雷公藤紅素通過抑制顆粒細胞炎癥因子基因表達改善肥胖小鼠生育能力的研究
2021-02-27 10:14:56宋東紅馬利方于洋
天津醫藥 2021年2期
宋東紅,馬利方,于洋
肥胖及其誘發的代謝紊亂已成為世界性公共衛生問題,特別是對女性生殖健康有負面影響,包括增加月經失調、無排卵和其他生育問題的風險[1-2]。此外,越來越多的證據表明,在超重或肥胖的不孕女性中,促排卵和輔助生殖技術(如體外受精)的成功率通常較低。肥胖增加了不孕癥的治療成本,患者需要使用更大劑量的促性腺激素[3-4],且胚胎著床率和妊娠率降低[5-7],流產風險增高[8],接受輔助生殖助孕治療的受孕率降低[9]。研究表明,肥胖患者胚胎質量下降與其顆粒細胞功能失調具有密切關系,改善其顆粒細胞功能,可以提高肥胖患者的卵母細胞和胚胎發育潛能[10-11]。
雷公藤紅素是雷公藤等中藥天然存在的一種五環三萜類的生物活性成分[12],在中醫領域用于治療發熱、關節疼痛、水腫及炎癥,包括類風濕性關節炎和系統性紅斑狼瘡等[13]。近年來研究表明,雷公藤紅素作為瘦素增敏劑可調節體質量,具有成為新型減肥藥物的潛力,同時可以緩解肥胖導致的糖尿病胰島素抵抗等病理表型[14]。但是,雷公藤紅素能否直接作用于肥胖患者卵巢局部微環境,改善因肥胖導致的顆粒細胞功能失調,提高卵母細胞及胚胎發育潛能目前仍不清楚。本研究使用雷公藤紅素干預肥胖小鼠模型,探討其對顆粒細胞功能的影響,以及改善肥胖小鼠生育能力的作用機制。
1 材料與方法
1.1 主要試劑與儀器 實驗動物SPF級CD-1小鼠均購自北京維通利華實驗動物科技有限公司。其中雌性5周齡62只,體質量21.4~23.2 g;雌性8 周齡20 只,體質量27.2~28.5 g;雄性10周齡10只,體質量30.3~31.6 g;雄性結扎小鼠10周齡10只,體質量29.5~30.3 g。雷公藤紅素(C0869)、細胞松弛素B(C6762)、體外操作液M2(M7167)、水合氯醛(23100)均購自美國Sigma公司,孕馬血清促性腺激素、人絨毛膜促性腺激素(寧波第二激素廠),透明質酸酶(美國Sigma 公司,H4272),總RNA提取試劑盒(美國Invitrogen公司,AM1921),cDNA逆轉錄試劑盒(德國Qiagen 公司,205111),KSOMaa 培養基(德國Merck-Millipore公司,MR-020P-D),熒光定量PCR試劑盒(TIANGEN 公司,FP303-01),CO2培養箱(美國Thermo Fisher Scientific 公司,3111),體式顯微鏡(日本尼康,SMZ1000),倒置顯微鏡(日本尼康,Ti-E),顯微操作系統(日本Narishige,NT-88-V3),顯微注射系統(日本PRIME-TECH,PMM-150),實時熒光定量核酸擴增儀(美國Thermo Fisher Scientific 公司,ABI 7900),納米滴分光光度計(美國Thermo Fisher Scientific 公司,NanoDrop2000),凝膠成像系統(美國Bio-Rad公司,Gel Doc XR),離心機(德國Eppendorf公司,5810)。
1.2 方法
1.2.1 實驗分組 62只5周齡雌性CD-1小鼠采用拋硬幣隨機方法分為正常飲食組(n=26)、高脂飲食組(n=26)和雷公藤紅素組(n=10);正常飲食組和高脂飲食組再通過拋硬幣隨機方法分為正常飲食A組(n=10)、正常飲食B組(n=6)、正常飲食C 組(n=10),高脂飲食A 組(n=10)、高脂飲食B 組(n=6)和高脂飲食C組(n=10)。
1.2.2 肥胖小鼠模型的構建 正常飲食組小鼠喂食正常飼料;高脂飲食組和雷公藤紅素組小鼠喂食高脂肪飲食(能量比:62%脂肪,18%蛋白質,20%碳水化合物)。高脂飲食組和雷公藤紅素組小鼠體質量大于正常飲食組小鼠平均體質量20%時,被認為肥胖小鼠模型造模成功。所有動物被單籠飼養,環境溫度為22~26 ℃,相對濕度為50%~60%,光暗周期為12 h/12 h,自由采食和飲水。6周后,對小鼠稱質量并確定肥胖模型造模是否成功。本研究得到北京大學第三醫院倫理委員會的批準,并遵守《實驗動物護理和使用指南》。
1.2.3 雷公藤紅素給藥方法 用含5%二甲基亞砜(DMSO)和1%吐溫20(Tween-20)的生理鹽水溶解雷公藤紅素粉末制備成的雷公藤紅素使用液(含雷公藤紅素500 mg/L)。雷公藤紅素組按照100 μg/(kg·d)的標準[15],每日下午4:00 采用灌胃方法給藥,連續給藥21 d。正常飲食C組與高脂飲食C 組同期以灌胃的方式給予等體積的含5% DMSO 和1%Tween-20的生理鹽水。3組小鼠每日下午3:00稱體質量。
1.2.4 交配實驗檢測小鼠產仔數 將正常飲食A 組與高脂飲食A組按照1∶1的比例,與10周齡雄性CD-1小鼠合籠,次日檢查雌性小鼠陰栓。有陰栓的小鼠移入新籠中單獨飼養,沒有陰栓的小鼠繼續合籠,直到檢查到陰栓為止。19.5 d 產仔后,記錄產仔數。
1.2.5 小鼠顆粒細胞及卵母細胞收集 正常飲食B組、高脂飲食B組、正常飲食C組、高脂飲食C組和雷公藤紅素組小鼠(其中正常飲食C 組、高脂飲食C 組和雷公藤紅素組6 只小鼠,均采用拋硬幣隨機分組方法,從每組10只小鼠中隨機抽取)腹腔注射10 IU 孕馬血清促性腺激素(PMSG),48 h 后腹腔注射10 IU 人絨毛膜促性腺激素(hCG)。注射hCG 后14~16 h,處死小鼠,取出輸卵管,顯微鏡下劃開壺腹部收集卵母細胞—卵丘顆粒細胞復合物。將卵母細胞—卵丘顆粒細胞復合物置于1 mg/L透明質酸酶中3 min,直到卵丘細胞分散。將散落在透明質酸酶中的卵丘顆粒細胞轉移到1.5 mL 試管中,在200×g 條件下離心5 min,去除上清液,將細胞懸浮在KSOMaa培養基中,在37 ℃、5%CO2的培養箱中培養。
1.2.6 實時熒光定量PCR(qPCR)法檢測小鼠顆粒細胞基因表達水平 使用RNA 分離試劑盒從1.2.5中5組小鼠顆粒細胞中提取總RNA。使用納米滴分光光度計測定分離RNA的濃度和質量。用cDNA逆轉錄試劑盒合成目的基因cDNA,利用熒光定量PCR 儀檢測目的基因的表達。用GADPH 作為PCR 內對照。PCR 反應體系的擴增條件:95 ℃變性120 s,60 ℃退火60 s,72 ℃延伸60 s,循環40次。用2-ΔΔCt法計算目的基因的相對表達。所有實驗至少重復3次。檢測的目的基因包括:B 細胞淋巴瘤-2 相關X 蛋白編碼基因(Bax)、BCL2拮抗/殺傷因子1(Bak1)、FAS相關死亡結構域蛋白編碼基因(Fadd)、腫瘤壞死因子受體相關因子2(Traf2)、半胱氨酸天冬氨酸蛋白酶8(Casp8)、半胱氨酸天冬氨酸蛋白酶9(Casp 9)、細胞色素P450-家族11-亞家族A-肽1(Cyp11a1)、細胞色素P450-家族19-亞家族A-肽1(Cyp19a1)、白細胞介素(IL)-1β和IL-6。其中Bax、Bak1、Fadd、Traf2、Casp8和Casp9反映顆粒細胞凋亡情況,Cyp11a1 和Cyp19a1 反映雌激素合成情況,IL-1β 和IL-6 反映顆粒細胞炎癥反應激活情況。引物序列見表1。
1.2.7 小鼠胚胎發育情況檢測 取10周齡CD-1雄鼠,斷頸法處死后從附睪中取出精子,放入含有5 mg/L 細胞松弛素B的M2 體外操作液中。用注射針吸取一個精子,注射入小鼠卵母細胞中。注射后的卵母細胞放入KSOMaa培養液中繼續培養。6 h后,在200倍倒置顯微鏡下觀察出現原核的受精卵數量,統計受精率(受精率=原核受精卵數÷卵母細胞總數×100%)。24 h后在200倍倒置顯微鏡下觀察2細胞胚胎數量,統計卵裂率,卵裂率=卵裂胚胎數÷卵母細胞總數×100%。3.5 d 后在200 倍倒置顯微鏡下觀察囊胚數量,統計囊胚率,囊胚率=囊胚數÷卵母細胞總數×100%。
1.2.8 胚胎移植 將8 周齡的CD-1 雌鼠與10 周齡CD-1 結扎雄鼠合籠交配,次日檢查陰栓。將具有陰栓的雌鼠單獨飼養2 d,即為假孕2.5 d 的小鼠,待用。移植當天,給小鼠腹腔注射4%水合氯醛,麻醉后,剪開皮膚和肌肉層,暴露出子宮,利用玻璃針將囊胚移入假孕2.5 d小鼠的子宮中,17.5 d仔鼠出生后,計算產仔數,產仔數與移植囊胚數的比值即為出生率。
1.3 統計學方法 采用SPSS 25.0 統計軟件進行統計分析。定量數據均采用±s表示,2組間比較采用t檢驗,多組間比較采用單因素方差分析,多重比較采用LSD-t 檢驗。P<0.05為差異有統計意義。
2 結果
2.1 肥胖對小鼠產仔數及顆粒細胞相關基因表達的影響 高脂飲食A 組小鼠11 周齡體質量顯著高于正常飲食A組(P<0.01),每窩平均產仔數顯著低于正常飲食A 組(P<0.01),見表2。與正常飲食B組比較,高脂飲食B 組小鼠顆粒細胞IL-1β、IL-6、Bax、Bak1、Fadd、Traf2、Casp8、Casp9表達顯著升高,而Cyp11a1和Cyp19a1表達顯著降低(均P<0.01),見表3。
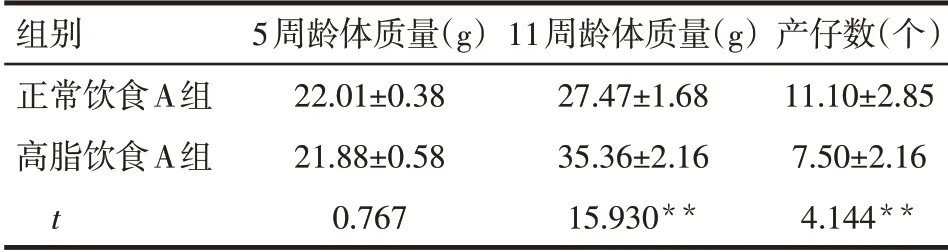
Tab.2 Effects of normal and high-fat diet on the body weight and litter size of mice表2 正常飲食與高脂飲食對小鼠體質量及產仔數的影響(n=10,x±s)
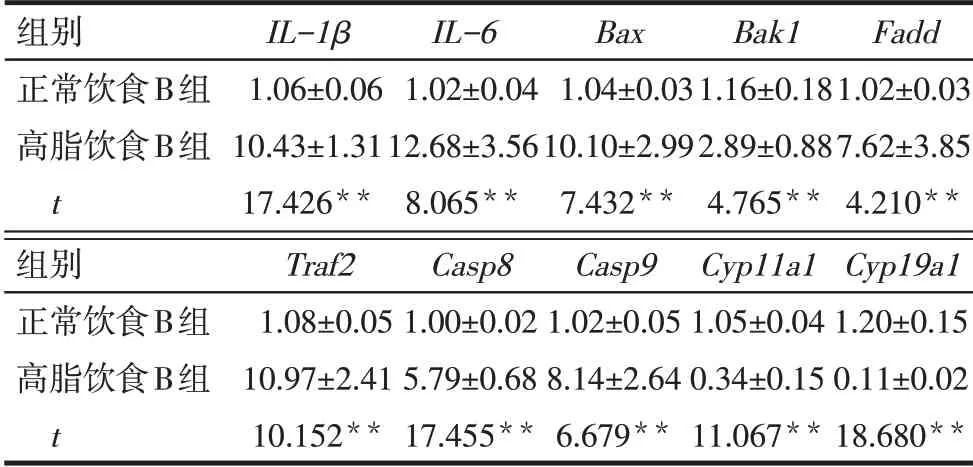
Tab.3 Effects of normal and high-fat diet on the gene expression levels of mouse granulosa cells表3 正常飲食與高脂飲食對小鼠顆粒細胞相關基因表達的影響 (n=6,x±s)
2.2 雷公藤紅素對肥胖小鼠顆粒細胞相關基因表達的影響 雷公藤紅素組與高脂飲食C組小鼠體質量差異無統計學意義,見表4。與高脂飲食C組相比較,雷公藤紅素組小鼠顆粒細胞IL-1β、IL-6、Bax、Bak1、Fadd、Traf2、Casp8、Casp9表達顯著降低(P<0.01),而Cyp11a1和Cyp19a1表達顯著升高(P<0.01)。與正常飲食C組小鼠相比,雷公藤紅素組小鼠顆粒細胞Il-1β、Il-6、Bax、Bak1、Casp8、Cyp11a1和Cyp19a1表達差異無統計學意義(P>0.05),而Fadd、Traf2、Casp9表達顯著升高(P<0.05),見表5。
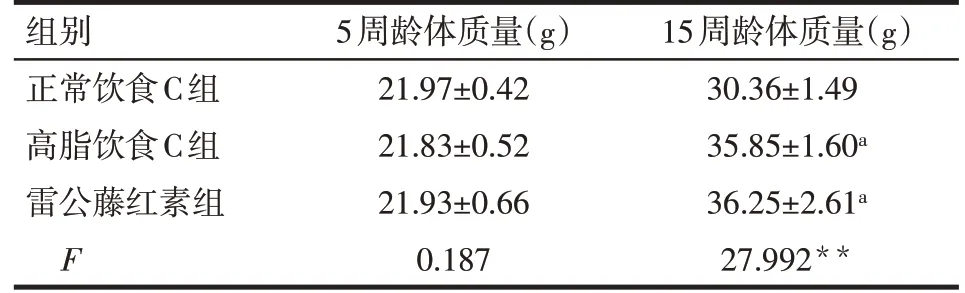
Tab.4 Effects of celastrol diet on the body weight of mice in normal,high-fat and celastrol-diet groups表4 喂食雷公藤紅素對正常飲食、高脂飲食和雷公藤紅素組小鼠體質量的影響 (n=10,x±s)
2.3 雷公藤紅素對小鼠生育能力影響 雷公藤紅素組和正常飲食C組小鼠排卵數、卵裂率、囊胚率和出生率均顯著高于高脂飲食C組(P<0.05),雷公藤紅素組和正常飲食C 組差異無統計學意義。3 組小鼠受精率差異無統計學意義(P>0.05)。見表6。
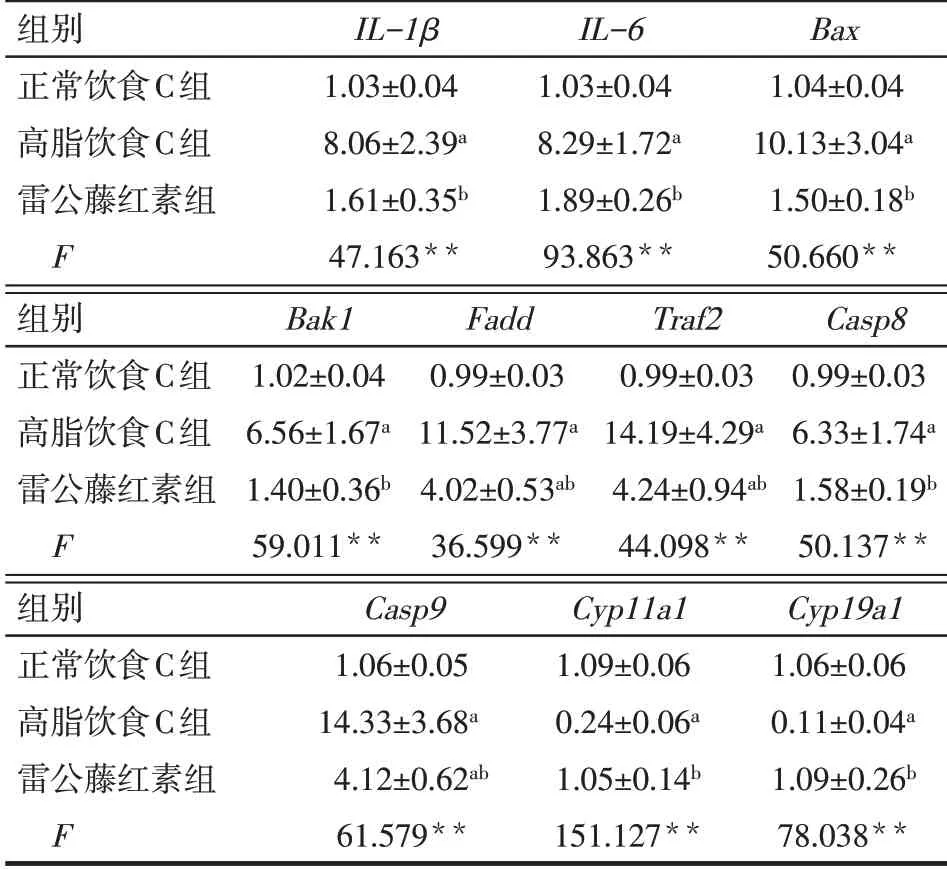
Tab.5 Effects of celastrol diet on the gene expression levels of mouse granulosa cells in normal,high-fat and celastrol-diet groups表5 喂食雷公藤紅素對正常飲食、高脂飲食和雷公藤紅素組小鼠顆粒細胞基因表達的影響(n=6,x±s)
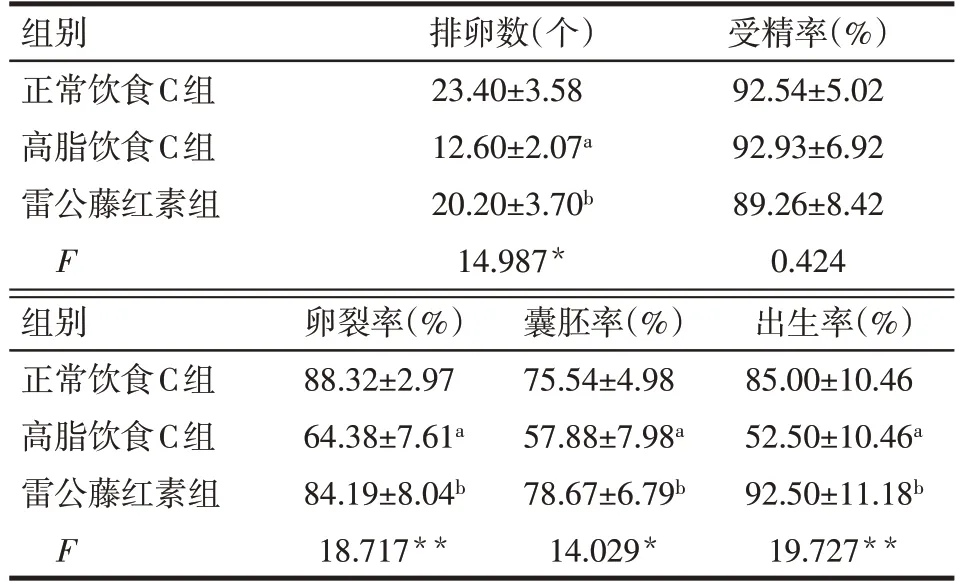
Tab.6 Effects of celastrol diet on the ovulation,fertilization,embryonic development and birth rate of mice in normal,high-fat and celastrol-diet groups表6 喂食雷公藤紅素對正常飲食、高脂飲食和雷公藤紅素組小鼠排卵、受精、胚胎發育及出生率的影響(n=6,x±s)
3 討論
3.1 肥胖與女性不孕 肥胖是導致女性內分泌失調,進而誘發不孕癥的主要因素之一。臨床研究表明,肥胖女性的激素分泌異常表現為雌激素水平顯著升高,而孕激素水平顯著降低[16]。同時,肥胖女性的流產率增加[8],即便在采用輔助生殖助孕技術后,其胚胎發育率、優質胚胎率仍然較非肥胖患者顯著降低[5-6,9]。目前,臨床上針對肥胖不孕癥患者仍然以降低體質量、調節機體內分泌等整體治療為主,而缺乏對改善卵巢局部功能的治療方式的深入探索。本文探究了一種具有多種生物活性的天然產物雷公藤紅素對小鼠卵巢顆粒細胞的作用,結果顯示雷公藤紅素能夠通過調控炎癥因子基因的表達,改善顆粒細胞的凋亡與激素合成相關基因的表達模式,進而提高小鼠生育能力。
3.2 肥胖引起顆粒細胞炎癥、凋亡及對雌激素合成基因表達的影響 顆粒細胞是女性卵巢內重要的細胞類型之一,其不僅為卵母細胞發育提供必須的代謝產物,同時也是雌激素和雄激素的重要合成場所。既往研究表明,肥胖女性體內炎癥基因水平增高,機體處于慢性炎癥狀態[17]。本研究檢測小鼠卵巢顆粒細胞中炎癥因子基因IL-1β和IL-6的表達水平,發現高脂飲食組小鼠顆粒細胞中兩個炎癥因子基因的編碼基因表達水平顯著上調,提示肥胖可誘導顆粒細胞產生炎癥反應。在腫瘤等疾病中,炎癥反應已被證實可以誘導細胞凋亡[18]。本研究中,細胞凋亡相關基因Bax、Bak1、Fadd、Traf2、Casp8與Casp9的表達水平在高脂飲食組小鼠顆粒細胞中均顯著上調,提示肥胖可誘導顆粒細胞發生細胞凋亡。細胞凋亡主要通過兩種方式進行,即線粒體途徑和死亡受體途徑。Bax、Bak1和Casp8屬于線粒體途徑的凋亡相關分子,而Fadd、Traf2和Casp9屬于死亡受體途徑的凋亡相關分子。本研究提示,高脂飲食誘導的肥胖會同時通過兩條經典細胞凋亡途徑誘導顆粒細胞發生凋亡。Cyp11a1和Cyp19a1是雌激素合成過程中重要的限速酶的編碼基因,其表達量的降低可導致雌激素分泌水平降低。本研究結果表明,高脂飲食組小鼠Cyp11a1和Cyp19a1基因表達水平顯著下調,這與既往研究結論一致[19-20]。以上研究結果說明,肥胖會通過誘發顆粒細胞促炎因子的表達,激活顆粒細胞的炎癥反應,誘導顆粒細胞凋亡,同時降低顆粒細胞激素合成限速酶編碼基因表達水平,影響顆粒細胞的激素分泌功能,進而導致肥胖小鼠產仔數的降低。
3.3 雷公藤紅素可改善肥胖小鼠顆粒細胞功能并提高其生育能力 本研究發現,對高脂飲食小鼠喂食雷公藤紅素后體質量沒有顯著降低,這與既往研究結果不同。近年來,雷公藤紅素已經在多項研究中被發現可以作為瘦素增敏劑來有效降低肥胖小鼠的體質量[21]。本研究與既往研究的這種差異可能來自于喂食雷公藤紅素的持續時間不同。既往研究中,小鼠需要喂食2個月,大鼠喂食6周后,體質量方能顯著降低[22-23],而本研究僅喂食21 d。盡管體質量沒有下降,但是本研究發現雷公藤紅素可以通過作用于卵巢局部,降低顆粒細胞炎癥因子基因的表達,提高顆粒細胞激素合成限速酶編碼基因表達水平,改善顆粒細胞的雌激素分泌功能。本研究還發現,雷公藤紅素組小鼠顆粒細胞中Bax、Bak1和Casp8的表達被顯著抑制,而Fadd、Traf2和Casp9的表達仍然顯著增高,提示雷公藤紅素通過抑制線粒體凋亡途徑,而不是死亡受體凋亡途徑來改善顆粒細胞的功能。這可能與雷公藤紅素具有較強的抗氧化應激能力有關,有研究證明雷公藤紅素可以對線粒體發揮作用[24],而線粒體是炎癥反應發生的重要細胞器之一。與雷公藤紅素具有相似功能的二甲雙胍和白藜蘆醇,已經被證明可以通過保護顆粒細胞線粒體功能,抑制炎癥反應,改善多囊卵巢綜合征女性的生育力[25],這也為本研究發現的雷公藤紅素可以通過抑制細胞凋亡和炎癥,改善肥胖小鼠生育力提供了佐證。
綜上所述,本研究初步證實肥胖可以誘導雌性小鼠卵巢局部的顆粒細胞炎癥因子基因表達異常,誘發顆粒細胞凋亡,降低顆粒細胞的雌激素分泌功能,進而導致生育能力降低。雷公藤紅素在不降低體質量的情況下,即可通過抑制卵巢局部顆粒細胞的炎癥反應和線粒體凋亡途徑的激活,上調顆粒細胞激素合成限速酶編碼基因表達水平,改善顆粒細胞的雌激素分泌功能,改善肥胖小鼠模型的生育能力。由于炎癥反應和細胞凋亡是通過多種細胞通路進行調控,雷公藤紅素改善顆粒細胞激素分泌功能的具體分子機制仍然有待進一步探索。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
學苑創造·A版(2020年9期)2020-10-13 09:41:02
中國生殖健康(2019年2期)2019-08-23 08:12:08
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
產品可靠性報告(2017年7期)2017-09-05 09:49:12
汽車觀察(2016年3期)2016-02-28 13:16:26
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
中國質量與標準導報(2014年1期)2014-02-28 22:21:28
