根癌農桿菌vbp2基因啟動子轉錄調控的探析
2021-02-10 07:37:34徐楠徐宇娟孫盼宗仁杰郭敏亮
生物技術通報 2021年12期
徐楠 徐宇娟 孫盼 宗仁杰 郭敏亮
(揚州大學生物科學與技術學院,揚州 225009)
根癌農桿菌(Agrobacterium tumefaciens),是一種廣泛存在于土壤中的革蘭氏陰性菌。作為一種常見的植物病原菌,它能夠使大多數雙子葉植物發生冠癭瘤疾病。其致病機制是將自身Ti質粒上的部分DNA片段(簡稱T-DNA)以T-復合物的形式轉運到宿主植物細胞,并整合到宿主的基因組中,使宿主植物細胞發生遺傳轉化而致病[1-3]。由于根癌農桿菌能將基因轉給宿主植物,因此,根癌農桿菌很快就成為了應用最廣泛的植物轉基因工具菌[1-6]。根癌農桿菌的致病(或轉基因)受宿主分泌的糖類、酚類等小分子化合物的誘導[7-9]。當環境中沒有誘導物時,絕大多數與致病有關的基因都不表達,根癌農桿菌不具備致病能力[9]。當誘導物存在時,就可以誘導相關的vir基因表達出一系列致毒蛋白(virulence protein),其中的VirD2蛋白具有核酸內切酶活性,從Ti質粒上切下單鏈的T-DNA,并通過共價鍵與T-DNA的5′-端連接形成VirD2-T-DNA復合物,進而加工成T-復合物[1-3,10-12]。T-復合物通過根癌農桿菌細胞兩端的四型分泌系統(type IV secretion system,T4SS)進入植物細胞,其中VirC1蛋白和VBP(VirD2-binding protein)蛋白可以提高T4SS對T-復合物的招募作用[13-15]。
VBP蛋白,又叫T-復合物招募蛋白,是一種可以與VirD2專一性結合,并參與T-DNA轉運的蛋白[14]。其具體功能是將T-復合物招募到根癌農桿菌細胞膜上的T4SS處,促進T-DNA的轉運。根癌農桿菌基因組中含有3個能編碼VBP的平行同源基因,分別為vbp1(atu5117)、vbp2(atu4860)、vbp3(atu4856)[13-14]。只有3個vbp基因全部敲除,根癌農桿菌的致瘤能力才大幅降低[13],3個同源基因在招募T-復合物上具有功能互補性。但是,從進化的觀點看,相對精簡的原核基因組為一項非必需功能保存3個同源基因,有些不太經濟,因此,我們推出VBP蛋白可能還具有招募T-復合物之外的其它功能[14]。為了進一步探尋VBP的其它功能,我們對3個vbp基因在致瘤誘導條件下的表達情況進行了研究,發現vbp1和vbp3基因的表達不受致瘤誘導條件的影響,而vbp2基因的表達卻受致瘤誘導條件的影響,同時還發現vbp1的缺失也會影響vbp2的表達[16]。這些研究表明3個vbp基因也可能相互之間影響彼此的表達。進一步研究vbp2基因轉錄表達調控的機制,不僅對優化根癌農桿菌轉基因的誘導條件,提高轉基因效率具有實踐指導意義,而且對深入解析VBP蛋白的其它生物學功能也具有重要的參考價值。
本研究在生物信息分析vbp2啟動子結構的基礎上,通過構建原位替代突變株,確定了不同誘導條件對vbp2啟動子的調控作用,以及最適的誘導濃度。根據生物信息學分析預測,通過啟動子截短的方法,確定出誘導作用調控元件所在的區域。
1 材料與方法
1.1 材料
1.1.1 菌株和質粒 根癌農桿菌(A. tumefaciens)C58為胭脂堿型野生菌,vbp1和vbp2缺失的根癌農桿菌菌株為 GMI9017Δvbp2,由 C58 衍生而來[13],大腸桿菌宿主菌為 DH5α(Escherichia coli DH5α);自殺質粒pEX18Km、表達載體pUCA19、pCMV6-AN-RFP(紅色熒光蛋白基因的質粒),eGFP-T(綠色熒光蛋白基因的質粒)均為本實驗室保存。本文所用到的引物如表1所示。
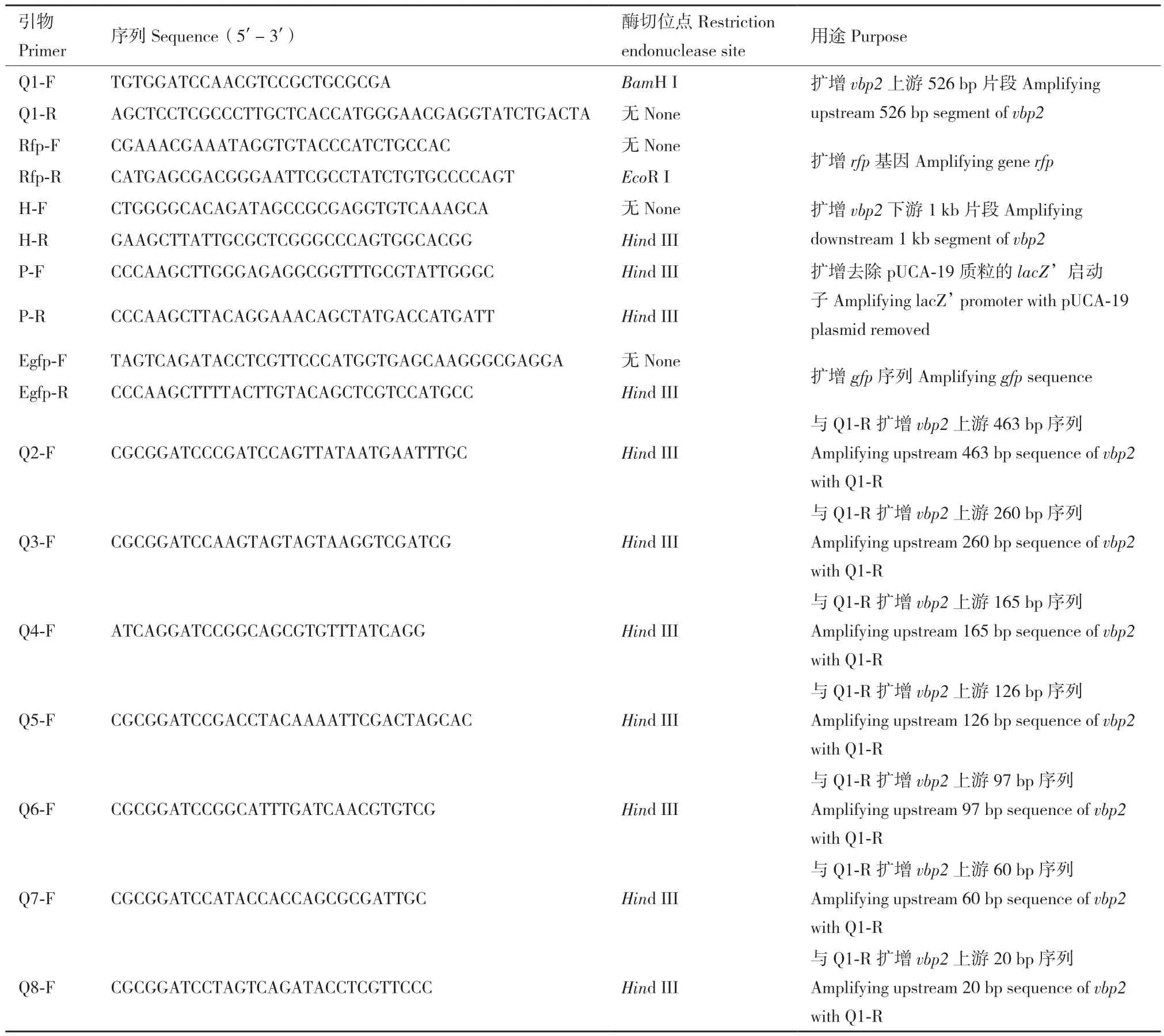
表1 本研究所使用的引物Table 1 Primers used in this study
1.1.2 培養基 大腸桿菌用LB培養基培養,根癌農桿菌用MG/L和AB蔗糖培養基培養,具體配方見文獻[13-14,16]。
1.2 方法
1.2.1 vbp2啟動子預測和分析 在HMM Database(www.ebi.ac.uk/Tools/hmmer/.)、Prokaryote Promoter Prediction(https://services.healthtech.dtu.dk/.) 和TESS(http://agave.humgen.upenn.edu/utess/tess) 網站對A. tumefaciens農桿菌T-復合物招募蛋白VBP2的編碼基因vbp2上游序列中可能存在的轉錄因子結合位點進行預測分析。
1.2.2 vbp2基因閱讀框被紅色熒光蛋白編碼基因rfp準確替代突變體菌株的構建 用紅色熒光蛋白編碼基因rfp準確替代vbp2基因的突變體的構建采用同源重組的方法。該方法的原理和詳細步驟參閱文獻[17]。以 A. tumefaciens C58基因組為模板,Q1-F、Q1-R為引物,通過PCR技術擴增vbp2基因翻譯起始位點上游526 bp的啟動子序列。以帶有紅色熒光蛋白基因的質粒pCMV6-AN-RFP為模板,用引物Rfp-F、Rfp-R擴增紅色熒光蛋白基因rfp。以前兩次PCR產物為模板,以Q1-F、Rfp-R為引物,利用重疊延伸PCR技術構建vbp2上游526 bp片段+ rfp片段融合基因。以A. tumefaciens C58基因組為模板,用引物H-F和H-R擴增出vbp2下游1 kb片段。最后利用引物Q1-F和H-R將vbp2上游526 bp片段+rfp片段與vbp2下游1 kb片段通過重疊PCR連接起來。用TaKaRa瓊脂糖凝膠DNA回收試劑盒,回收擴增的目的片段和用BamH I和Hind III切過的載體pEX18Km。用T4連接酶將兩者4℃過夜連接后轉化至大腸桿菌DH5α,經菌落PCR和質粒雙酶切鑒定正確后送測序,確定測序結果正確后,將所構質粒pEX18Km-rfp電轉至C58感受態細胞,涂布于含卡那霉素抗性的MG/L固體培養基上進行第一次抗性篩選。將長出的單克隆在新的卡那抗性的MG/L固體培養基上純化,以保證其具有卡那抗性。將純化的具有卡那抗性的單克隆劃線于含5%蔗糖的MG/L固體培養基上,若不長則說明賦予了蔗糖敏感性;選取同時具有卡那霉素抗性和蔗糖敏感性的轉化子,劃線于含5%蔗糖的MG/L固體培養基上,進行第二次篩選,此時會出現包括野生型和替代突變株,兩種類型的單菌落,將長出來的菌落在5%蔗糖的MG/L上純化一次,再劃線于含卡那霉素的MG/L固體培養基上,確認卡那抗性和蔗糖敏感性均已丟失。挑取無卡那霉素抗性和蔗糖不敏感的單菌落進行PCR 鑒定,鑒定成功后將產物送測序。
1.2.3 誘導物對vbp2啟動子的誘導及啟動子活性的測定 在取得vbp2基因閱讀框被紅色熒光蛋白編碼基因rfp準確替代的突變體菌株后,vbp2啟動子的轉錄活性就可以通過測定其控制的紅色熒光蛋白RFP的表達量來表征。具體實驗設計是用不同濃度的不同誘導物(乙酰丁香酮AS和鼠李糖Rha)誘導上述突變體菌株。如果vbp2的啟動子能被誘導,而表現出不同的轉錄活性,就可以表達出不同量的紅色熒光蛋白RFP,RFP在細胞粗提液總蛋白中的占比就不同。因此,可以用細胞粗提液中單位總蛋白的熒光強度來表示熒光蛋白的表達量。詳細實驗步驟如下:將上述突變體菌株的單菌落在MG/L培養基中培養至OD600= 0.5-0.7,離心收集菌體后轉移至AB蔗糖培養基過渡培養4-5 h,然后分成若干等份,不同的等份分別用不同濃度的不同誘導物進行誘導培養12-16 h。AS的誘導濃度設為0、50、100、150、200和 250 μmol/L。Rha的誘導濃度設為 0、50、100、150、200 和 250 μmol/L。收集經誘導的細胞,用50 mmol/L PBS緩沖液(pH 7.4)洗滌兩遍,重懸為總體積10 mL的菌體懸浮液,菌體懸浮液通過調整OD600一致,以保證每份經誘導的菌體懸浮液具有相同的細胞量。用超聲波(220 W,3 s on/5 s off,20 min)破碎細胞后,離心獲得菌體細胞粗提液。粗提液中的總蛋白濃度采用考馬斯亮藍染料結合法測定[18]。在測得每份待測樣品的總蛋白濃度之后,用PBS緩沖液將每份待測樣品的總蛋白濃度調成一致。采用Varian Eclipse熒光分光光度計測定每份細菌粗提液的熒光強度,計算單位總蛋白的紅色熒光強度。用553 nm波長的光激發紅色熒光蛋白的熒光,測定其發射出的574 nm波長的光強[19-21]。
1.2.4 vbp2啟動子區被截短的熒光表達載體的構建及被截短的啟動子活性的測定 為了確定個別調控位點在啟動子區的位置,我們對vbp2啟動子區進行截短,得到不同長度的vbp2基因上游片段。將這些不同長度的vbp2基因上游片段用作啟動egfp基因表達綠色熒光蛋白GFP的啟動子,連接到egfp基因起始密碼子的上游。然后將這些連上了不同長度的vbp2啟動子區DNA片段的egfp基因分別插入到沒有啟動子的質粒中,以構建用截成不同長度的vbp2啟動子啟動egfp基因表達的系列表達質粒。無啟動子的質粒構建方法是:以P-F和P-R(均具有Hind III酶切位點)為引物,以pUCA19質粒為模板進行全質粒PCR。之后通過Hind III酶切,T4連接酶連接,得到不含有lacZ’啟動子的pUCA19載體,即pUCA19-N質粒。
根據生物信息學分析的結果,在vbp2編碼區上游設計多條引物,分別為Q1-F、Q2-F、Q3-F、Q4-F、Q5-F、Q6-F、Q7-F、Q8-F(均具有 BamH I酶切位點),它們都可與引物Q1-R組成一對引物,通過PCR擴增相應的啟動子片段,片段大小分別為526 bp、463 bp、260 bp、165 bp、126 bp、97 bp、60 bp、20 bp。然后以eGFP-T質粒為模板,Egfp-F、Egfp-R為引物,通過PCR擴增綠色熒光蛋白egfp基因。利用重疊PCR技術,將上述不同長度的vbp2啟動子片段分別與egfp基因相連,構建受不同長度vbp2啟動子控制的egfp基因系列表達片段。然后將這些表達片段通過BamH I和Hind III酶位點插入到無啟動子質粒pUCA19-N中,即得到了一系列受不同長度vbp2啟動子控制的egfp基因表達質粒。在這些egfp基因表達質粒測序正確后,電轉至根癌農桿菌GMI9017Δvbp2菌株。讓該菌株在誘導物的誘導下由不同長度vbp2啟動子控制egfp基因的表達,收集誘導后的細胞,提取細胞粗提液,測定粗提液中單位總蛋白的綠色熒光強度。用488 nm波長的光激發綠色熒光蛋白的熒光,測定其發射出的508 nm波長的光強[22-23]。
2 結果
2.1 vbp2啟動子結構的生物信息學預測結果
用公共數據庫對vbp2基因啟動子區進行生物信息學預測,結果提示,vbp2的啟動子區可能存在多個轉錄調控元件。預測的調控元件及其位點如圖1所示。主要有4種,分別是σ38因子結合位點、環腺苷酸受體蛋白結合位點(cap/crp)、fadR和rhaS。如果這些生物信息學的預測結果有一定可靠性的話,vbp2(atu4860)基因的表達調控機制將相當復雜,不僅受多種因素調控,也可能受某種未知的調控機制所調節。鑒于vbp2基因可能存在復雜的調控機制,現階段我們僅對幾種最有可能存在的誘導調控機制進行研究。

圖1 生物信息學預測的vbp2基因啟動子區可能存在的轉錄調控元件Fig.1 Feasible transcriptional regulator in the promoter region of vbp2 gene predicted by bioinformatics
2.2 vbp2基因被rfp基因原位替代突變體菌株的構建
為了能夠準確地表征vbp2啟動子啟動轉錄的活性,我們將vbp2啟動子控制的vbp2基因替換成編碼紅色熒光蛋白的rfp基因,即可通過測定紅色熒光蛋白的表達量來表征該啟動子的活性。因此,構建vbp2基因被rfp基因原位替代的突變體菌株就很重要。首先需要構建用于根癌農桿菌進行同源重組突變的質粒。圖2所示是已經構建好的用于根癌農桿菌進行同源重組突變的質粒pEX18Km-rfp的酶切和PCR鑒定結果。測序結果進一步驗證該質粒構建正確。將該質粒電轉至根癌農桿菌C58感受態細胞中,通過同源重組整合至根癌農桿菌基因組中。經第一次卡那霉素抗性正向篩選和第二次蔗糖敏感反向篩選,得到無卡那霉素抗性和蔗糖不敏感的原位替代菌株。對所獲得的突變體菌株,通過PCR和測序,以確定所獲突變體菌株準確無誤,并將所獲的vbp2基因被rfp基因原位替代的突變體菌株取名為C58vbp2→rfp菌株。

圖2 用于進行同源重組構建根癌農桿菌突變體的質粒pEX18Km-rfp的鑒定Fig.2 Identification of plasmid pEX18Km-rfp used for the construction of A. tumefaciens mutant via homologous recombination
2.3 根癌農桿菌粗提液中熒光蛋白濃度與熒光強度的相關性驗證
為了明確根癌農桿菌粗提液中的其它成分是否會干擾熒光蛋白的測定,我們將野生型C58(不含熒光蛋白)和替代菌C58vbp2→rfp(含熒光蛋白)的粗提液中的蛋白濃度調成一致,并按不同比例混合,獲得含RFP蛋白粗提液占比分別為0%、20%、40%、60%、80%和100%的混合粗提液,并用熒光分光光度計測定這些混合粗提液的熒光強度。粗提液中RFP的比例與熒光強度之間的關系見圖3,由圖可知,粗提液的熒光強度與粗提液中RFP蛋白的含量呈線性關系,說明粗提液中的其它物質不會干擾RFP蛋白的熒光強度的測定。可以用粗提液的熒光強度來表示熒光蛋白的表達量。

圖3 根癌農桿菌粗提液中RFP的含量與粗提液的熒光強度的關系Fig.3 Relationship between fluorescent intensity and RFP content in A. tumefaciens crude extract
2.4 乙酰丁香酮(AS)和鼠李糖(Rha)能夠誘導 vbp2啟動子啟動熒光蛋白的表達
乙酰丁香酮(AS)是最常用來誘導根癌農桿菌轉基因的誘導物,同時VBP蛋白的一項重要功能就是將T-復合物招募到四型分泌系統處。因此,我們首先想到的就是AS是否能夠調控vbp2啟動子的活性。用不同濃度的AS誘導替代菌C58vbp2→rfp,提取這些經不同AS濃度誘導的細胞粗提液,測定這些細胞粗提液的熒光強度。從圖4-A可以看出,熒光強度所代表的由vbp2啟動子啟動表達的RFP的量是受AS誘導調控的。當AS濃度為150 μmol/L時,誘導vbp2啟動子啟動rfp基因表達的能力最強。
多種單糖也是誘導根癌農桿菌轉基因致瘤的誘導物。前面的生物信息學預測,vbp2啟動子可能存在一個RhaS蛋白結合位點,在大腸桿菌中RhaS與L-鼠李糖結合后可以激活相關基因的啟動子,啟動相關基因的表達。因此,我們測試了鼠李糖對vbp2啟動子控制的RFP表達的影響。圖4-B的結果顯示鼠李糖確實對vbp2基因啟動子控制的RFP的表達有調控作用。誘導vbp2基因啟動子控制的RFP表達的最適鼠李糖濃度為100 μmol/L。

圖4 不同濃度的乙酰丁香酮(A)和L-鼠李糖(B)誘導vbp2啟動子表達RFP的情況Fig. 4 vbp2 promotor-promoting RFP expressions induced by different concentrations of AS(A)or Rha(B)
2.5 vbp2啟動子中AS和Rha調控元件位置的確定
為了進一步確定對AS和Rha誘導做出響應的調控元件在vbp2啟動子區的具體位置。我們對vbp2的啟動子區域進行了截短,被截短片段的長度和被截去的可能調控元件如圖5-A所示。讓這些不同長度的vbp2啟動子片段控制egfp基因的表達。再將這些帶有不同長度的vbp2啟動子片段的egfp基因分別插入到沒有啟動子的質粒pUCA19-N中,獲得egfp基因分別受不同長度的vbp2啟動子片段控制的系列egfp表達質粒。然后將這些egfp表達質粒分別電轉至根癌農桿菌GMI9017Δvbp2感受態細胞,將長出來的菌落進行PCR鑒定,確定這些egfp表達質粒已經分別轉入根癌農桿菌GMI9017Δvbp2菌株的細胞內。鑒定結果見圖5-B,電泳條帶a-h對應的片段大小分別 為 1 246 bp、1 183 bp、980 bp、885 bp、846 bp、817 bp、780 bp和740 bp,與預計相符,說明攜帶受不同長度的vbp2啟動子片段控制的egfp表達質粒的系列根癌農桿菌菌株構建成功。

圖5 受不同長度的vbp2啟動子片段控制的egfp表達質粒的構建和鑒定。Fig.5 Construction and identification of the egfp expression plasmids controlled by different lengths of vbp2 promotor
在取得了這些攜帶受不同長度的vbp2啟動子片段控制的egfp表達質粒的根癌農桿菌菌株后,我們分別用最適濃度的AS(150 μmol/L)和Rha(100 μmol/L)誘導這些菌株,測定這兩種物質誘導不同長度的vbp2啟動子啟動egfp基因表達綠色熒光蛋白GFP的情況。從圖6-A可以看出,當vbp2啟動子區縮短至起始密碼子上游260 bp處時,受其控制的egfp基因仍然能夠被AS誘導表達出綠色熒光蛋白,當vbp2啟動子區縮短至起始密碼子上游165 bp處時,AS就不能誘導啟動子啟動egfp基因表達了。由此可以得出響應AS誘導的調控元件位于vbp2上游的165 bp-260 bp區域。圖6-B是L-鼠李糖(Rha)誘導這些菌株表達GFP的情況,結果顯示,當vbp2啟動子區縮短至起始密碼子上游165 bp處時,Rha仍然能夠誘導GFP的表達,當vbp2啟動子區縮短至起始密碼子上游126 bp處時,Rha就誘導不了GFP的表達,說明,響應Rha誘導的調控元件位于vbp2上游的126 bp-165 bp區域。
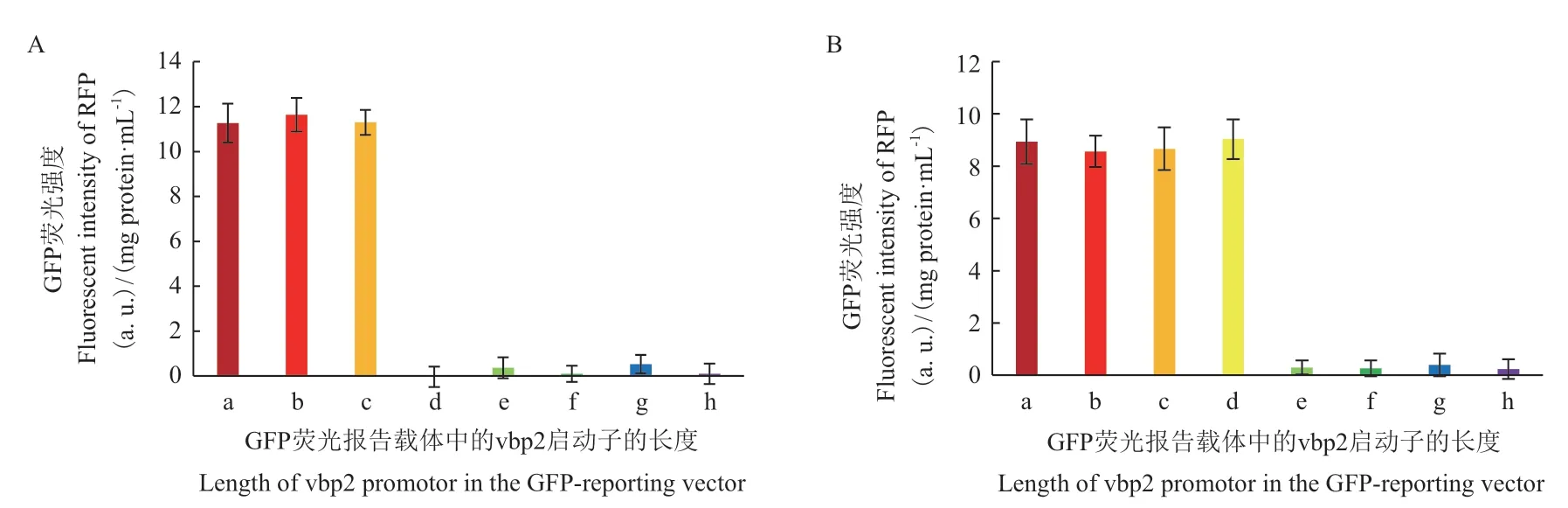
圖6 分別以150 μmol/L AS(A)和100 μmol/L Rha(B)誘導不同長度vbp2啟動子表達GFP的情況Fig. 6 GFP expression promoted by different lengths of vbp2 promotor under the induction of 150 μmol/L AS(A)or 100 μmol/L Rha(B)
3 討論
本研究通過構建將vbp2基因的開放閱讀框(ORF)準確地替換成rfp基因ORF的菌株來定量地研究vbp2啟動子的活性,使紅色熒光基因rfp在vbp2基因的原位上由vbp2基因的啟動子調控rfp基因的表達,這樣不僅可以準確地定量vbp2啟動子的活性,而且可以真實地還原根癌農桿菌在接收到誘導物信號之后是如何啟動vbp2基因表達的。通過細菌粗提液中熒光蛋白的含量與熒光強度的相關性測定,證明了根癌農桿菌細胞粗提液中的其它物質對熒光強度的測定沒有干擾。用AS和Rha兩種物質分別對原位替代菌株誘導,發現這兩種物質都能誘導vbp2啟動子啟動rfp基因表達RFP蛋白。
利用截斷vbp2啟動子區的方法,進一步確定了響應AS誘導的調控元件位于vbp2上游的165 bp-260 bp區域。生物信息學分析預測,vbp2啟動子的165 bp-260 bp區域含有一個σ38因子結合位點。因此,我們推測AS可能是通過影響σ38因子與該位點的結合來調控vbp2基因的啟動子。在其它革蘭氏陰性菌中的研究結果表明,σ38因子主要識別響應環境脅迫的基因的啟動子,特別是識別一些與致病性(virulence)有關基因的啟動子[24-26]。這些實驗結果與AS是誘導根癌農桿菌致病的最好誘導物的推測是非常吻合的[8,27]。AS誘導根癌農桿菌的主要致病基因表達的機制已經很清楚,那就是AS通過VirA/VirG雙組分系統誘導位于Ti質粒上的vir基因表達[28-29]。VBP蛋白也影響根癌農桿菌的致病,但vbp2基因位于根癌農桿菌的線型染色體上,不在Ti質粒上[13]。在這里我們證明AS也能誘導vbp2基因的表達,而且可能是通過影響σ38因子與啟動子的結合來調控vbp2基因的表達,這與已知的AS誘導調控根癌農桿菌致病基因表達的機理是完全不同的,因此,值得我們進一步研究和探討。
通過截斷vbp2啟動子區的方法,我們還確定了響應Rha誘導的調控元件位于vbp2上游的126 bp-165 bp區域。生物信息預測結果顯示,該區域可能含有一個rhas位點。在大腸桿菌中,啟動子區rhaS位點的調控機制是RhaS蛋白需要與L-鼠李糖結合后才具有活性[30-31]。因此,下一步我們希望能夠表達根癌農桿菌的RhaS蛋白,并驗證根癌農桿菌的RhaS蛋白是否能與L-鼠李糖形成復合物,并能夠特異性地結合vbp2啟動子區的這段DNA。此外,現有的研究還表明,RhaS蛋白調控的基因與細菌的碳源利用有關,調節處于不同生存條件(如病原菌的獨立生存或作為病原菌生存)下的碳源利用途徑[31-32]。我們以前的研究結果推測,VBP蛋白除了招募T-復合物到四型分泌系統處外可能還有其它生物學功能[14,16]。因此,根據這些研究結果和合理分析,我們初步推測,L-鼠李糖誘導vbp2基因表達的另一項功能可能是使根癌農桿菌適應宿主體內的碳源利用途徑。當然,還需要更多的實驗證據來支持這樣的推測。
4 結論
證明了vbp2基因分別受乙酰丁香酮和L-鼠李糖誘導,并確定了兩種誘導物的最適誘導濃度。同時,明確了vbp2啟動子區受乙酰丁香酮和L-鼠李糖誘導的調控元件分別位于vbp2起始密碼子上游165-260 bp和126-165 bp區域。