植物對(duì)低溫脅迫的生理響應(yīng)及外源脫落酸緩解脅迫效應(yīng)的研究進(jìn)展
2021-01-27 03:25:50項(xiàng)洪濤鄭殿峰何寧李琬王曼力王詩(shī)雅
草業(yè)學(xué)報(bào) 2021年1期
關(guān)鍵詞:植物
項(xiàng)洪濤,鄭殿峰,何寧,李琬,王曼力,王詩(shī)雅
(1.黑龍江省農(nóng)業(yè)科學(xué)院耕作栽培研究所,黑龍江 哈爾濱150086;2.廣東海洋大學(xué)農(nóng)學(xué)院,廣東 湛江524088;3.黑龍江八一農(nóng)墾大學(xué)農(nóng)學(xué)院,黑龍江 大慶163319)
植物在自然界中生長(zhǎng)和生存,其生育進(jìn)程中不可避免地會(huì)受到低溫、干旱、鹽堿等極端環(huán)境因子的影響,其中溫度對(duì)植物生長(zhǎng)發(fā)育影響較大,甚至是導(dǎo)致植物死亡的關(guān)鍵環(huán)境因子[1]。低溫作為重要的環(huán)境因子影響著植物的地理分布和物種多樣性,并且對(duì)農(nóng)業(yè)可持續(xù)發(fā)展造成了嚴(yán)重的威脅[2]。在全球氣候變暖的背景下,氣溫變化卻比較波動(dòng),極端天氣頻繁出現(xiàn),作物的低溫、寒凍災(zāi)害日趨嚴(yán)重,導(dǎo)致產(chǎn)量降低、品質(zhì)下降[3],且嚴(yán)重時(shí)可引起植株死亡,從而給農(nóng)業(yè)生產(chǎn)造成巨大損失[4]。
我國(guó)農(nóng)業(yè)生產(chǎn)歷史悠久,勞動(dòng)人民智慧地總結(jié)出了多種多樣的抵御低溫的方法,比如熏煙法、培育抗冷品種、對(duì)植物進(jìn)行低溫馴化等不同形式的物理學(xué)、育種學(xué)和栽培學(xué)等方法,這些方法較為有效但又存在明顯的弊端。通過(guò)挖掘植物自身的抗冷潛力,開(kāi)啟植物體自我保護(hù)機(jī)制是一種重要的抗逆途徑。通過(guò)施用外源激素或植物生長(zhǎng)調(diào)節(jié)劑來(lái)刺激植物理化過(guò)程并提高抗逆能力,已經(jīng)越來(lái)越受到廣泛關(guān)注。近年來(lái)外源脫落酸(abscisic acid,ABA)在我國(guó)農(nóng)業(yè)生產(chǎn)中應(yīng)用越來(lái)越多,因其具有抑制作物生長(zhǎng)的生理效應(yīng),在壯根健苗、促進(jìn)早熟、穩(wěn)產(chǎn)增產(chǎn)以及抗逆減災(zāi)領(lǐng)域中均有報(bào)道,尤其是近年來(lái)分子生物學(xué)的發(fā)展,已經(jīng)明確了植物抗冷過(guò)程中存在著一條明顯的依賴(lài)內(nèi)源ABA 途徑,這間接說(shuō)明了施用外源ABA 可能具有很好的抵御低溫的效應(yīng)。因此,通過(guò)全面總結(jié)植物對(duì)低溫脅迫的應(yīng)激反應(yīng)變化及外源ABA 的緩解效應(yīng),對(duì)尋找提高植物抗冷、耐冷途徑,增強(qiáng)植物抵御低溫的能力、挖掘有效的農(nóng)業(yè)措施具有極大幫助,同時(shí)對(duì)提高農(nóng)業(yè)生產(chǎn)效能也具有重要的現(xiàn)實(shí)意義。
1 植物對(duì)低溫脅迫的生理應(yīng)激響應(yīng)
植物的生長(zhǎng)發(fā)育受內(nèi)外因素的共同影響,外部因素包括光照強(qiáng)度、光周期和溫度變化等,內(nèi)部因素則有激素水平,理化進(jìn)程和信號(hào)傳導(dǎo)等[2]。植物在整個(gè)生命過(guò)程中不斷適應(yīng)外部環(huán)境并通過(guò)信號(hào)傳導(dǎo)調(diào)整內(nèi)外部因素平衡,安全完成一個(gè)完整的生命周期活動(dòng)。但在一個(gè)生命周期內(nèi),植物的發(fā)育進(jìn)程中不可避免地遭受或強(qiáng)或弱的逆境脅迫,比如干旱、風(fēng)雹、水淹、低溫等。低溫對(duì)植物的形態(tài)結(jié)構(gòu)發(fā)展、理化代謝途徑、生長(zhǎng)發(fā)育進(jìn)程都具有較大影響甚至傷害,低溫條件下植物自身會(huì)產(chǎn)生復(fù)雜的生物化學(xué)和生理學(xué)上的應(yīng)激響應(yīng),以應(yīng)對(duì)和適應(yīng)低溫環(huán)境。植物對(duì)低溫脅迫的適應(yīng)性和抵抗能力既受系統(tǒng)發(fā)育的遺傳基因控制,又受個(gè)體發(fā)育過(guò)程中生理狀態(tài)制約[5],植物遭受低溫脅迫后,會(huì)開(kāi)啟一系列應(yīng)激的自我防護(hù)機(jī)制,這些保護(hù)系統(tǒng)之間相互有機(jī)協(xié)調(diào),以最大程度降低傷害,具體如圖1 所示。
1.1 低溫脅迫對(duì)植物光合生理的影響
光合作用是植物生長(zhǎng)最根本的基礎(chǔ),其以二氧化碳(CO2)和水分(H2O)為反應(yīng)底物,通過(guò)較為復(fù)雜的生化過(guò)程生成光合碳同化物,是植物體內(nèi)有機(jī)營(yíng)養(yǎng)和碳架結(jié)構(gòu)的物質(zhì)來(lái)源。Ainsworth 等[6]指出光合作用是植物生長(zhǎng)的物質(zhì)和能量基礎(chǔ),但對(duì)冷害也最為敏感。低溫脅迫對(duì)光合作用的影響很大,主要原因是低溫能夠通過(guò)光抑制影響光合系統(tǒng),嚴(yán)重的低溫脅迫還會(huì)導(dǎo)致光氧化被破壞,再者低溫會(huì)降低植物體內(nèi)的光合作用酶活性,從而影響光合作用[7]。
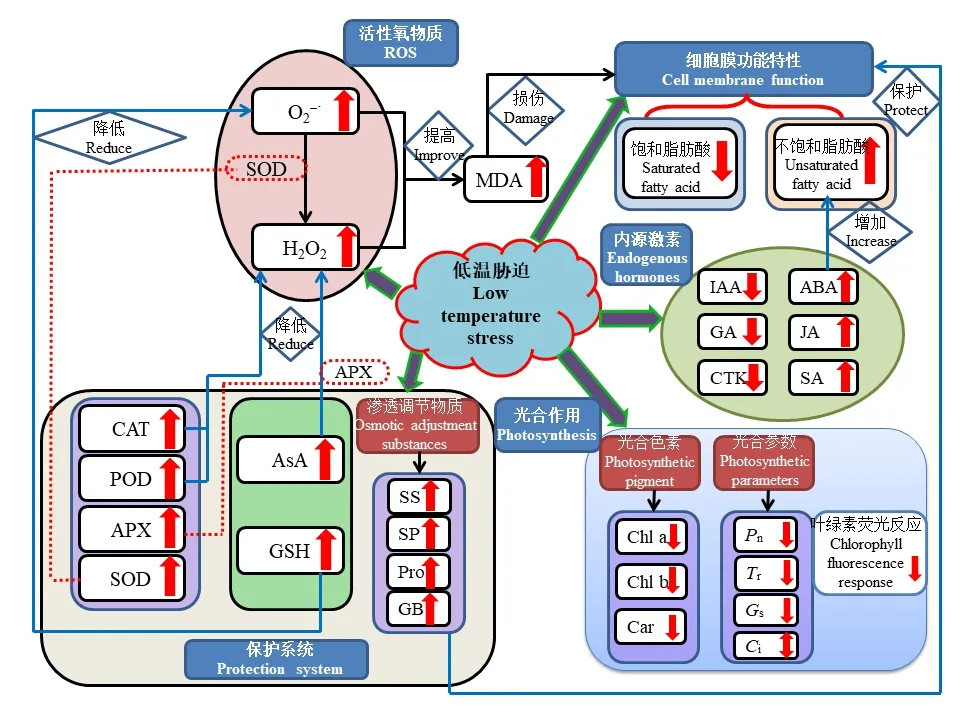
圖1 低溫脅迫對(duì)植物生理代謝的影響Fig.1 Effect of low temperature stress on plant physiological metabolism
1.1.1 低溫脅迫對(duì)植物光合色素的影響 葉綠素和類(lèi)胡蘿卜素參與光合作用,涉及光能的吸收、傳遞和轉(zhuǎn)化的相關(guān)過(guò)程。植物葉綠素包括葉綠素a 和葉綠素b,一份在光反應(yīng)中心負(fù)責(zé)將光能轉(zhuǎn)變?yōu)榛瘜W(xué)能,其余葉綠素則負(fù)責(zé)光能的捕獲和傳遞[8]。低溫脅迫可以破壞葉綠體結(jié)構(gòu),使葉綠素生物合成受阻[9],引起葉片內(nèi)葉綠素a、葉綠素b 和總?cè)~綠素含量降低、植株黃化[10]。楊碧云等[11]研究表明低溫脅迫達(dá)到一定強(qiáng)度時(shí),葉綠素a、葉綠素b、葉綠素(a+b)含量、葉綠素a/b 值均呈下降趨勢(shì)。類(lèi)胡蘿卜素在植物體內(nèi)可起到保護(hù)葉綠素類(lèi)免受逆境因子影響導(dǎo)致光氧化破壞的作用,低溫條件下類(lèi)胡蘿卜素含量也發(fā)生明顯變化,有報(bào)道指出類(lèi)胡蘿卜素含量隨著處理溫度的下降和處理時(shí)間延長(zhǎng)而降低[12]。周真珍等[7]表明低溫脅迫降低了植物葉片內(nèi)葉綠素含量,并明確指出兩個(gè)原因:一是因?yàn)榈蜏亟档土巳~綠素合成酶活性,使葉綠素生物合成受阻,二是原有葉綠體色素氧化分解,引起葉綠素含量減少。葉綠素含量的減少不利于葉綠體對(duì)光能的吸收利用[13],影響光能捕獲,光合電子傳遞受阻[14],導(dǎo)致光合能力下降,碳同化受到抑制,植株生長(zhǎng)遲緩,產(chǎn)量與質(zhì)量下降[4]。
1.1.2 低溫脅迫對(duì)植物光合參數(shù)的影響 低溫直接影響植物光合器官的結(jié)構(gòu)和活性,也間接影響其光合作用,光合參數(shù)可直接反映其光合作用的強(qiáng)弱[15]。光合參數(shù)主要包含以下4 項(xiàng)指標(biāo):氣孔導(dǎo)度(Gs)、細(xì)胞間CO2濃度(Ci)、葉片凈光合速率(Pn)以及蒸騰速率(Tr)。氣孔是植物與大氣進(jìn)行CO2和水汽交換的通道,植物在光下進(jìn)行光合作用,通過(guò)氣孔吸收CO2,氣孔導(dǎo)度的高低會(huì)影響光合作用[16]。CO2是植物進(jìn)行光合作用的底物,細(xì)胞間CO2濃度影響植物的光合速率[7]。而凈光合速率是植物光合系統(tǒng)功能的直接體現(xiàn),也是植物光合系統(tǒng)工作正常與否的判斷依據(jù)[7]。有報(bào)道指出低溫導(dǎo)致光合參數(shù)受到顯著抑制,降低葉片Pn、Gs以及Tr[17],楊晶等[18]也指出隨著低溫脅迫時(shí)間的延長(zhǎng),Pn、Gs以及Tr會(huì)持續(xù)下降,這與崔波等[19]的研究相同。但不同研究中Ci的變化趨勢(shì)卻不完全相同,鄭春芳等[17]指出低溫使Ci顯著增加,楊晶等[18]的研究結(jié)論是Ci會(huì)小幅下降,而崔波等[19]的研究表明Ci先下降后上升。光合參數(shù)之間進(jìn)行協(xié)同工作,共同抵御低溫對(duì)光合作用的影響。低溫對(duì)光合作用最明顯的影響就是Pn的降低,F(xiàn)arquhar 等[20]研究認(rèn)為,低溫導(dǎo)致光合速率下降的因素既有氣孔因素也有非氣孔因素。如果Pn、Gs及Tr同時(shí)下降,Ci也相應(yīng)下降,即氣孔因素占主導(dǎo)[19];如果Pn、Gs及Tr下降的同時(shí)Ci升高,則是由非氣孔因素阻礙CO2的利用,造成Ci的積累所致[21]。
1.1.3 低溫脅迫對(duì)植物葉綠素?zé)晒夥磻?yīng)的影響 葉綠素?zé)晒庑盘?hào)由植物體內(nèi)發(fā)出,并且具有豐富的光合作用信息,這與植物受脅迫程度相關(guān)。低溫脅迫影響光能的吸收、轉(zhuǎn)換與光電子傳遞,致使過(guò)剩的激發(fā)能大量積累于光合系統(tǒng)Ⅱ(PSⅡ)反應(yīng)中心,進(jìn)而損傷植物葉片的PSⅡ反應(yīng)中心,并導(dǎo)致葉片光合作用能力減弱[22]。有報(bào)道指出,短時(shí)低溫脅迫下,植物葉片光合色素含量和構(gòu)成比例幾乎沒(méi)有變化,但是葉片PSⅡ的有效光化學(xué)效率(Fv′/Fm′)和實(shí)際光化學(xué)效率(ΦpsⅡ)出現(xiàn)明顯降低,光化學(xué)淬滅系數(shù)(qP)的變化也相似,而葉片的非光化學(xué)淬滅系數(shù)(NPQ)迅速增大[8]。劉蕾蕾等[23]對(duì)小麥(Triticum aestivum)的研究表明隨著低溫持續(xù)時(shí)間的延長(zhǎng),葉片最大光化學(xué)效率(Fv/Fm)、實(shí)際光化學(xué)效率(ΦpsⅡ)呈先下降后上升的趨勢(shì),并指出一般在低溫脅迫2~5 d,葉片ΦpsⅡ降至最低。葉片光化學(xué)猝滅系數(shù)(qP)隨低溫持續(xù)時(shí)間的延長(zhǎng)呈先下降后升高趨勢(shì),并且隨溫度的降低呈下降趨勢(shì),而葉片的非光化學(xué)淬滅系數(shù)(NPQ)與低溫持續(xù)時(shí)間關(guān)系不大,但隨著低溫強(qiáng)度加劇呈較為顯著的上升趨勢(shì)。
1.2 低溫脅迫對(duì)植物逆境生理的影響
低溫脅迫通常會(huì)導(dǎo)致植物的形態(tài)和生理生化發(fā)生變化,其對(duì)細(xì)胞內(nèi)部的生理改變主要有以下4 個(gè)方面:活性氧類(lèi)物質(zhì)積累、膜質(zhì)過(guò)氧化物含量變化、抗氧化酶活性及非酶抗氧化劑含量變化以及滲透調(diào)節(jié)物質(zhì)含量變化。一般來(lái)說(shuō)低溫脅迫可以引發(fā)植物體內(nèi)活性氧(reactive oxygen species,ROS)類(lèi)物質(zhì)如過(guò)氧化氫(H2O2)、超氧陰離子(O2—·)等物質(zhì)的積累,從而增加植物的氧化應(yīng)激反應(yīng)。大量的氧自由基對(duì)細(xì)胞膜傷害很大,引起膜質(zhì)過(guò)氧化反應(yīng),產(chǎn)生大量的丙二醛(MDA),MDA 能夠抑制細(xì)胞保護(hù)酶活性,從而加劇膜質(zhì)過(guò)氧化,同時(shí)其本身也是具有細(xì)胞毒性的物質(zhì),它能夠和酶蛋白結(jié)合、交聯(lián),從而使保護(hù)酶失去活性,也就進(jìn)一步破壞膜結(jié)構(gòu)。一定脅迫程度內(nèi),抗氧化酶體系和非酶抗氧化劑能夠有效清除ROS 并消除它們的不利影響。同時(shí)植物應(yīng)激低溫脅迫也會(huì)帶來(lái)滲透調(diào)節(jié)物質(zhì)含量的變化,如脯氨酸、甜菜堿等物質(zhì)會(huì)有所提高,來(lái)維持細(xì)胞膜的滲透調(diào)節(jié)功能,從而增強(qiáng)植物體對(duì)低溫脅迫的抵抗能力[24]。
1.2.1 低溫脅迫對(duì)植物活性氧物質(zhì)積累及膜質(zhì)過(guò)氧化的影響 植物代謝所產(chǎn)生的ROS 包括超氧陰離子(O2—·)、氫氧根離子(OH-)、羥基自由基(?OH)、過(guò)氧化氫(H2O2)等[25]。活性氧作為植物信號(hào)轉(zhuǎn)導(dǎo)中第二信使,在植物生長(zhǎng)發(fā)育中起重要作用[26]。正常條件下,植物能夠維持自身ROS 平衡,但當(dāng)植物暴露于低溫環(huán)境下,細(xì)胞穩(wěn)態(tài)被破壞并產(chǎn)生過(guò)量ROS,如果超量的ROS 未被及時(shí)清除便積累在植物體內(nèi),會(huì)加劇植物細(xì)胞膜質(zhì)過(guò)氧化傷害,并對(duì)細(xì)胞膜系統(tǒng)產(chǎn)生損害[27],干擾作物的生理生化過(guò)程,最終制約作物產(chǎn)量和品質(zhì)[28]。李海燕等[29]研究表明低溫脅迫使O2—·產(chǎn)生速率和H2O2含量顯著增加,并指出低溫時(shí)間越長(zhǎng),增加趨勢(shì)越明顯。張愛(ài)冬等[30]也指出隨著溫度降低,植物葉片O2—·產(chǎn)生速率和H2O2含量均呈上升的趨勢(shì),這促使細(xì)胞膜透性增大,胞內(nèi)電解質(zhì)外滲,加劇膜質(zhì)過(guò)氧化作用。MDA 是細(xì)胞膜過(guò)氧化產(chǎn)物,其含量高低是質(zhì)膜受損嚴(yán)重與否的重要指標(biāo)[31],MDA 含量與植物耐寒性間呈負(fù)相關(guān),脅迫程度越強(qiáng),MDA 積累量越多。項(xiàng)洪濤等[32]指出低溫導(dǎo)致植物體內(nèi)MDA 含量增加,并隨脅迫強(qiáng)度和脅迫時(shí)間的增加而增多。鄧化冰等[33]也指出低溫能夠?qū)е轮参锶~片MDA 含量顯著增加,加劇膜質(zhì)過(guò)氧化程度,導(dǎo)致膜結(jié)構(gòu)破壞,質(zhì)膜透性加大,電解質(zhì)外滲,從而影響葉片的生理生化機(jī)能。
1.2.2 低溫脅迫對(duì)植物抗氧化系統(tǒng)的影響 植物在自然選擇中進(jìn)化出多種ROS 清除機(jī)制來(lái)進(jìn)行自我防護(hù),通過(guò)這些機(jī)制來(lái)響應(yīng)氧化應(yīng)激。植物體內(nèi)這些抗氧化機(jī)制包括保護(hù)酶系統(tǒng)和非酶抗氧化劑[24]。當(dāng)遭遇逆境脅迫時(shí),植物會(huì)自動(dòng)開(kāi)啟保護(hù)系統(tǒng)抵御逆境因子帶來(lái)的威脅,進(jìn)而盡量維持正常的生理代謝活動(dòng),免受損害。保護(hù)酶系統(tǒng)主要包括超氧化物歧化酶(SOD)、過(guò)氧化物酶(POD)、過(guò)氧化氫酶(CAT)和抗壞血酸過(guò)氧化物酶(APX)。正常情況下,這些酶之間相互配合、銜接,能夠自動(dòng)氧化體內(nèi)不斷產(chǎn)生的活性氧類(lèi)物質(zhì),這些物質(zhì)的產(chǎn)生和清除處于一種動(dòng)態(tài)平衡的狀態(tài),所以植物不會(huì)受到傷害。低溫脅迫起初誘發(fā)抗氧化酶活性上升,但隨著脅迫加劇,這些抗氧化酶活性下降,嚴(yán)重的低溫脅迫積累了過(guò)量的活性氧,使活性氧和防御系統(tǒng)的動(dòng)態(tài)平衡遭到了破壞,從而加劇膜質(zhì)過(guò)氧化作用,產(chǎn)生大量的MDA,進(jìn)而導(dǎo)致膜系統(tǒng)受到損傷使得植物受到傷害。植物通過(guò)SOD、POD 和CAT 三者協(xié)同作用,使體內(nèi)的氧自由基維持在較低水平,可以在一定程度上減緩或防御低溫脅迫。項(xiàng)洪濤等[34]研究指出低溫可導(dǎo)致植物體內(nèi)SOD、POD 和CAT 活性先升高后下降,短時(shí)期內(nèi)可清除多余的ROS,但隨著低溫脅迫時(shí)間的延長(zhǎng),這種保護(hù)機(jī)能逐漸下降甚至崩潰,這與李春燕等[35]的報(bào)道相似,其指出低溫處理一段時(shí)間后保護(hù)酶活性表現(xiàn)出持續(xù)下降的趨勢(shì),并表明這種持續(xù)下降的現(xiàn)象是低溫對(duì)作物造成了不可恢復(fù)的傷害。非酶抗氧化劑主要有抗壞血酸(AsA)和谷胱甘肽(GSH),AsA 具有清除H2O2、OH-等功能,而GSH 是重要的活性物質(zhì),其能夠幫助保持免疫系統(tǒng)不被破壞,并且具有抗氧化作用和整合解毒作用,防止細(xì)胞膜過(guò)氧化,從而保證細(xì)胞膜完整并行使正常功能[4]。有報(bào)道指出,隨著低溫強(qiáng)度的增加,AsA 和GSH 都呈先增加后降低的變化趨勢(shì)[36],說(shuō)明達(dá)到一定低溫閾值后,即超出了AsA?GSH 循環(huán)的調(diào)節(jié)適應(yīng)能力后,二者含量迅速降低,還原能力下降,植物受害加重。鄭春芳等[17]指出AsA?GSH 循環(huán)是調(diào)控植物響應(yīng)低溫的重要途徑,并表明低溫脅迫降低了AsA 和GSH的含量。
1.2.3 低溫脅迫對(duì)植物滲透調(diào)節(jié)物質(zhì)含量的影響 植物在遭受冷害的同時(shí)還會(huì)受到水分脅迫和滲透脅迫,一些具有保水作用的滲透調(diào)節(jié)物質(zhì),如脯氨酸、甜菜堿、可溶性糖和可溶性蛋白等物質(zhì)會(huì)大量增加,以維持細(xì)胞的滲透平衡,減小低溫傷害[37]。脯氨酸含量與植物脅迫之間存在正相關(guān)關(guān)系,它可以通過(guò)維持細(xì)胞膨脹或滲透平衡來(lái)賦予植物脅迫耐受性,從而穩(wěn)定膜結(jié)構(gòu),防止電解質(zhì)泄露,使活性氧的濃度維持在正常范圍內(nèi),在植物抗脅迫中發(fā)揮著重要作用[24]。植物體內(nèi)可溶性物質(zhì)如可溶性糖和可溶性蛋白,能夠提高細(xì)胞液濃度,增加細(xì)胞持水組織中的非結(jié)冰水,是重要的滲透調(diào)節(jié)物質(zhì),其含量的增加有利于提高植物的抗寒性[5]。甜菜堿(GB)是一種水溶性生物堿,無(wú)毒無(wú)害且廣泛存在于動(dòng)植物中,可以調(diào)節(jié)細(xì)胞滲透壓以維持生物膜及蛋白質(zhì)結(jié)構(gòu)和功能的完整,具有較好的抗逆功能[38]。植物受到低溫脅迫后,體內(nèi)的滲透調(diào)節(jié)物質(zhì)含量都會(huì)有所增加,共同抵御低溫脅迫,維持正常的生理代謝,這在水稻(Oryza sativa)[34]、玉米(Zea may)[24]、紫花苜蓿(Medicago sativa)[38]、豇豆(Vigna unguiculata)[27]、絲瓜(Luffa cylindrica)[30]以及馬鈴薯(Solanum tuberosum)[7]等多種作物中得到廣泛驗(yàn)證。
1.3 低溫脅迫對(duì)植物細(xì)胞膜組分和結(jié)構(gòu)的影響
細(xì)胞膜是植物體細(xì)胞及細(xì)胞器與外界環(huán)境之間進(jìn)行物質(zhì)和能量交換的場(chǎng)所,同時(shí)也是細(xì)胞感應(yīng)外界環(huán)境中各種逆境脅迫信號(hào)的細(xì)胞結(jié)構(gòu)[39]。細(xì)胞膜系統(tǒng)的穩(wěn)定性與植物抗逆性存在密切關(guān)系,其穩(wěn)定性和流動(dòng)性是細(xì)胞生命活動(dòng)的基礎(chǔ),也是形成植物抗逆能力的關(guān)鍵[40]。細(xì)胞膜的結(jié)構(gòu)是一種動(dòng)態(tài)平衡體系,構(gòu)成細(xì)胞膜磷脂雙分子層中的脂肪酸成分及其飽和水平與膜穩(wěn)定性密切相關(guān),細(xì)胞膜的穩(wěn)定性得益于細(xì)胞膜膜質(zhì)的流動(dòng)性[41]。當(dāng)植物細(xì)胞受到低溫脅迫達(dá)到一定的相變溫度時(shí),脂膜會(huì)由流動(dòng)性的液晶相轉(zhuǎn)變?yōu)榻┯驳哪z相,引發(fā)膜質(zhì)的脂肪酸鏈由無(wú)序排列變?yōu)橛行蚺帕校@阻礙了膜的正常生理功能,會(huì)導(dǎo)致膜透性增大,細(xì)胞液外滲,最終導(dǎo)致細(xì)胞死亡[40?42]。膜質(zhì)相變程度與不同脂肪酸含量及比例關(guān)系密切,通常認(rèn)為植物的膜質(zhì)相變溫度降低是由于不飽和脂肪酸含量的增高,進(jìn)而增加了脂肪酸鏈的不飽和程度,不飽和程度越大,膜的流動(dòng)性越大,穩(wěn)定性越好,使植物耐寒性得到提高[39,41]。當(dāng)受到低溫脅迫時(shí),許多植物體內(nèi)會(huì)產(chǎn)生較多不飽和度較高的脂肪酸(亞麻酸、油酸、亞油酸等)來(lái)抵御低溫[4]。相關(guān)報(bào)道也指出隨氣溫下降膜質(zhì)組分中飽和脂肪酸減少,不飽和脂肪酸特別是亞油酸大幅增加,使膜質(zhì)相變溫度下降,對(duì)于保持膜流動(dòng)性和正常生理功能有重要作用[40],類(lèi)似的研究結(jié)果在繡線菊(Spiraea salicifolia)[40]、小麥[42]、結(jié)縷草(Zoysia japonica)[39]等植物中都有報(bào)道。
1.4 低溫脅迫對(duì)植物內(nèi)源激素水平的影響
植物激素是植物生長(zhǎng)發(fā)育的重要調(diào)節(jié)物質(zhì),對(duì)逆境產(chǎn)生響應(yīng)的激素信號(hào)分子,廣泛參加作物生理過(guò)程的調(diào)控[43]。ABA、水楊酸(SA)和茉莉酸(JA)是非生物脅迫反應(yīng)的重要影響因素。ABA 作為一種能夠提高植物抗逆能力的“非生物應(yīng)激激素”,既是植物響應(yīng)非生物脅迫的信號(hào),也是引起植物體內(nèi)適應(yīng)性調(diào)節(jié)反應(yīng)和基因表達(dá)的重要因子,參與非生物脅迫下的調(diào)控應(yīng)答[44]。SA 和JA 介導(dǎo)的信號(hào)傳遞途徑與植物抗性密切相關(guān),細(xì)胞內(nèi)增加的SA 能激活抗氧化酶系統(tǒng),使冷應(yīng)答基因的表達(dá)量上調(diào),從而減少細(xì)胞膜的氧化損傷,增強(qiáng)植物的抗寒能力[45]。JA 作為信號(hào)分子參與調(diào)控植物的生長(zhǎng)發(fā)育及低溫、干旱等非生物脅迫,有報(bào)道指出外源茉莉酸甲酯(methyl jasmonate,MeJA)能夠誘導(dǎo)熱激蛋白家族轉(zhuǎn)錄,通過(guò)增加抗氧化劑合成、降低脂氧合酶活性從而增加植物抵御冷害能力,說(shuō)明JA 信號(hào)途徑參與了植物對(duì)低溫的響應(yīng)與適應(yīng)過(guò)程[46]。生長(zhǎng)素(IAA)、赤霉素(GA)和細(xì)胞分裂素(CTK)是植物體內(nèi)重要的生長(zhǎng)類(lèi)激素,對(duì)植物生長(zhǎng)和發(fā)育起重要作用。其中IAA 和GA 廣泛地參與到植物抗低溫系統(tǒng)中,GA 被認(rèn)為與植物抗寒性有關(guān),但是作用效果不如ABA 明顯[31]。李春燕等[35]研究表明低溫處理能促使植物體內(nèi)ABA 含量增加、GA 含量降低。項(xiàng)洪濤等[34]也表明當(dāng)植株受到低溫脅迫時(shí),生活力下降,內(nèi)源激素水平發(fā)生明顯變化,主要表現(xiàn)為ABA 大量增加,IAA 和GA 含量減少,ABA/GA 和ABA/IAA 提高,抑制作物生長(zhǎng)發(fā)育,降低損耗,各激素之間協(xié)同作用抵御低溫脅迫。而劉學(xué)慶等[31]研究指出,低溫脅迫后,植物體內(nèi)的ABA、IAA 和GA 的含量都呈先增高后降低的變化趨勢(shì),這與植物的品種特性及低溫處理的時(shí)間有關(guān)。
1.5 低溫脅迫對(duì)植物重要冷反應(yīng)基因表達(dá)的影響
植物的抗寒過(guò)程是一個(gè)由多個(gè)轉(zhuǎn)錄因子或多種信號(hào)途徑相關(guān)基因相互平行或交叉協(xié)作,共同調(diào)控的復(fù)雜網(wǎng)絡(luò),最終產(chǎn)生結(jié)構(gòu)和生理物質(zhì)的變化,從而增強(qiáng)植物的抗寒能力[47]。近年來(lái),基因表達(dá)的變化成為植物提高抗冷性理化機(jī)制的研究熱點(diǎn)。低溫引起植物生長(zhǎng)停滯,涉及光合能量生成的許多基因被下調(diào),這可能是為了防止光氧化損傷的更大風(fēng)險(xiǎn)。脂肪酸脫氫酶(fatty acid desaturase,F(xiàn)AD)是植物膜質(zhì)中不飽和脂肪酸合成的關(guān)鍵酶,負(fù)責(zé)調(diào)控脂肪酸不飽和度[48]。研究表明低溫脅迫誘導(dǎo)擬南芥(Arabidopsis thaliana)AtFAD8 基因[49]、大豆(Glycine max)GmFAD3 基因[50]、紫蘇(Perilla frutescens)FAD3 基因[48]的表達(dá),進(jìn)而提高植株體內(nèi)不飽和脂肪酸含量,保持細(xì)胞膜的穩(wěn)定性,增強(qiáng)了植物抵抗冷害的能力。隨著研究的不斷深入,人們對(duì)以CBF(C-repeat-binding factors)轉(zhuǎn)錄因子為核心的低溫信號(hào)通路有了更為清晰的認(rèn)識(shí),CBF家族成員串聯(lián)排列在擬南芥第4 條染色體上,分別命名為CBF1(DREB1B)、CBF2(DREB1C)和CBF3(DREB1A)。報(bào) 道 指 出 過(guò) 量 表 達(dá)CBF1、CBF2 和CBF3(DREB1A)均能大幅提高植株的抗凍性,并顯著誘導(dǎo)植株體內(nèi)COR(Cold-regulated)基因的表達(dá)[51]。在擬南芥野生型植株中,過(guò)表達(dá)CBF基因能夠顯著增強(qiáng)植株的抗凍性。同樣,CBF1 和CBF3 基因敲減的植株對(duì)凍敏感。然而,CBF2 突變體(cbf2)植株則表現(xiàn)抗凍的表型,基因表達(dá)分析發(fā)現(xiàn),CBF2 負(fù)反饋調(diào)控CBF1 和CBF3 的表達(dá)。將擬南芥中CBF基因過(guò)表達(dá)到其他植物中同樣能夠增強(qiáng)其抗凍能力,暗示著CBF 在植物響應(yīng)低溫過(guò)程中是非常重要而且保守的[52]。CBF的基因表達(dá)同時(shí)受轉(zhuǎn)錄激活因子和抑制子的調(diào)控。現(xiàn)在已知的正調(diào)控因子有ICE1(inducer of CBF expression 1)、ICE2 和CAMATA(calmodulin-binding transcription actieator)蛋白[52]。ICE1 蛋白能夠結(jié)合到CBF3 的啟動(dòng)子區(qū)調(diào)節(jié)CBF3 基因的表達(dá)。ice1-1 突變體對(duì)低溫敏感,而過(guò)表達(dá)ICE1 基因則導(dǎo)致植物抗凍。ICE2 是ICE1 的同源基因,能夠通過(guò)調(diào)節(jié)CBF1 的表達(dá)正向調(diào)控植物對(duì)低溫的響應(yīng)[52?53]。冷害脅迫下,植物體內(nèi)CBF3 表達(dá)增加可以使總可溶性糖含量增加,包括蔗糖、棉子糖、葡萄糖和果糖[54],進(jìn)而維持細(xì)胞的滲透平衡,緩解低溫脅迫。
2 外源ABA 緩解植物低溫脅迫的效應(yīng)
植物內(nèi)源激素調(diào)控植物的生長(zhǎng)及對(duì)逆境的應(yīng)答[55]。ABA 是20 世紀(jì)60 年代發(fā)現(xiàn)和鑒定出的一種植物內(nèi)源激素,在植物對(duì)脅迫耐受性和抗性中發(fā)揮著重要作用。外源激素通過(guò)改變內(nèi)源激素水平調(diào)節(jié)植物生理代謝,低溫脅迫下外源ABA 能夠促進(jìn)植物體內(nèi)ABA 的合成和運(yùn)輸[56],進(jìn)而調(diào)控植物的抗逆性。
2.1 低溫脅迫下外源ABA 緩解光合生理的效應(yīng)
低溫脅迫對(duì)植物最直接的影響就是光合功能下降,一般認(rèn)為低溫破壞葉片細(xì)胞中葉綠體的結(jié)構(gòu),導(dǎo)致葉綠素降解,影響光能捕獲、轉(zhuǎn)化及分配,光合電子傳遞受阻、光合機(jī)構(gòu)遭到破壞、碳同化受到抑制,最終導(dǎo)致葉片光合能力下降。噴施外源ABA 后,作物葉片的保護(hù)機(jī)能提高,過(guò)多的自由基可以被及時(shí)清除,以減少細(xì)胞膜質(zhì)過(guò)氧化產(chǎn)物的生成,并降低自由基對(duì)葉綠體的破壞,在促進(jìn)光合色素合成的同時(shí),保護(hù)光合色素結(jié)構(gòu),使葉片中葉綠素和類(lèi)胡蘿卜含量處于較高水平,最終保護(hù)光合系統(tǒng)[57]。祁利潘等[58]指出低溫條件下噴施低濃度的ABA 能夠提高PSⅡ的光化學(xué)效率(Fv/Fm),李平等[59]報(bào)道外源ABA 能有效防御低溫傷害,其可減緩葉片光合作用的下降和葉綠素?zé)晒獾拇銣纾岣咦魑锎婊盥省|S宇等[60]的研究指出噴施外源ABA 能減緩低溫下葉片Pn、Gs、Tr及Ci,提高葉片的光合能力,進(jìn)而提高植物幼苗的抗冷性。田禮欣等[61]也指出噴施適當(dāng)濃度的外源ABA 能夠顯著緩解由低溫脅迫導(dǎo)致的作物葉片Pn、Gs及Tr的下降幅度,提高Fv/Fm和Fv/Fo,降低Fo水平,進(jìn)而提高幼苗葉片的光合能力,提高PSⅡ反應(yīng)中心活性,促進(jìn)物質(zhì)積累,提高玉米幼苗的耐冷性。
2.2 低溫脅迫下外源ABA 調(diào)控抗逆生理的效應(yīng)
外源ABA 具有調(diào)控植物的抗逆生理的功能,在抵御低溫脅迫方面具有多條路徑,其核心機(jī)制是減少活性氧類(lèi)物質(zhì)的積累,降低膜質(zhì)過(guò)氧化程度,減少其對(duì)細(xì)胞膜的損失,保障細(xì)胞膜的正常滲透調(diào)節(jié)功能以維持正常的生理代謝。主要應(yīng)激反應(yīng)是增強(qiáng)抗氧化酶活性,增加非酶抗氧化劑含量,提高滲透調(diào)節(jié)物質(zhì)的含量,以調(diào)節(jié)MDA含量降低、減少H2O2等ROS 物質(zhì)的積累。項(xiàng)洪濤等[34]報(bào)道指出外源ABA 能夠提高低溫脅迫時(shí)植物體內(nèi)SOD、POD 以及CAT 的活性,并可同時(shí)提高脯氨酸、可溶性糖等可溶類(lèi)物質(zhì)的含量,抵御低溫傷害。李平等[59]指出低溫條件下,外源ABA 能夠降低葉片內(nèi)MDA 的積累,緩解脅迫。方彥等[62]指出外源ABA 能提高作物氧化酶活性,同時(shí)減緩膜質(zhì)過(guò)氧化物MDA 的積累,具有提高作物抗寒性的誘導(dǎo)效應(yīng),Xiang 等[63]表明低溫條件下外源ABA能提高水稻葉片內(nèi)脯氨酸、可溶性糖和可溶性蛋白含量,有效提高SOD、POD 和CAT 活性,增強(qiáng)水稻的抗冷性。蒲高斌等[64]指出低溫脅迫下外源ABA 能有效提高西瓜(Citrullus lanatus)幼苗葉片中SOD 和CAT 活性、降低MDA 的積累,維持膜的完整性,同時(shí)促進(jìn)滲透保護(hù)物質(zhì)脯氨酸和可溶性糖的增加,增強(qiáng)植株的抗寒性,同時(shí)也表明這是外源ABA 有效提高西瓜幼苗抗冷性、減輕低溫對(duì)西瓜苗傷害的主要生理基礎(chǔ)。
2.3 低溫脅迫下外源ABA 調(diào)控細(xì)胞膜組分及結(jié)構(gòu)的效應(yīng)
抗冷能力較高的植物之所以有這樣的特性,從膜脂結(jié)構(gòu)的角度來(lái)看,一是因?yàn)樵陂L(zhǎng)期對(duì)低溫的適應(yīng)過(guò)程和自然選擇過(guò)程中,其本身形成了具有較多不飽和脂肪酸的狀態(tài),這由遺傳基因決定,二是因?yàn)樵趹?yīng)激低溫的理化反應(yīng)中,能在細(xì)胞膜中優(yōu)先積累較多的不飽和脂肪酸并使之迅速參與膜的結(jié)構(gòu)以適應(yīng)抵御低溫環(huán)境的生理需求[65]。膜質(zhì)中脂肪酸不飽和度對(duì)于植物的低溫適應(yīng)性尤為關(guān)鍵,調(diào)整脂肪酸的不飽和度是植物適應(yīng)低溫脅迫的重要方式[66],較多的不飽和脂肪酸能維持細(xì)胞的流動(dòng)性,為跨膜蛋白質(zhì)和酶功能發(fā)揮提供適宜的環(huán)境[67]。王軍虹等[65]指出低溫條件下,外源ABA 能夠使小麥體內(nèi)的不飽和脂肪酸明顯提高,并表明其原因是ABA 的噴施有利于亞麻酸(18:3)的積累,進(jìn)而提高抗寒力。付翀等[68]也表明外源ABA 能相對(duì)提高亞麻酸(18:3)的含量,進(jìn)而提高作物的抗冷能力。徐琛[69]的研究也證實(shí)了外源ABA 能提高低溫條件下作物體內(nèi)不飽和脂肪酸含量,增強(qiáng)作物的抗寒性。
2.4 低溫脅迫下外源ABA 調(diào)控內(nèi)源激素水平的效應(yīng)
內(nèi)源激素在植物生長(zhǎng)和生存的各個(gè)生物學(xué)進(jìn)程均發(fā)揮著關(guān)鍵作用,目前已經(jīng)發(fā)現(xiàn)乙烯(ethylene,ET)、油菜素內(nèi)酯(brassinolide,BR)和茉莉酸(JA)可通過(guò)CBF 信號(hào)通路影響植物的低溫耐受性[2]。前人研究指出各類(lèi)激素間有相互拮抗和相互促進(jìn)的關(guān)系,各種激素之間的動(dòng)態(tài)平衡是調(diào)節(jié)植物生長(zhǎng)和生存的關(guān)鍵所在[34],段娜等[70]指出內(nèi)源激素水平及激素間的平衡變化對(duì)植物應(yīng)激非生物脅迫起到重要作用。通常認(rèn)為內(nèi)源ABA 與植物抗逆關(guān)系最為密切,有報(bào)道指出ABA 是應(yīng)激低溫的重要信號(hào)因子,對(duì)細(xì)胞的微管結(jié)構(gòu)具有保護(hù)功能,低溫脅迫下植物大量快速積累ABA,發(fā)揮保護(hù)功能[43]。黃杏等[71]指出低溫條件下施用外源ABA 改變了作物體內(nèi)激素水平,ABA含量增加、GA 含量降低。李馨園等[43]也表明低溫脅迫能促進(jìn)作物內(nèi)源ABA 水平提高,外施ABA 可以更進(jìn)一步加強(qiáng)內(nèi)源ABA 的合成,加強(qiáng)各種生理生化保護(hù)機(jī)制,提高植物的抗冷性。Xiang 等[63]指出低溫條件下,葉面噴施外源ABA 促進(jìn)葉片內(nèi)源ABA 含量增加,同時(shí)降低了IAA 和GA 的含量,并指出外源ABA 提高了作物抵御低溫的機(jī)能。
3 結(jié)論與展望
我國(guó)地大物博,幅員遼闊,南北跨度大,各地區(qū)生態(tài)環(huán)境迥異,氣候條件差別大,這對(duì)農(nóng)業(yè)生產(chǎn)要求更為嚴(yán)格和復(fù)雜。雖然全球氣候變暖,但我國(guó)北方夏季突發(fā)低溫情況反而有加劇的趨勢(shì),南方的倒春寒天氣也時(shí)有發(fā)生,這對(duì)農(nóng)業(yè)生產(chǎn)極為不利,如何抵御突發(fā)的氣象災(zāi)害來(lái)穩(wěn)定農(nóng)業(yè)生產(chǎn)是一個(gè)重大的并且持續(xù)的科學(xué)問(wèn)題。
低溫對(duì)作物產(chǎn)量影響巨大,嚴(yán)重時(shí)甚至可以導(dǎo)致絕產(chǎn)。目前,研究學(xué)者已經(jīng)逐步掌握了植物抵御低溫的生理機(jī)制,在此基礎(chǔ)上順延摸索緩解或抵御低溫的措施,外施植物生長(zhǎng)物質(zhì)來(lái)提高植物的抗逆能力已經(jīng)達(dá)成廣泛共識(shí),通過(guò)化控方法抵御逆境脅迫雖然取得了一些成效,但還是較為片面,而且更大程度上是緩解,而沒(méi)有達(dá)到安全度過(guò)突發(fā)低溫的短暫時(shí)期,這還需要廣大農(nóng)業(yè)科研工作者繼續(xù)挖掘新方法,探索新途徑。一是通過(guò)現(xiàn)有掌握的植物抗逆生理機(jī)制,合成新的、效果更好的調(diào)節(jié)劑,或者開(kāi)發(fā)新的調(diào)節(jié)劑復(fù)配產(chǎn)品,使得應(yīng)用效果更好;二是根據(jù)試驗(yàn)和生產(chǎn)實(shí)踐發(fā)現(xiàn)更多的抗低溫品種,通過(guò)育種手段,逐代篩選,育成高抗且高產(chǎn)優(yōu)質(zhì)的植物新品種,同時(shí)也可以通過(guò)全基因組關(guān)聯(lián)分析快速鑒定與深入挖掘耐低溫的相關(guān)基因,結(jié)合多組學(xué)聯(lián)合分析等技術(shù)更加方便地解析調(diào)控機(jī)理,從分子水平提高植物耐寒性,最終應(yīng)用于育種改良達(dá)到提高作物耐寒性的目的,實(shí)現(xiàn)災(zāi)年穩(wěn)產(chǎn)、平年豐產(chǎn)的生產(chǎn)目標(biāo)。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級(jí)版(2021年3期)2021-03-19 06:12:40
小天使·二年級(jí)語(yǔ)數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫(huà)刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
