煙草轉錄因子NtMYB4a響應干旱、低溫和茉莉酸甲酯脅迫的功能分析
2021-02-07 04:46:22蔣悅羅倩姜超英孫光軍聶瓊
作物研究 2021年1期
蔣悅,羅倩,姜超英,孫光軍,聶瓊,2*
(1貴州大學煙草學院,貴陽 550025;2貴州大學農學院,貴陽 550025;3貴州省煙草公司,貴陽 550081)
植物在生長發育的各個階段都會受到各種生物或非生物脅迫的影響而導致減產。為了適應自然界中的各種脅迫,植物在長期的生存與進化中形成了一系列的應對措施,從分子、細胞、生理和生化水平做出適應性調整,以抵御和適應脅迫。如積累甜菜堿、脯氨酸等有益代謝產物來維持細胞內的滲透壓平衡,誘導相關抗逆基因的表達,合成具有特殊功能的酶等來增強植物的抗逆性[1]。在植物抗逆應答過程中,轉錄因子起著關鍵的調控作用。當植物遭受干旱、低溫、鹽等非生物脅迫時,轉錄因子會在轉錄水平和代謝水平迅速做出響應,誘導和調控脅迫響應相關基因的表達[2]。MYB轉錄因子是植物體內最大的一類轉錄因子,大量研究表明其參與非生物脅迫的應答和調控。如過表達SbMYB15的煙草能夠顯著提高其對PEG模擬干旱和鹽脅迫的耐性[3];MaMYBR1在香蕉參與響應低溫、干旱和鹽脅迫中起著重要的作用[4];在鹽和熱脅迫下,Os-MYB340在水稻葉和根中的表達量明顯上調[5];過表達BpMYB4基因能讓白樺在低溫脅迫中產生具有一定的抵抗低溫能力的物質[6];AtMYB49表達量的增加提高了擬南芥葉片中的Ca2+水平,增強了擬南芥抗氧化能力[7];在藍莓愈傷組織中過表達Vc-MYB4a增強了其對鹽、干旱、寒冷、冷凍和熱脅迫的敏感性[8]。
脯氨酸是植物體內最重要的滲透調節物質之一,具有維持植物細胞膜結構穩定性和清除活性氧的作用[9]。在逆境條件下(熱、凍、旱、鹽堿、冷),植物體內脯氨酸的含量顯著增加,而脯氨酸的含量在一定程度上反映了植物的抗逆性,抗旱性和抗寒性強的品種中往往積累較多的脯氨酸[10]。丙二醛是膜脂過氧化的產物之一,它的產生能加劇膜的損傷,其含量與植物的抗性呈負相關[11]。
煙草起源于雨量充沛的熱帶,對生態環境非常敏感,是我國重要的經濟作物之一。發掘煙草中非生物脅迫相關的轉錄因子對通過分子生物學手段增強煙草的抗逆能力具有重要的意義。目前,有關煙草MYB轉錄因子參與非生物脅迫的研究較少。本研究以NtMYB4a過表達和CRISPR/Cas9敲除株系及其野生型煙株為材料,分析了干旱、低溫和茉莉酸甲酯(MeJA)脅迫下NtMYB4a表達變化以及脯氨酸和丙二醛含量的差異,探討了NtMYB4a對干旱、低溫和MeJA脅迫的應答反應和可能參與的抗逆功能,為闡明NtMYB4a的功能和在植物抗逆基因工程中的應用提供依據。
1 材料與方法
1.1 試驗材料
野生型GDH88煙株、NtMYB4a敲除煙株(K)和過表達煙株(O)為貴州大學煙草品質重點實驗室保存。采用漂浮育苗的方式,將播種的育苗盤置于培養液中于28℃的光照培養箱中培養,待幼苗長至6片完整葉時選取長勢一致的壯苗進行脅迫處理。分別將培養盤里的培養液換成50μM的MeJA溶液和20% PEG6000進行MeJA、模擬干旱處理;放入4℃的光照培養箱進行低溫脅迫處理。對照為28℃光照培養箱中培養液培養的煙苗。每個處理20株,3次重復,分別取處理后0、1、3、6、12、24和48 h葉片,分為2份,用液氮速凍后保存于-80℃冰箱備用。
1.2 RNA提取與cDNA合成
以1.1所保存的葉片為材料,采用天根生化科技有限公司的RNAprep pure Plant Kit總RNA提取試劑盒(DP432)提取RNA,采用天根生化科技有限公司的FastKing一步法除基因組cDNA第一鏈合成預混試劑,合成cDNA第一鏈。
1.3 實時熒光定量PCR
以L25(Gen Bank登錄號為L18908)為內參,通過實時熒光定量PCR(RT-qPCR)測定NtMYB4基因的相對表達量。根據其基因序列,采用Primer5軟件設計NtMYB4(Gen Bank登錄號為MN_178131)基因(F:5′-GAAGAAAACCAAAGGAAATGAG-3′,R:5′-GCTGCTGTCTGATGAAGAAAC-3′)和內參基因L25(F:5′-CCCCTCACCACAGAGTCTGC-3′,R:5′-AAGGGTGTTGTTGTCCTCAATCTT-3′)的引物。實時熒光定量PCR反應體系為:2×Talentl qPCR PreMix 12.5μL;正向引物(10μM)0.75μL;反向引物(10μM)0.75μL;cDNA模板2μL;RNasefree ddH2O 9μL,總量25μL。PCR反應程序:94℃預變性30 s,94℃變性5 s,55℃退火30 s,39個循環。每個反應重復3次,采用2-ΔΔCt法計算NtMYB4a的相對表達量。
1.4 脯氨酸和丙二醛的測定
采用酸性茚三酮法測定脯氨酸含量[12],采用硫代巴比妥酸法測定丙二醛含量[13]。
2 結果與分析
2.1 干旱(PEG)脅迫下NtMYB4a表達及脯氨酸和丙二醛含量變化
以野生型煙株為對照,干旱脅迫下,NtMYB4a在過表達和敲除突變體中的表達量變化如圖1-A。敲除突變體中NtMYB4a的表達量在處理前顯著低于對照,干旱處理后,其表達量呈緩慢上升的趨勢,處理后1 h,表達量與對照相當,處理后48 h達到最大值,是對照的2.5倍;過表達突變體中NtMYB4a的表達量在處理前和處理后都極顯著高于敲除突變體和野生型煙株,相對表達量在處理后6 h達到最大值,是對照的2 810倍。
以處理前(0 h)的葉片為對照,過表達、敲除突變體和野生型煙株中NtMYB4a表達量的變化如圖1-B。PEG處理后,敲除突變體中NtMYB4a的表達量迅速升高,在處理后1 h時達到最大值,是處理前的5.76倍,之后迅速下降,到處理后3 h又平緩上升。野生型煙株中NtMYB4a表達量的變化趨勢與敲除突變體一致。而過表達突變體中NtMYB4a的表達量在整個脅迫過程中沒有顯著變化。說明Nt-MYB4a的表達易受干旱脅迫的誘導,但過表達后干旱脅迫的影響變小。
干旱(PEG)脅迫處理下,過表達、敲除突變體和野生型煙株中丙二醛和脯氨酸含量的變化趨勢一致(圖1-C、1-D)。從圖1-C可見,過表達、敲除突變體和野生型煙株中丙二醛含量都在處理后6 h達到最大值,之后緩慢下降。敲除突變體的丙二醛含量顯著高于過表達和野生型煙株,過表達突變體略低于野生型煙株。從圖1-D可見,過表達、敲除突變體和野生型煙株中脯氨酸含量均在處理1 h后隨處理時間的延長呈現上升—下降—上升的趨勢,處理后48 h達到最大值。
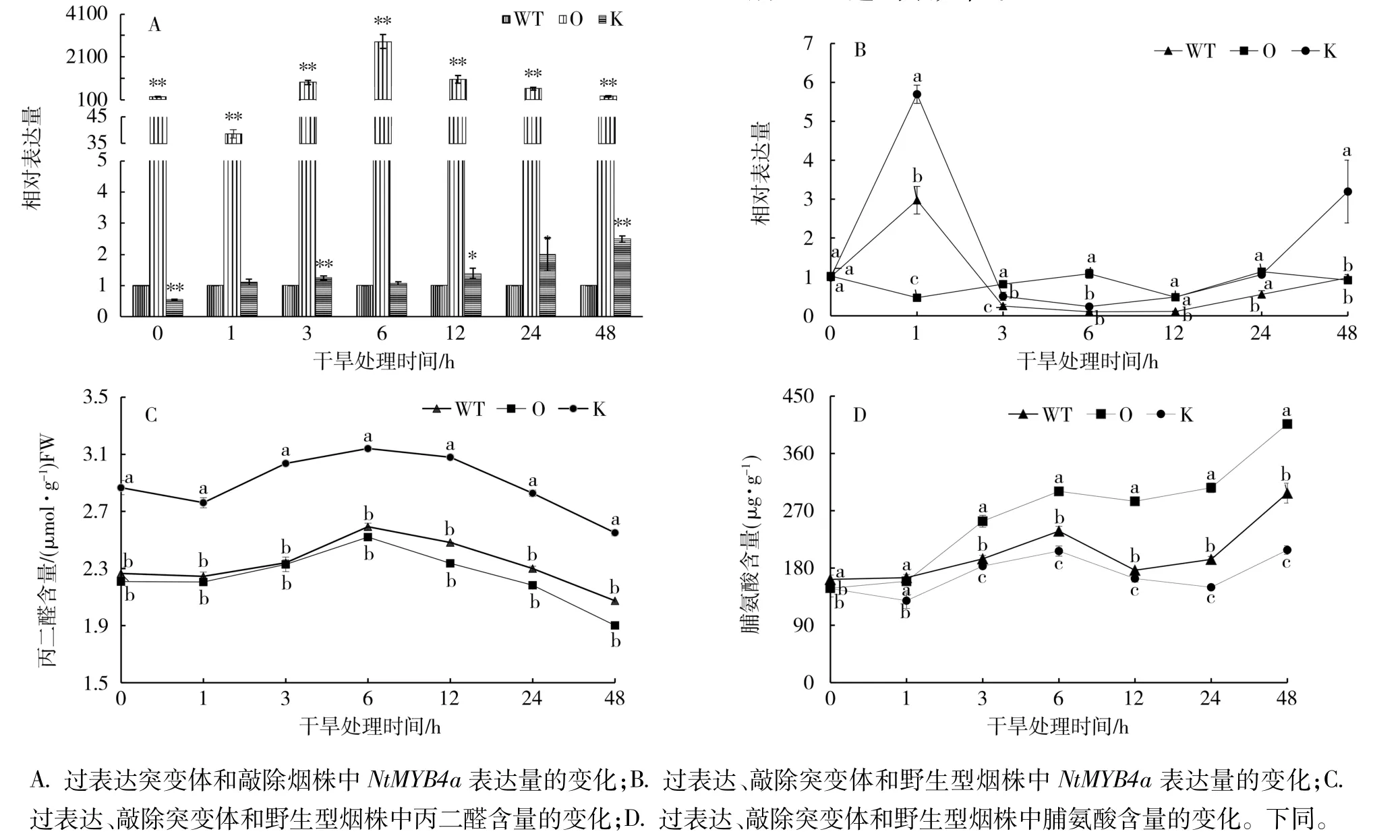
圖1 NtMYB4a對干旱(PEG)脅迫的響應Fig.1 Response of NtMYB4a to drought(PEG)stress
2.2 MeJA脅迫下NtMYB4a表達及脯氨酸和丙二醛含量變化
以野生型煙株為對照,過表達和敲除突變體中NtMYB4a表達量的變化如圖2-A。敲除突變體中NtMYB4a的表達在處理前低于對照,處理后呈現增加—降低—增加—降低的變化趨勢,且在處理后12 h達到最大值,是對照的2.1倍。過表達突變體中NtMYB4a的表達量在處理前后都極顯著高于敲除突變體和野生型煙株,在處理后6 h達到最大值,是對照的495倍。
以0 h的過表達、敲除突變體和野生型煙株為對照,過表達、敲除突變體和野生型煙株中Nt-MYB4a表達量的變化如圖2-B。MeJA脅迫處理后,敲除突變體中NtMYB4a表達量1 h內持續升高,處理后6 h有所回落,之后再次上升,并在48 h表達量達到最大值,是對照的9.79倍,其變化趨勢和野生型煙株一致。而過表達突變體中NtMYB4a表達量變化呈現平緩的趨勢。說明NtMYB4a的表達容易受到MeJA脅迫的誘導,過表達后脅迫的影響變小。
MeJA脅迫處理下,過表達、敲除突變體和野生型煙株中丙二醛和脯氨酸含量的變化趨勢一致(圖2-C和2-D)。從圖2-C可見,過表達、敲除突變體和野生型煙株中丙二醛含量在處理后6 h達到最小值,之后緩慢增加。從圖2-D可見,過表達、敲除突變體和野生型煙株中脯氨酸含量隨著脅迫時間的延長呈現增加—降低的趨勢,都在處理后6 h達到最大值。
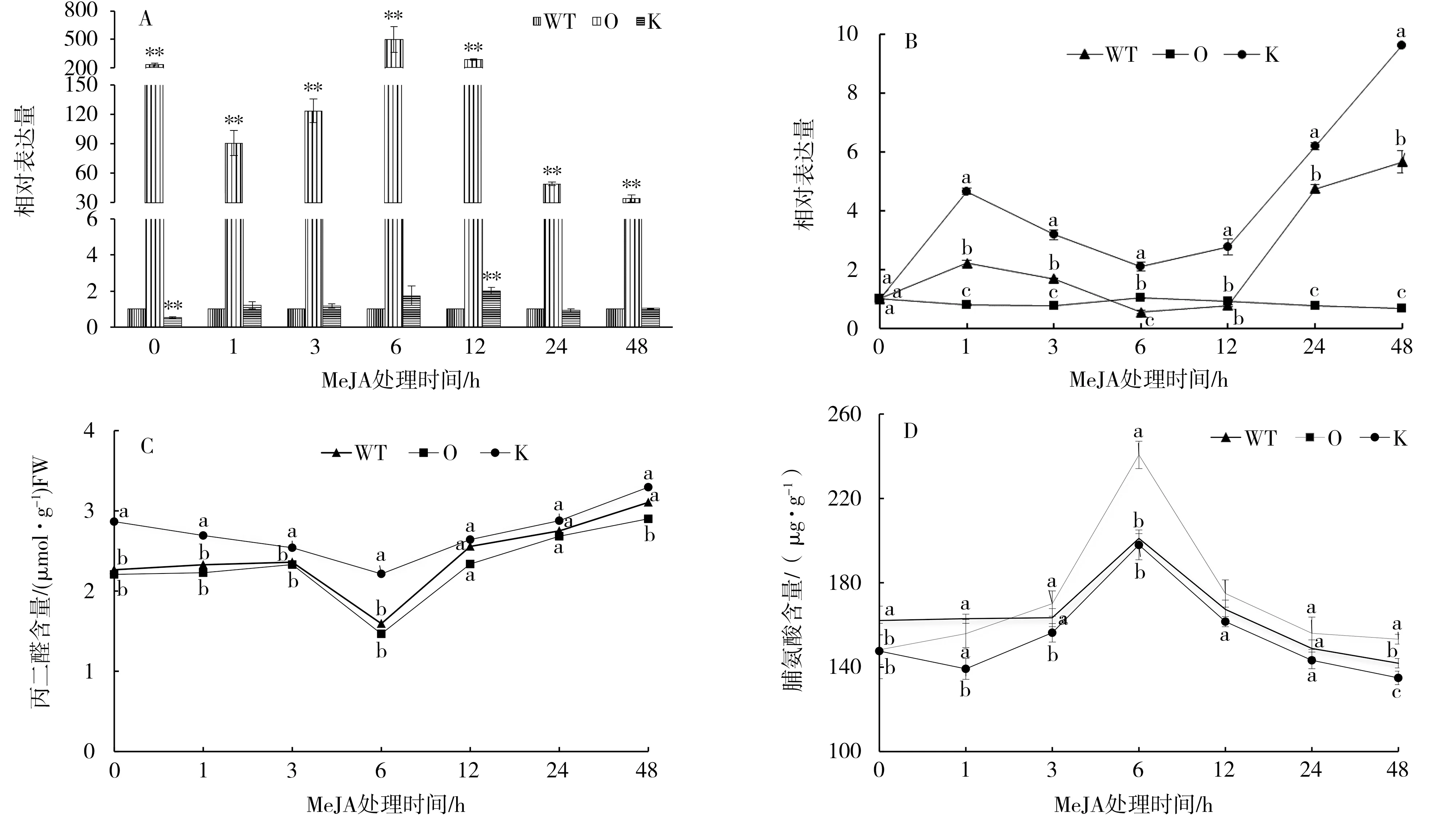
圖2 NtMYB4a對MeJA脅迫的響應Fig.2 Response of NtMYB4a to MeJA stress
2.3 低溫(4℃)脅迫下NtMYB4a表達及脯氨酸和丙二醛含量變化
以野生型煙株為對照,過表達和敲除突變體中NtMYB4a表達量的變化如圖3-A。敲除突變體中NtMYB4a的表達量在處理前低于對照,處理后呈緩慢上升的趨勢,處理后3 h與對照相當,在處理后12 h達到最大值,為對照的1.58倍:過表達突變體中NtMYB4a的表達量處理前后都顯著高于敲除和野生型煙株,在處理前最高,為對照的235.6倍,處理后表達量降低。
以處理前(0 h)為對照,過表達、敲除突變體和野生型煙株中NtMYB4a表達量的變化如圖3-B。低溫處理后,敲除突變體中NtMYB4a的表達量迅速增加,在處理后6 h達到最大值,為對照的10.15倍,之后迅速下降,與野生型煙株中NtMYB4a表達量的變化趨勢一致。而過表達突變體處理前后Nt-MYB4a的表達量無顯著變化。說明NtMYB4a的表達易受低溫脅迫的誘導,過表達后脅迫的影響變小。
低溫脅迫處理下過表達、敲除突變體和野生型煙株丙二醛和脯氨酸含量的變化趨勢不一致(圖3-C和3-D)。從圖3-C可見,敲除突變體丙二醛含量在處理后24 h時達到最大值,顯著高于過表達突變體和野生型煙株,而過表達突變體丙二醛含量低于野生型煙株。從圖3-D可見,過表達、敲除突變體和野生型煙株脯氨酸含量隨著處理時間的延長均呈增加—降低—增加的趨勢,均在處理后48 h達到最大值。
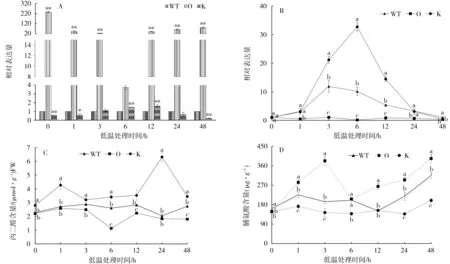
圖3 NtMYB4a對低溫脅迫的響應Fig.3 Response of NtMYB4a to low temperature stress
2.4 NtMYB4a啟動子順式作用元件分析
從NCBI(美國國家生物技術信息中心)煙草基因組數據庫中獲取NtMYB4a起始密碼子上游2 000 bp的序列,利用Plant-CARE數據庫對其順式響應元件進行分析,發現NtMYB4a啟動子含有多個作用元件(表1)。其中最多的是參與光反應的元件,如ATCT-motif、Box4、G-Box、MRE、MRE和TCTmotif;還含有赤霉素響應元件P-box、干旱誘導位點MBS、低溫響應元件LTR、厭氧調節因子ARE、脫落酸反應元件ABRE、60K蛋白結合位點Unnamed-1、生長素響應元件TGA-element、防御和應激反應元件TC-rich repeats、茉莉酸甲酯(MeJA)反應順式作用調節元件CGTCA-motif、啟動子和增強子的共同順式作用元件CAAT-box等。表明煙草NtMYB4a的表達可能受激素、光、低溫、干旱、氧等調節。

表1 煙草NtMYB4a啟動子序列分析結果Table 1 Tobacco NtMYB4a promoter sequence analysis
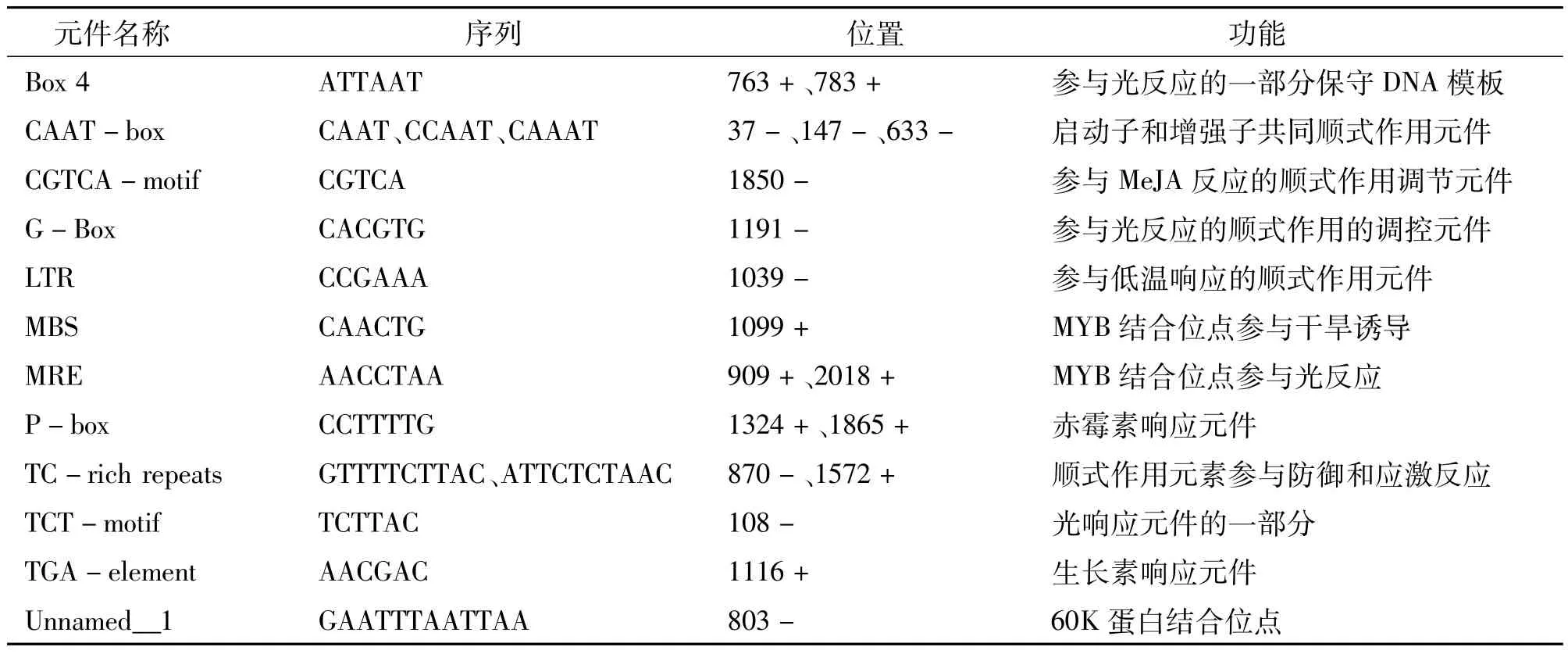
續表1
3 討論
低溫和干旱是作物在生長發育過程中最常見的非生物脅迫,對作物的產量和質量有很大的影響。植物受到脅迫時,外源MeJA能夠激發防御基因的表達,從而誘導植物的化學防御。在所有的生物體內都存在轉錄因子,它通過控制靶基因的轉錄速率來完成生物體的不同代謝過程[14]。當生物遭受脅迫時,轉錄因子上的結構域就會與其他基序結合抑制或激活轉錄以響應內源或外源脅迫。研究表明,過量表達BpMYB4可明顯增強滲透調節物質和保護酶系統在白樺樹中的含量[16];在鹽旱脅迫下,小麥Tamyb31基因在0.5 h就明顯上調表達,說明其參與了早期鹽旱脅迫信號的轉導[15]。當植株遭受脅迫時,體內的滲透調節物質脯氨酸的積累量通常增多,使植株體內滲透勢發生改變,從而提高植株的抗逆性[16]。一般低溫脅迫下抗凍性強的品種脯氨酸的含量顯著高于抗凍性弱的品種[17]。當脅迫持續加劇時,植物自身保護酶系統平衡就會被打破,而產生的大量活性氧離子加劇了對細胞膜的傷害,丙二醛(MDA)等自由基不斷增加,對植物造成毒害作用。因此MDA可以用作間接預測植物抗逆性的指標[18]。
4 結論
本研究通過對煙草NtMYB4a基因在干旱、Me-JA和低溫脅迫下的表達分析及對丙二醛和脯氨酸含量的測定,發現NtMYN4a在不同材料、不同脅迫處理下表達模式各不相同。過表達突變體中Nt-MYB4a表達量很高,在受到脅迫時反應遲鈍,表達量變化不明顯,且脯氨酸含量最高,丙二醛含量最低;而敲除突變體和野生型煙株中NtMYB4a表達量本身較低,在受到脅迫時極其敏感,其表達量變化波動大且變化趨勢相似。但與野生型煙株相比,敲除突變體脯氨酸含量更低,丙二醛含量更高。在干旱、MeJA和低溫脅迫下,敲除突變體中丙二醛含量高于過表達突變體和野生型煙株,脯氨酸含量低于過表達突變體和野生型煙株。隨著處理時間的延長,敲除突變體NtMYB4a表達量上升,同時丙二醛含量呈現下降的趨勢,而脯氨酸作為滲透調節物質則緩慢增加,說明NtMYB4a表達量的改變增強了煙株的抗逆性。進一步分析發現,NtMYB4a的啟動子上有響應低溫、干旱、MeJA及其他防御和應激反應元件。根據實驗結果,推測NtMYB4a參與了煙草對低溫、干旱、MeJA脅迫的應答,可對其在非生物脅迫中的功能及其在煙草抗逆基因工程中的應用進行進一步的研究。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
當代化工研究(2016年9期)2016-03-20 16:22:08
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
聲屏世界(2014年6期)2014-02-28 15:18:09
