用,黑暗下葉綠素和CP24積累迅速減少[5-11]。CP24響應光照和黑暗直接參與葉綠素代謝,哪些基因參與CP24表達尚未見報道。
擬南芥RSM1參與其光形態建成和水稻OsRL3調控葉綠素代謝的結果,明確了RSM是光照和黑暗響應基因;但轉錄因子RSM1如何響應光照和黑暗變化有待研究確定。為了確定轉錄因子RSM1決定的表型及響應光照和黑暗變化的路徑,本研究基于反向遺傳學思路,以功能未知的馬鈴薯(Solanum tuberosumL.)基因StRSM1為研究對象,運用基因轉化、陽性轉化株系表型測定和目的基因表達定性測定等方法,揭示了StRSM1通過調控CP24表達及葉綠素積累響應光照和黑暗變化。
1 材料與方法
1.1 材料
供試材料為實驗室自行保存和維護的普通煙草(Nicotiana tabacumL.)K326和馬鈴薯品種紫花白組培苗,以其作為野生型對照和目的基因轉化受體;根癌農桿菌(Agrobacterium tumefaciens)菌株GV3101、大腸桿菌重組菌BL-pC-35S均由本實驗室保存,委托上海生工生物科技公司合成本研究所用引物,其系列如表1所示。

表1 引物序列
轉化株系及WT室內培養條件為(24±2)℃,10 h光照,14 h黑暗,光照強度為100 μmol·m-2·s-1,相對濕度60%-80%,土壤水含量70%±5%。采集移栽生長第30天的幼葉置于液氮,用于RNA提取。
1.2 方法
1.2.1 重組StRSM1構建和體內轉化 委托上海生工生物科技公司合成StRSM1基因全長ORF cDNA,完成重組載體pC-StRSM1+和pC-StRSM1-構建,構建過程如圖1所示。電激轉化重組載體于根癌農桿菌菌株GV3101,卡娜霉素和潮霉素抗性篩選和SRSMF/R引物PCR鑒定,獲得重組菌株GV-StRSM1+和GV-StRSM1-;分別按照已報道的外源基因馬鈴薯(Solanum tuberosum)和本氏煙草(Nicotiana benthamiana)體內轉化方法[12],重組菌侵染外植體、愈傷誘導、陽性苗GUS染色選擇后,用特異引物SRSMF/SRSMR和葉片RNA建立RT-PCR反應,確認陽性株系StRSM1表達,依此獲得本氏煙草和馬鈴薯體內StRSM1過量和抑制表達株系,兩者轉化株系中,選取StRSM1過量和抑制表達株系各2株進行以下實驗。
用引物qRSMF/R建立PCR反應體系,以L25為內參基因,完成RSM1 mRNA積累的實時熒光定量PCR測定,反應條件為95℃ 5min預變性后,95℃ 15 s,62℃ 30 s,45個循環,熔解曲線65℃-95℃;3次樣品重復測定3次,用比較Ct值法計算StRSM1相對表達量。
1.2.2 目的株系生理表型測定 用日本KONICA MINOLTO公司生產的SPAD-502 Plus型葉綠素儀,測定普通煙草WT和其轉化株35 d幼苗全部葉片及馬鈴薯WT和其轉化株40 d幼苗全部葉片葉綠素含量,測定重復3次,每次測定重復3次,以SPAD作為葉綠素含量計量單位。
1.2.3StRSM1過量表達而變化的葉綠素代謝及結合蛋白的基因識別 室內培養條件下,設定14 h光照(CL)和10 h黑暗(CD)作為對照光照,8 h光照(SL)和16 h黑暗(LD)作為短光照處理,在對照和處理光照下,培養StRSM1過量表達株系35 d幼苗5 d,幼苗苗齡40 d時,采集其葉片樣品3份,用上述同樣方法,提取和處理葉片RNA,反轉錄獲得cDNA。
基于StRSM1過量表達改變了葉綠素積累,檢索馬鈴薯基因組數據庫,獲得參與葉綠素代謝和結合的基因cDNA序列,依據序列設計葉綠素代謝和結合的基因特異引物,用光照處理葉片RNA和合成的特異性引物,建立RT-PCR反應。依據StRSM1過量表達株系光照處理樣品RT-PCR產物的變化,確定StRSM1過量表達變化的葉綠素代謝和葉綠素結合蛋白的基因。
其中,依據馬鈴薯基因組數據庫記載的CP24(ID:DMG400012591)DNA序列設計并合成其特異引物612F/R,以L25為內參基因完成PCR,反應條件為95℃預變性3 min、94℃變性30 s、50℃退火35 s、72℃延伸1min的條件下循環25次后,72℃延伸10 min,4℃保存;反應產物用1%瓊脂糖膠電泳觀察不同處理下CP24定性表達結果。
1.2.4 數據分析 用Statistical Program for Social Sciences(SPSS)V19.0中t檢驗完成所有測定數據的統計分析。
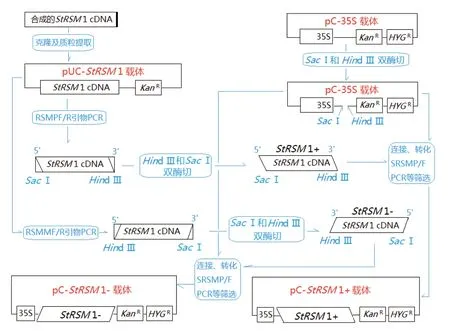
圖1 載體pC-StRSM 1+和pC-StRSM 1-構建過程示意圖
1.2.5 序列檢索及分析 用茄科基因組數據庫https://solgenomics.net/和馬鈴薯基因組數據庫http://solanaceae.plantbiology.msu.edu/、Uniprot數據庫https://www.uniprot.org檢索相關序列、clustal Omega在線http://www.clustal.org/軟件序列比對分析。
2 結果
2.1 StRSM 1及其茄科同源物具有茄科特異性
本實驗室前期研究光照周期處理馬鈴薯,轉錄組測序識別了StRSM1和StRSM1含有255 bp的開放閱讀框,編碼84個氨基酸組成的蛋白質;預測氨基酸序列檢索馬鈴薯基因組數據庫,識別了4個與StRSM1一致性大于56%的氨基酸序列,比對結果如圖2-A所示,其中與StRSM1氨基酸序列最接近的DMP400001385(DMG400000709)一致性僅為65%;與StRSM1同源的擬南芥序列為蛋白質RADIALIS-like 1(AT4G39250),兩者氨基酸序列比對如圖2-B所示,兩者一致性僅為69%。
番茄(Solanum lycopersicum)、野生煙草(Nicotianaattenuata)、普通煙草(Nicotiana tabacumL.)和甜椒(Capsicum annuum)等茄科植物的StRSM1同源物序列比對,如圖2-C所示,其一致性大于80%,且功能結構域高度保守。

圖2 StRSM 1與不同同源物氨基酸序列比對
檢索結果發現,與StRSM1同源的擬南芥序列一致性較低,而其茄科同源序列高度保守,表明StRSM1及其茄科同源基因具有茄科特異性,StRSM1是馬鈴薯基因組中的單拷貝基因,其功能有待確定。
2.2 StRSM 1表達反向調控葉綠素積累
以pC-35S和StRSM1 cDNA構建其過量和抑制表達載體pC-StRSM1+和pC-StRSM1-,載體酶切分析和測序證實后,分別轉化普通煙草和馬鈴薯基因組,依據報告基因GUS染色和RSM1表達定量結果進行選擇,獲得了RSM1過量表達普通煙草陽性轉化株系ntrsm+1和ntrsm+2、抑制表達陽性轉化株系ntrsm-1和ntrsm-2,馬鈴薯過量和抑制表達StRSM1株系strsm+1和strsm+2及strsm-1和strsm-2。
RSM1表達RT-qPCR相對定量結果如圖3-A和3-B所示,過量表達株系ntrsm+1和ntrsm+2及strsm+1和strsm+2 中RSMmRNA積累倍數極顯著高于WT(P≤0.01),表明RSM+cDNA體內表達促進了NtRSM1和StRSM1同源mRNA表達;抑制表達陽性株系ntrsm-1和ntrsm-2及strsm-1和strsm-2葉片RSMmRNA積累倍數極顯著低于WT(P≤0.01),表明RSM-cDNA體內表達抑制了NtRSM1和StRSM1同源mRNA表達。
普通煙草和馬鈴薯葉片過量和抑制表達StRSM1,改變了兩者葉片顏色,如圖3-C所示,與WT相比,過量StRSM1表達馬鈴薯陽性株系葉色較淺,但抑制StRSM1表達馬鈴薯陽性株系葉片綠色明顯加深。
葉綠素含量(SPAD)測定結果如圖3-D和3-E所示,普通煙草和馬鈴薯StRSM1抑制表達陽性株系ntrsm-1和ntrsm-2及strsm-1和strsm-2葉片葉綠素含量極顯著高于對照(P≤0.01),ntrsm+1和ntrsm+2及strsm+1和strsm+2株系葉片葉綠素含量極顯著低于對照(P≤0.01);結果表明,StRSM1過量表達減少了葉綠素積累;抑制StRSM1表達,葉綠素積累則會增加;抑制StRSM1表達的結果,與已報道的RSM3突變體倒置使黑暗誘導的葉綠素減少的研究結果一致。
2.3 隨StRSM表達變化而改變的葉綠素代謝和葉綠素結合蛋白基因
檢索數據庫識別了72個葉綠素代謝和葉綠素結合蛋白(CMB)編碼基因,用這些基因各自特異引物進行RT-PCR,依此作為CMB基因表達定性結果,比較對照和光照處理條件下StRSM1過量表達株系中CMB基因隨黑暗和光照的表達變化,確定隨StRSM1過量表達而改變表達的CMB基因。
RT-PCR結果顯示,72個CMB基因中,葉綠素合成酶、葉綠素b還原酶、葉綠素酶、原葉綠素氧化還原酶等編碼基因表達在處理和對照間無可見穩定變化,其中葉綠素結合蛋白CP24基因在光照處理和對照間表現穩定變化。
如圖4所示,在馬鈴薯和普通煙草的野生型(WT)中,RT-PCR僅檢測到CP24在對照光照(CL)條件下表達,對照黑暗(CD)未檢出可見產物;StRSM1過量表達馬鈴薯和普通煙草株系中,CL和CD條件下均獲得明顯可見的RT-PCR產物;StRSM1過量表達株系在LD和SL條件下,CP24呈現與CL和CD條件下相同的表達模式。結果表明,StRSM1過量表達改變了CP24的表達模式,即增加了黑暗條件下CP24的表達量。

圖4 隨RSM 1過量表達的CP24基因RT-PCR產物
3 討論
早期許多研究報道了光照對葉綠素積累的影響[13-14],其后的研究就此得到的基本結論認為,強光條件下,葉綠體中類胡蘿卜素組分積累增加,發揮光保護作用;弱光條件下,葉綠素積累相對增加;黑暗下,葉綠素和葉綠素結合蛋白積累迅速減少[5-6]。不同植物色素吸收光譜不同,光照強弱變化改變了LHC中的色素組成。
葉綠體和類胡蘿卜素合成均來自類異戊二烯(MEP)途徑,兩者有共同的合成底物[15-16];弱光條件下,葉綠素積累增加[5-6];盡管需更多實驗證據支持,但依此有理由推測,高光照誘導類胡蘿卜素等光保護色素積累增加,其合成消耗了更多合成底物,葉綠素合成相對減少;弱光下,光保護色素合成減少,更多合成底物用于葉綠素合成,葉綠素相對增加。RSM1表達減少增加葉綠素積累的結果與弱光條件下葉綠素積累增加一致,RSM1表達減少促進葉綠素積累的結果也表明,RSM1反向調控葉綠素積累。該結果與已報道的水稻RSM3突變體倒置使黑暗誘導葉綠素減少的結果一致。
葉綠素結合蛋白CP具有聚集和結合色素及傳遞電子等功能,色素聚集并結合于CP[9],維持色素活性和可測狀態,黑暗下葉綠素和CP24積累迅速減少[11-14],這些結果同時也表明,除了不同色素合成平衡響應光照外,CP在響應光照變化中發揮更重要作用。本研究黑暗(CD)下未檢測到WT中CP24存在,證實了黑暗下CP24積累迅速減少;StRSM1過量表達株系中CD和LD下CP24存在的原因是StRSM1過量表達,結果明確了StRSM1參與CP24積累。
天線蛋白聚集色素形成光子吸收和電子傳遞復合體,光照調控色素與其結合蛋白積累變化,維持復合體光保護和光合的動態;天線蛋白結構及功能的研究已較多,但其如何受到上游基因表達調控還不是很清楚。
本研究初步揭示了StRSM1隨光照調控CP24表達,繼而改變葉綠素積累;該結果建立了光照、RSM1轉錄因子、CP24和葉綠素積累間的聯系,結果對于進一步明確RSM1如何響應光照和深刻理解RSM1參與的光照響應具有作用。
StRSM1及其茄科同源基因均含有高度保守的單個MYB結構域,StRSM1在馬鈴薯基因組中是單拷貝,其與擬南芥同源基因相似性較低,這表明了其差異進化的特征;單個MYB結構域重復形成了其它MYB轉錄因子的事實,表明了StRSM1是更早出現在基因組中的基因,StRSM1功能的初步確定有助于揭示光合作用這一古老生理現象。
研究中發現,過量表達RSM1,增加了馬鈴薯和普通煙草體內光照和黑暗下CP24 mRNA的積累,尤其是黑暗下CP24積累增加明顯可見;普通煙草轉化株系中,短光照(SL)和對照光照(CL)下未出現CP24 mRNA積累的可見差異,但長黑暗(LD)下,CP24 mRNA積累明顯更多,其原因有待進一步研究。
4 結論
本研究發現,茄科植物特異單MYB轉錄因子StRSM1光照響應,反向調控葉綠素積累,葉綠素結合蛋白CP24參與了StRSM1對葉綠素積累的調控。
