解淀粉芽孢桿菌化感作用對球等鞭金藻生長抑制效果
2021-02-03 13:26:16李曉英陳義華趙裕超馮森磊李雯倩董志國
海洋科學 2021年1期
李曉英, 陳義華, 趙裕超, 馮森磊, 李雯倩, 朱 強, 董志國
解淀粉芽孢桿菌化感作用對球等鞭金藻生長抑制效果
李曉英1, 陳義華2, 趙裕超1, 馮森磊1, 李雯倩1, 朱 強1, 董志國1
(1. 江蘇海洋大學 江蘇省海洋生物技術重點實驗室, 連云港 222005; 2. 上海海洋大學 水產動物遺傳育種中心上海市協同創新中心, 上海 200090)
為探明解淀粉芽孢桿菌及其代謝物對海水養殖環境中微藻的化感作用, 本文以實驗生態學方法研究了解淀粉芽孢桿菌對球等鞭金藻生長的生態作用。實驗設計了不同劑量(100mL、500mL、1 000mL)的解淀粉芽孢桿菌純菌液、解淀粉芽孢桿菌代謝產物、解淀粉芽孢桿菌及代謝產物混合液三種組合研究其對球等鞭金藻的培養影響。結果顯示, 添加純菌液組金藻在第1~2 d增長緩慢, 藻細胞數無顯著差異, 但均低于對照組, 其中第3~4 d顯著低于對照組(<0.05); 添加菌代謝產物組第1 d金藻細胞數與對照組相比無顯著差異, 2~4 d顯著低于對照組(<0.05); 添加菌液及代謝產物混合液組中100mL組金藻生長較緩慢, 500 μL和1 000 μL組藻細胞數呈下降趨勢, 藻細胞數顯著低于對照組(<0.05), 1 000 μL組金藻第2 d已全部死亡。各劑量組藻細胞數均呈現解淀粉芽孢桿純菌液、解淀粉芽孢桿菌代謝產物、解淀粉芽孢桿菌及代謝產物混合液依次下降, 另外, 除菌代謝物1 000 μL劑量組藻細胞數大于500 μL劑量組外, 其余各組添加量越大藻細胞數越少, 呈現劑量效應關系。解淀粉芽孢桿純菌液、解淀粉芽孢桿菌代謝產物、解淀粉芽孢桿菌及代謝產物混合液抑制率最高分別為35.84%、69.75%、100%, 表明解淀粉芽孢桿菌對球等鞭金藻的作用為化感抑制, 且抑制效果具有劑量效應。本研究為解淀粉芽孢桿菌在水產養殖中應用效果評價提供了科學依據, 同時為赤潮藻類治理提供新思路。
球等鞭金藻; 解淀粉芽孢桿菌; 化感作用; 生長性能
微藻作為優質餌料以及改善水質的首選生物一直在水產養殖中占據重要的地位。球等鞭金藻()無細胞壁, 粒徑小, 營養豐富易吸收, 是多種海產動物喜食的餌料生物[1]。益生菌的使用在水產養殖中已經非常普遍, 其安全問題一直是人們關注的熱點[2]。另外競爭抑制是益生菌的重要作用機制[3], 這也給水中微藻生長和繁殖造成很大的影響[4], 因此研究微生物對微藻的作用效果意義重大。
化感作用多指植物通過釋放化學物質到環境中從而產生對其他植物直接或間接、有利或不利作用[5], 但化感作用同樣普遍存在于微生物之間[6-7]。解淀粉芽孢桿菌()屬芽孢桿菌屬, 是廣泛存在于自然界的一種非致病性細菌, 能夠分泌抗生素、抗菌蛋白或多肽類物質, 對多種病原菌、真菌等具有良好的抑制作用[8], 如對煙草的促生長及抗煙草花葉病毒的作用[9], 抑制煙草青枯病菌()[10]、黃瓜花葉病毒的侵染[11]。目前, 大型水生植物以及微藻間的化感作用研究較多[12-15], 微生物對與藻類之間的相互作用也有研究。韓光耀等發現溶藻細菌DH-e代謝產物可以抑制東海原甲藻()的生長[16], Zheng等在河口土壤和海水中發現了多種殺藻細菌[17], Wang等在赤潮水體中分離出了一種可以使塔瑪亞歷山大藻()溶解的細菌分泌蛋白[18]。解淀粉芽孢桿菌已被作為水產養殖中常用水質改良劑或復合微生態制劑的主要有益菌之一被廣泛開發應用[19], 但其對海洋微藻的作用究竟是化感促進還是化感抑制還有待探討。因此, 本文首次以海水養殖池塘中常見的微藻餌料球等鞭金藻為試驗材料, 探明解淀粉芽孢桿菌及其代謝物對這種海藻的化感作用, 從而為解淀粉芽孢桿菌在水產養殖中應用效果評價提供科學依據, 同時為赤潮藻類治理提供新思路。
1 材料與方法
1.1 解淀粉芽孢桿菌及其代謝物的制備
試驗所用解淀粉芽孢桿菌B5從對蝦養殖海水中直接分離、篩選、鑒定后通過培養而獲得。將經培養后的解淀粉芽孢桿菌接種到種子培養基中制備種子液。種子培養基: 葡萄糖2.5 g, 氯化鈉25 g, 胰蛋白胨17 g, 磷酸氫二鉀2.5 g, 蒸餾水1000 mL, pH自然, 121℃, 滅菌25 min。將制備的種子液接種到發酵培養基中制備發酵液。發酵培養基: 玉米粉2.9 g, 豆餅粉12.5 g, 磷酸氫二鉀1.5 g, 氯化鈉27.6 g、蒸餾水1000 mL, pH自然, 121℃, 滅菌25 min。發酵采用16 L發酵罐。發酵條件: 裝液系數0.75, 初始pH 7.5, 接種量3%, 發酵溫度31℃, 機械攪拌轉速180 r/min, 通氣量1.2 V/(V·min), 發酵周期36 h。將一部分發酵產物用0.22mm的微孔過濾膜過濾, 獲得不含解淀粉芽孢桿菌的發酵液(解淀粉芽孢桿菌代謝產物); 另一部分不經過濾, 為解淀粉芽孢桿菌液和發酵液混合液, 備用。
1.2 藻種及培養條件
球等鞭金藻藻種由上海海洋大學藻種室提供。活化后在光照培養箱中培養, 參數設定: 溫度20℃, 光強60 μmol photons/(m2·s), 光周期12 L︰12 D。培養用海水為經0.45mm濾膜抽濾、滅菌處理的自然海水, 營養液為f/2培養液。每天定時搖動培養瓶兩次, 防止其附壁生長, 期間維持藻液pH在7.9~8.1。
1.3 球等鞭金藻細胞數測定
取1 mL藻液, 用血球計數板在光學顯微鏡下計數, 計算出此藻液濃度。稀釋藻液, 使其濃度分別為原來的20%、40%、60%、80%、100%, 在663 nm波長處分別測量吸光值, 繪制藻濃度與吸光度的線性標準曲線, 用于后續實驗球等鞭金藻細胞數的計算。球等鞭金藻細胞密度與吸光值之間的線性回歸方程及相關系數為:=104.97+0.153 9,=0.992 7, 線性曲線見圖1。
1.4 菌藻作用試驗設計
實驗用三角瓶規格為250 mL, 均經反復沖洗及高壓滅菌處理。將處于對數生長期的球等鞭金藻50 mL接種到三角瓶中, 瓶中海水體積為200 mL, 內含f/2培養液, 初始密度為1.83×106個/mL。實驗進行4 d, 菌藻共培條件同1.2。
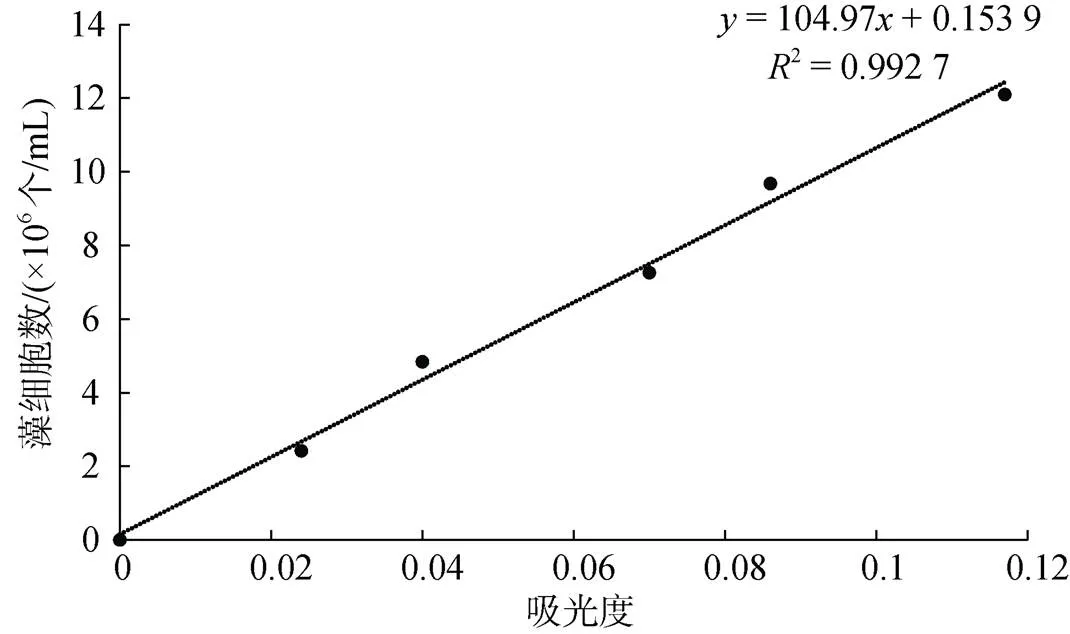
圖1 球等鞭金藻細胞數與吸光值之間的線性曲線
將實驗分為對照組、實驗組和空白組, 每組均設置三個重復。對照組只接種50 mL球等鞭金藻(I組)。實驗組分為三組, 每組先接種與對照組等量的球等鞭金藻, 第一組再接種不含發酵液的解淀粉芽孢桿菌純菌液(純菌液組, 密度為1.0×109個/mL), 第二組再接種解淀粉芽孢桿菌代謝產物(代謝物組), 第三組再接種解淀粉芽孢桿菌液和代謝產物的混合液(混合液組, 其中含解淀粉芽孢桿菌密度為1.0×109個/mL),劑量均分別為100 μL、500 μL、1000 μL, 相關表示方法見表1。空白組也分為三組, 不接藻種, 分別對應三個對照組添加相同劑量的三種解淀粉芽孢桿菌添加液。每天定時測量每組的吸光值, 計算球等鞭金藻細胞濃度, 其中實驗組藻細胞濃度計算時, 最終吸光值為對照組吸光值減去對應空白組的吸光值。
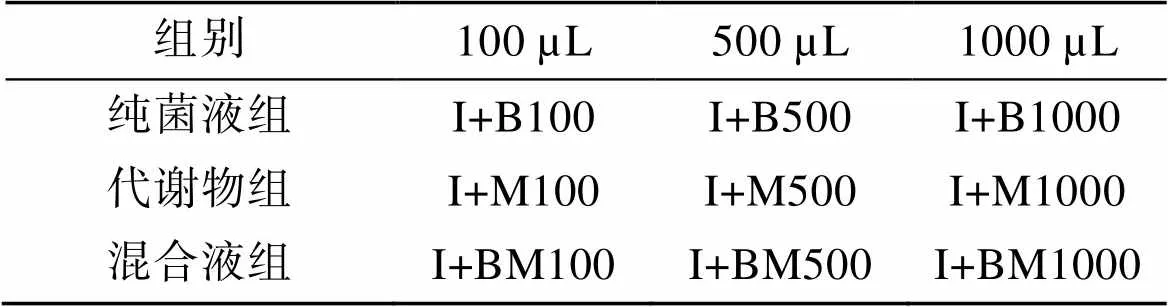
表1 解淀粉芽孢桿菌與球等鞭金藻共培養表示方法
注: I: 球等鞭金藻; B: 解淀粉芽孢桿菌; M: 菌代謝產物
1.5 數據處理
抑制率公式: PI(%)=(1–N/N0)×100%。其中, N0和N分別為對照組球等鞭金藻濃度和試驗組球等鞭金藻濃度; PI為抑制比例, PI=50%時對應的濃度為EC50。
數據用Statistica 5.5軟件做ANOVA分析, 并進行Duncan多重比較, 以<0.05作為差異顯著水平。
2 結果與討論
2.1 解淀粉芽孢桿菌對球等鞭金藻生長的影響
與對照組相比, I+B100、I+B500、I+B1000三組球等鞭金藻在第1~3 d增長緩慢, 藻細胞數無顯著差異, 但均低于對照組, 其中第3~4 d三組均顯著低于對照組(<0.05, 圖2a)。各組抑制率均沒有超過50%(圖2b)。前3 d, I+B1000組抑制率持續增加, I+B100、I+B500兩組抑制率在第1 d時增加明顯且高于I+B1000組, 在第2 d時下降。第3 d后I+B100和I+B500兩組抑制率開始下降, 其藻細胞數也開始明顯增加, 但仍顯著低于對照組(<0.05), I+B1000組抑制率高于另外兩組, 藻細胞數最低。實驗結束, I+B100、I+B500、I+B1000三組中的球等鞭金藻藻細胞數分別比對照組低了7.95%、23.77%、35.80%。
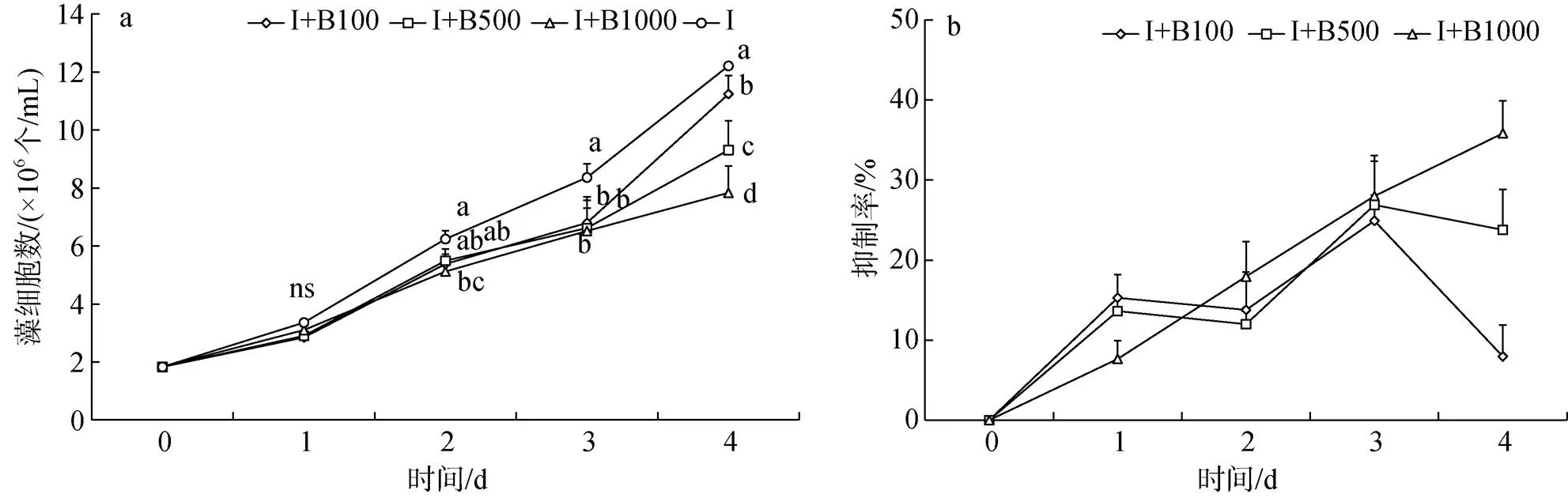
圖2 添加解淀粉芽孢桿菌純菌液組球等鞭金藻生長曲線和抑制率
注: 圖2a中相同時間不同添加量間字母不同表示藻細胞數存在顯著差異(<0.05), ns表示不存在顯著差異。
2.2 解淀粉芽孢桿菌代謝產物對球等鞭金藻生長的影響
第1 d, I+M100、I+M500、I+M1000三組球等鞭金藻細胞數與對照組相比無顯著差異(>0.05, 圖3a), 第2 d和第3 d三組均顯著低于對照組(<0.05), 其中I+M500和I+M1000兩組差異不顯著(>0.05), 第4 d四組均存在顯著差異(<0.05), 添加代謝物量越多, 藻細胞數越低。抑制率在第1 d后明顯增加(圖3b), 第3 d后抑制率增加減緩, I+M1000組抑制率下降。實驗結束, I+M100、I+M500、I+M1000三組中的球等鞭金藻藻細胞數分別比對照組下降了45.16%、69.67%、53.52%。
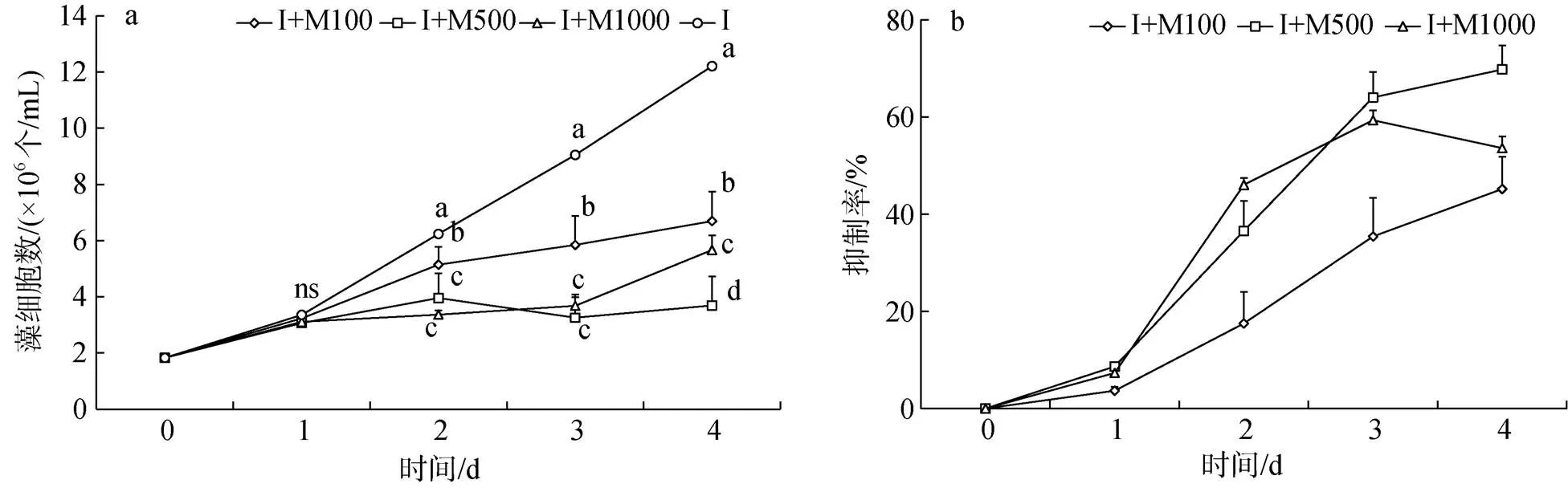
圖3 添加解淀粉芽孢桿菌代謝產物組球等鞭金藻生長曲線和抑制率
注: 圖3a中相同時間不同添加量間字母不同表示藻細胞數存在顯著差異(<0.05), ns表示不存在顯著差異。
2.3 解淀粉芽孢桿菌及代謝產物混合液對球等鞭金藻生長的影響
與對照組相比, I+BM100組球等鞭金藻生長較緩慢, I+BM500、I+BM1000兩組藻細胞數呈下降趨勢(圖4a), 藻細胞數顯著低于對照組(<0.05), 其中I+BM1000組球等鞭金藻第2 d已經全部死亡。實驗開始后三組抑制率增加明顯(圖4b), 1 d后緩慢增加, I+BM1000組抑制率第2 d已經為100%。實驗結束, I+M100、I+M500、I+M1000三組中的球等鞭金藻藻細胞數分別比對照組下降了57.54%、94.19%、100%。
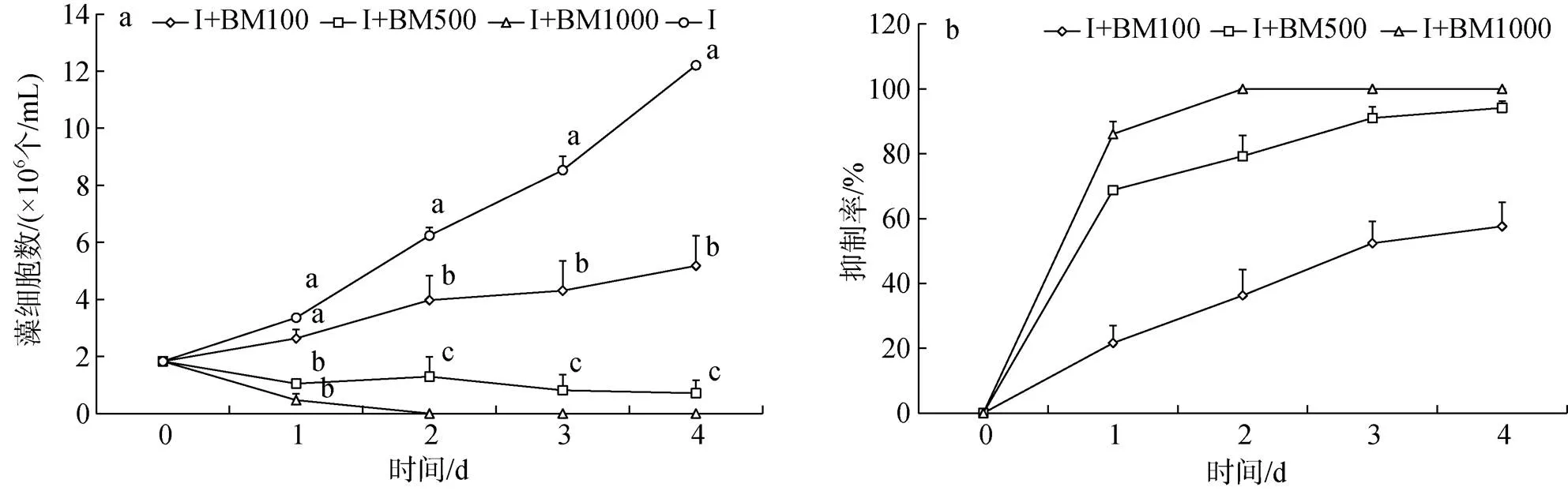
圖4 添加混合液組球等鞭金藻生長曲線和抑制率
注: 圖4a中相同時間不同添加量間字母不同表示藻細胞數存在顯著差異(<0.05)
2.4 相同條件下不同添加物對球等鞭金藻生長的影響
為便于分析三種不同解淀粉芽孢桿菌添加物對球等鞭金藻影響的差異, 圖5為實驗結束后球等鞭金藻細胞數和抑制率。添加解淀粉芽孢桿菌純菌液、代謝物以及混合物球等鞭金藻細胞數均低于對照組。添加不同濃度藻細胞數也有差異(圖5a), 各濃度組藻細胞數均呈現解淀粉芽孢桿菌純菌液、代謝物以及混合物依次下降, 另外除I+M1000組藻細胞數大于I+M500外, 其余各組添加劑量越大藻細胞數越少。抑制率則與藻細胞數相反(圖5b), 各濃度組抑制率均呈現解淀粉芽孢桿純菌液、代謝物以及混合物依次上升, 除I+M1000組抑制率小于I+M500外, 其余各組添加劑量越大抑制率越高。
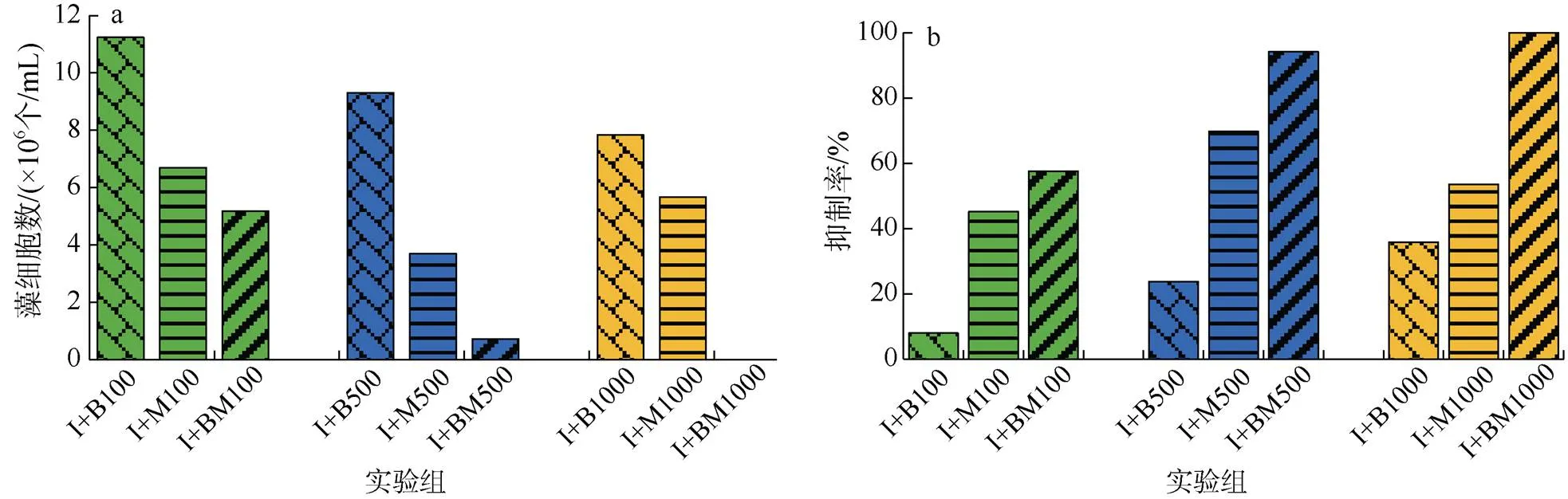
圖5 添加不同物質組球等鞭金藻細胞數和抑制率對比
3 討論
益生菌因具有提高水產動物免疫力, 改善水質等優點在水產養殖中被大量使用, 但是部分益生菌可以對水產動物甚至人體造成損害, 例如芽孢桿菌屬[20-21]。解淀粉芽孢桿菌屬于芽孢桿菌屬, 但是已有研究表明該種菌的微膠囊對水產動物沒有毒性[19]。微藻是水產貝類以及其他經濟動物常用的生物餌料, 解淀粉芽孢桿菌對有益微藻的作用效果研究相對較少。球等鞭金藻是水產中常用的微藻餌料, 本實驗目的就是探明解淀粉芽孢桿菌及其代謝產物能否對球等鞭金藻生長造成影響, 進而探究這種影響是正面的還是負面的。
化感作用多被用于植物之間的相關研究, 微生物與微藻之間的化感作用對水產養殖生產具有很好的指導作用。抑藻細菌的作用方式可分為直接接觸抑制和釋放抑制物質的間接抑制[22-23], 解淀粉芽孢桿菌在生長繁殖過程中會產生一些代謝產物, 這些代謝產中含有抑藻物質, 從而起到間接的抑藻作用。許波在對解淀粉芽孢桿菌的研究中發現其代謝產物中含有的賴氨酸和苯丙氨酸, 二者可共同對微藻水華起到抑制作用[24]。本研究發現, 解淀粉芽孢桿菌對球等鞭金藻同樣具有抑制效果, 添加純菌液組、代謝物組和混合液組均表現出了較強的抑制作用, 球等鞭金藻細胞數顯著低于未添加組。其中純菌液組抑制率最低, 抑制率最高35.84%, 100 μL劑量組3 d后抑制率下降, 球等鞭金藻還可以繼續恢復增長, 可以用于減緩球等鞭金藻老化; 混合液組抑制效果最為顯著, 抑制率最高可達到100%, 100 μL劑量組球等鞭金藻生長極為緩慢, 可用于球等鞭金藻的長期保存。Li等[25]發現菌藻之間是通過化學信號交流的, 而在植物化感作用研究中, 多為植物的提取液或者研磨液[26-27], 可以推測這種化學物質是存在于植物體內, 需要對其處理后才能大量釋放, 從而對其他物種起到化感抑制作用, 而本身具有的抑制作用較小, 這可能是添加純菌液組抑制效果最差的原因。抑藻細菌可通過產生抗生素、肽類化合物、氨基酸及小分子化合物等對微藻起到抑制作用[23], 但是本文結果顯示添加混合液組的抑制作用遠強于其他兩組并能產生致死效果, 這也說明可能存在接觸抑制或者直接溶藻的方式[23]。因此解淀粉芽孢桿菌具體的作用機理將是本研究后續的主要方目標。
微生物對微藻的化感作用除了抑制還有促進作用, 有些細菌可以產生植物細胞分類素和茁壯素等植物激素起到促進生長的作用[28], Dimitrieva等也發現檸檬假交替單胞菌()可以促進海帶()的生長[29], 本研究對解淀粉芽孢桿菌的三種添加物分別設置了100、500 μL、1000 μL三種濃度, 發現對球等鞭金藻均是抑制作用, 而且后續又做了10 μL、30 μL的添加實驗, 同樣沒有發現促進效果, 說明解淀粉芽孢桿菌對球等鞭金藻的生長確實存在抑制效應。沙俊濤等研究發現化感作用存在濃度效應[30], 且呈現低促高抑的規律。本研究中三種添加解淀粉芽孢桿菌添加液明顯對球等鞭金藻產生了抑制, 并隨濃度的升高效果更明顯, 表明解淀粉芽孢桿菌化感作用存在濃度效應。添加純菌液組和混合液組抑制率均隨解淀粉芽孢桿菌添加劑量的增加而增加, 但是添加代謝物組在3 d后500 μL劑量組的抑制率超過了1000 μL劑量組, 說明解淀粉芽孢桿菌代謝物抑制作用并不是濃度越高越好, 這其中原因還需要進一步的實驗研究。當抑制物濃度達到一定閾值才出現明顯的化感抑制作用, 在閾值之上濃度越高抑制作用越顯著[31]。在同等的濃度下, 添加代謝產物組不能致死, 而添加混合液組所顯示出的抑制作用遠強于其他兩組并能產生致死效果, 其抑制效果大于其他兩組的疊加。可能是由于解淀粉芽孢桿菌的直接溶藻作用增加了混合液對球等鞭金藻的抑制效果, 這也能解釋為什么代謝物1000 μL劑量組抑制率低, 而組混合液1000 μL劑量組抑制率提高的原因。赤潮藻治理問題成為近幾年的研究熱點, 本研究發現添加混合液組對球等鞭金藻有致死效果, 并且正在進行的解淀粉芽孢桿菌對赤潮藻——三角褐指藻()的影響實驗中也發現了很好的抑制效果, 這也為赤潮藻的治理提供了新思路。
4 結論
解淀粉芽孢桿菌及其代謝產物可抑制球等鞭金藻細胞的生長, 其中代謝產物的抑制作用優于解淀粉芽孢桿純菌液, 而兩種混合后的抑制作用最顯著, 具有致死效果。抑制效果具有劑量效應, 基本呈現抑制效果與添加劑量成正相關。另外, 解淀粉芽孢桿菌純菌液100 μL球等鞭金藻還可以繼續恢復增長, 可用于減緩球等鞭金藻老化; 混合液100 μL劑量組球等鞭金藻生長極為緩慢, 可用于球等鞭金藻的長期保存。
[1] 孔周雁, 張羽帆, 凌婷, 等. 球等鞭金藻三級培養過程中細菌群落多樣性分析[J]. 核農學報, 2020, 34(3): 506-514. Kong Zhouyan, Zhang Yufan, Ling Ting, et al. Analysis of bacterial community diversity in tertiary culture of[J]. Journal of Nuclear Agricultural Sciences, 2020, 34(3): 506-514.
[2] 玄振盈, 于洋. 水產動物飼料中益生菌作用機制的研究進展[J]. 山東畜牧獸醫, 2017, 38(7): 79.Xuan Zhenying, Yu Yang. Research progress on the mechanism of probiotics in aquatic animal feeds[J]. Shandong Vocational Animal Science and Veterinary, 2017, 38(7): 79.
[3] 李海兵, 宋曉玲, 李赟, 等. 水產動物益生菌研究進展[J]. 動物醫學進展, 2008, 29(5): 94-99. Li Haibing, Song Xiaoling, Li Yun, et al. Progress on probiotic bacteria in aquiculture[J]. Progress in Veterinary Medicine, 2008, 29(5): 94-99.
[4] 劉君, 宋曉玲, 陳志鑫. 益生菌對水產動物的作用研究進展[J]. 動物醫學進展, 2009, 30(9): 78-81. Liu Jun, Song Xiaoling, Chen Zhixin. Progress on the function of probiotics in aquaculture[J]. Progress in Veterinary Medicine, 2009, 30(9): 78-81.
[5] 袁美麗, 李韶霞, 王寧. 5種外來入侵植物對小麥的化感作用[J]. 貴州農業科學, 2016, 44(12): 58-62.Yuan Meili, Li Shaoxia, Wang Ning. Allelopathic effects of five invasive plants species on wheat[J]. Guizhou Agricultural Sciences, 2016, 44(12): 58-62.
[6] 宋文娟, 張昊. 植物他感作用的研究進展[J]. 內蒙古林業科技, 2013, 39(1): 54-57. Song Wenjuan, Zhang Hao. Research progress on allopathy of plants[J]. Journal of Inner Mongolia Forestry Science and Technology, 2013, 39(1): 54-57.
[7] Rice E L. Allelopathy (2nd ed) [M]. New York: Academic Press Inc, 1984: 1-5, 309-315.
[8] 王鵬, 郭麗, 孔璐, 等. 堿浸提和纖維素酶輔助提取藍莓多糖及其抗氧化能力的對比研究[J]. 糧食與油脂, 2017, 30(4): 94-97. Wang Peng, Guo Li, Kong Lu, et al. Comparison of antioxidant capacity of blueberry polysaccharide by alkali extraction and cellulose assisted treatment[J]. Cereals & Oils, 2017, 30(4): 94-97.
[9] 申莉莉, 王鳳龍, 錢玉梅, 等. 解淀粉芽孢桿菌Ba33對煙草的促生及抗TMV作用[J]. 吉林農業大學學報, 2010, 32(4): 383-386. Shen Lili, Wang Fenglong, Qian Yumei, et al. Tobacco growth-promotion effect and TMV resistance ofBa33[J]. Journal of Jilin Agricultural University, 2010, 32(4): 383-386.
[10] Soad A Algam, Xie Guan lin, Li Bin. Comparat ive performance ofspp. in growth promotion and suppression of tomato bacterial wilt caused by[J]. Journal of Zhejiang University (Agriculture & Life Science), 2004, 30(6): 603-610.
[11] Murphy J F, Reddy M S, Ryu C M, et al. Rhizobacteria-Mediated growth promotion of tomato leads to protection against[J]. Phytopathology, 2003, 93: 1301-1307.
[12] Idriss E E, Makarew icz O, Farouk A, et al. Extracellular phytase activity ofFZB45 contributes to its plant growth promoting effect[J]. Microbiology, 2002, 148: 2097-2109.
[13] Felpeto A B, ?liwińska-Wilczewska, Sylwia, Klin M, et al. Temperature-dependent impacts of allopathy on growth, pigment, and lipid content between a subpolar strain ofsp. CCBA MA-01 and coexisting microalgae[J]. Hydrobiologia, 2019, 835: 117-128.
[14] 巨穎琳, 李小明. 南四湖3種沉水植物對銅綠微囊藻化感作用研究[J]. 山東大學學報(理學版), 2011, 46(3): 1-8. Ju Yinglin, Li Xiaoming. Allelopathic effects of three submerged macrophytes in the Nansi Lake on[J]. Journal of Shandong University (Natural Science), 2011, 46(3): 1-8.
[15] 張議文, 王江濤, 譚麗菊. 旋鏈角毛藻對中肋骨條藻化感作用的影響因素及化感物質性質初探[J]. 海洋學報, 2014, 36(2): 123-129. Zhang Yiwen, Wang Jangtao, Tan Liju. The study on the influencing factors ofallelopa-thic effect onthe primary research on the properties ofalle-lochemical[J]. Acta Oceanologica Sinica, 2014, 36(2): 123-129.
[16] 韓光耀, 謝麗玲, 畢瀟, 等. 溶藻細菌DH-e代謝產物對東海原甲藻的抑制作用[J]. 應用海洋學學報, 2017, 36(2): 151-157.Han Guangyao, Xie Liling, Bi Xiao, et al. Inhibitory effect of metabolites from algicidal bacterium DH-e on[J]. Journal of Applied Oceanography, 2017, 36(2): 151-157.
[17] Zheng N N, Ding N, Gao P K, et al. Diverse algicidal bacteria associated with harmful bloom-formingin estuarine soil and seawater[J]. Science of the Total Environment, 2018, 631-632: 1415-1420.
[18] Wang B, Yang X R, Lu J, et al. A marine bacterium producing protein with algicidal activity against[J]. Harmful Algae, 2012, 13(1): 69-88.
[19] 曹海鵬, 衛若鵬, 何珊, 等. 水產養殖用解淀粉芽孢桿菌微膠囊的安全性評價[J]. 中國生物工程雜志, 2012, 32(5): 58-65. Cao Haipeng, Wei Ruopeng, He Shan, et al. Safety evaluation ofmicrocapsules used for aquaculture[J]. China Biotechnology, 2012, 32(5): 58-65.
[20] Cho H, Liu L, Liu K, et al. Phenotypic characterization and phylogenetic analysis of a virulentstrain from the Tiger frog,Wiegmann[J]. African Journal of Microbiology Research, 2010, 4(24): 2780-2786.
[21] 陳玉梅, 程茜. 益生菌的安全性[J]. 中國微生態學雜志, 2010, 22(9): 851-853. Chen Yumei, Cheng Xi. Safety of probiotics[J]. Chinese Journal of Microecology, 2010, 22(9): 851-853.
[22] Li Y, Lei XQ, Zhu H, et al. Chitinase producing bacteria with direct algicidal activity on marine diatoms[J]. Scientific Reports, 2016, 6(1): 21984.
[23] 焦彥凱, 嚴小軍, 李小兵. 溶藻細菌及溶藻化合物研究進展[J]. 工業微生物, 2018, 48(4): 56-62. Jiao Yankai, Yan Xiaojun, Li Xiaobing. Advances in research of algicidal bacteria and algae-lysing compounds[J]. Industrial Microbiology, 2018, 48(4): 56-62.
[24] 許波. 一株芽孢桿菌溶藻活性物質的分離及其結構分析[D]. 湖北武漢: 武漢工業學院, 2012.Xu Bo. The separation and structure analysis of algici-dal active substances in a strain of Bacillus[D]. Wuhan, Hubei Province: Wuhan Polytechnic University, 2012.
[25] Li J, Fergola P, Ma Z. Effects of allelochemicals produced by one population in a chemostat-like environment[J]. Journal of Theoretical Biology, 2011, 284(1), 99-105.
[26] 劉序, 張如義, 楊小建, 等. 核桃葉水浸液對蘿卜種子萌發的化感作用[J]. 江蘇農業科學, 2020, 48(3): 146-151. Liu Xu, Zhang Ruyi, Yang Xiaojian, et al. Allelopathic effect of water extracts from walnut leaves on seed germination of radish[J]. Jiangsu Agricultural Sciences, 2020, 48(3): 146-151.
[27] 常孟陽, 董靜, 李晨露, 等. 菹草對小球藻和卵囊藻的形態化感效應研究[J]. 水生態學雜志, 2020, 41(2): 91-97. Chang Mengyang, Dong Jing, Li Chenlu, et al. Allelopathic effect of the macrophyteon colony formation ofandsp[J]. Journal of Hydroecology, 2020, 41(2): 91-97.
[28] Maruyama A, Maeda M, Simidu U. Distribution and classification of marine bacteria with the ability of cytokinin and auxin production[J]. Bulletin of Japanese Society of Microbial Ecology, 1990, 5(1): 1-8.
[29] Dimitrieva G Y, Crawford R L, Yüksel G U. The nature of plant growth-promoting effects of a pseudoalteromonad associated with the marine algaeand linked to catalase excretion[J]. Journal of Applied Microbiology, 2006, 100(5): 1159-1169.
[30] 沙俊濤, 陳青青, 繆雨靜, 等. 茶樹凋落葉浸提液對菘藍生理生化的化感效應[J]. 核農學報, 2020, 34(6): 1327-1336. Sha Juntao, Chen Qingqing, Miu Yujing, et al. Allelopathic effect of leaf litter extracts ofon the physiology and biochemistry ofFort[J]. Journal of Nuclear Agricultural Sciences, 2020, 34(6): 1327-1336.
[31] 霍元子, 田千桃, 徐柵楠, 等. 滸苔對米氏凱倫藻生長的克生作用[J]. 海洋環境科學, 2010, 29(4): 496-499, 508. Huo Yuanzi, Tian Qiantao, Xu Shannan, et al. Allelopathic effects ofon growth of[J]. Marine Environmental Science, 2010, 29(4): 496-499, 508.
Allelopathic effect ofon the growth of
LI Xiao-ying1, CHEN Yi-hua2, ZHAO Yu-chao1, FENG Sen-lei1, LI Wen-qian1, ZHU Qiang1, DONG Zhi-guo1
(1. Jiangsu Key Laboratory of Marine Biotechnology, Jiangsu Ocean University, Lianyungang 222005, China; 2. Shang Hai Collaborative Innovation for Aquatic Animal Genetics and Breeding, Shanghai Ocean University, Shanghai 200090, China)
To investigate the allelopathic effects ofand its metabolites on microalgae in the marine aquaculture environment, we studied the ecological functions ofon the growth ofby using experimental ecological methods. We used three combinations of,metabolites, and a mixture ofand its metabolites at different doses (100 μL, 500 μL, and 1000 μL) to study their effects on the cultivation of. The results showed that in the group with, the algae cell number ofgrew slowly in the first two days with no significant difference, but was lower than that in the control group (>0.05). In 3~4 days, the algal cell number of three groups with different doses was significantly lower than that of the control group (<0.05). In the group with metabolites of, the algal cell number of three groups with different doses had no significant difference with the control group, but it was significantly lower than that of the control group in 2~4 days (<0.05). In the group with the mixture ofand its metabolites, the growth of 100 μL group was slower than that of the other groups. The algae cell number of the 500 μL group and 1000 μL group showed a downward trend, and was significantly lower than that of the control group (<0.05). In the 1000 μL group, the mortality of 100% was achieved on the second day after treatment. In general, studies have shown that the algae cell number in each dose group showed a sequential decrease in,metabolites, and the mixture ofand its metabolites. Moreover, the algae cell number of 1000 μL group ofmetabolites was larger than that in the 500 μL group. The larger dose would indicate a fewer number of algae cells, thus showing a dose effect. The inhibition rates of,metabolites, and the mixture ofand its metabolites were 35.84%, 69.75%, and 100%, respectively. The study showed that the allelopathy ofonshowed an inhibition effect, which further showed a dose effect. This study provided a scientific basis for the evaluation of the effect ofin aquaculture. It also provided a new idea for the management of algae in the red tide.
;; allelopathic effect; growth performance
Jun. 6, 2020
Q789, S917
A
1000-3096(2021)01-0062-08
10.11759/hykx20200606001
2020-06-06;
2020-07-13
江蘇省自然資源發展專項資金海洋科技創新項目(JSZRHYKJ202008);江蘇省農業自主創新項目(CX(20)3150);國家貝類產業技術體系(CARS-49); 江蘇省高等學校自然科學研究重大項目(18KJA240001); 江蘇省漁業科技類項目(Y2018-27)
[Marine Science and Technology Innovation Project of Jiang-su Province Natural Resources Development Special Fund, No. JSZRHYKJ202008; The independent Agricultural Innovation Project of Jiangsu Province, No. CX(20)3150;Modern Agro-industry Technology Research System, No. CARS-49; the Natural Science Foundation of the Jiangsu Higher Education Institutions of China, No. 18KJA240001; Jiangsu Fisheries Science and Technology Project, Y2018-27]
李曉英(1975-), 女(漢族), 黑龍江海倫縣人, 碩士, 實驗師, 研究方向為水產種苗工程, E-mail: 327085439@qq.com; 董志國,通信作者, 教授, E-mail: dzg7712@163.com
(本文編輯: 趙衛紅)
