中國瀕危紅樹植物研究與恢復現狀
2021-03-02 07:03:46陳光程鐘才榮
應用海洋學學報 2021年1期
張 穎,陳光程,鐘才榮
(1.福建省海洋生態保護與修復重點實驗室,福建 廈門 361005; 2.嶺南師范學院生命科學與技術學院,廣東 湛江 524048;3.海南省林業科學研究院、海南省紅樹林研究院,海南 海口 571129)
紅樹林是熱帶、亞熱帶海灘上特有的綠色植物群落,是陸地向海洋過渡的一種獨特森林生態系統。作為海岸濕地生態系統中唯一的木本植物,紅樹林起到了海岸森林的作用,在防風消浪[1]、促淤護岸、凈化生態環境[2]、維持生物多樣性[3]等方面具有重要意義。在世界范圍內,紅樹林創造著巨大的生態服務價值[4]。中國紅樹林面積僅占世界紅樹林面積的 0.14%,但紅樹植物種類達到全世界紅樹植物種類的約 1/3,在紅樹林植物種類多樣性保護中占據特殊的地位[5]。在人類活動和全球氣候變化的雙重影響下,紅樹林生態系統正面臨著嚴峻的生物多樣性喪失問題[6],中國37種原生紅樹植物和半紅樹植物中有近一半的種類處于不同程度的珍稀瀕危狀態[7]。相較之下,在世界范圍內的70種真紅樹植物中,有11種(16%)紅樹植物被列入極度瀕危、嚴重瀕危和易危狀態[3,8]。本文綜合國際自然及自然資源保護聯盟(the International Union for Conservation of Nature and Natural Resources, IUCN)物種紅色名錄瀕危等級和標準(3.1版)[9]、《中國珍稀瀕危保護植物名錄》[10]、《中國植物紅皮書(稀有瀕危植物)》[11]、《國家重點保護野生植物名錄(第一批)》[12]、《中國物種紅色名錄》[13]、《中國生物多樣性紅色名錄(高等植物卷)》[14]、各省瀕危植物保護名錄和圖鑒,以及瀕危紅樹植物的相關研究報導等文獻資料,全面梳理了我國瀕危紅樹植物的瀕危現狀、保護性研究以及物種恢復現狀,以期為我國瀕危紅樹植物的科學保護和恢復提供支撐。
1 我國瀕危紅樹植物的瀕危現狀
根據IUCN的標準[9],我國20種瀕危紅樹植物中,屬于極危(Critically Endangered, CR)等級的有:紅欖李(Lumnitzeralittorea)、海南海桑(Sonneratia×hainanensis)、卵葉海桑(S.ovata)和拉氏紅樹(Rhizophora×lamarckii)4種[5,7,15-19];屬于瀕危(Endangered, EN)等級的有:擬海桑(S.×gulngai)、小花老鼠簕(Acanthusebracteatus)、尖葉鹵蕨(Acrostichumspeciosum)和海濱貓尾木(Dolichandronespathacea)4種[5,15,17];海桑(S.caseolaris)和海蓮(Bruguierasexangula)屬于近危(Near Threatened, NT)等級[5];屬于易危(Vulnerable Endangered,VU)等級的有:瓶花木(Scyphiphorahydrophyllacea)、正紅樹(R.apiculata)、尖瓣海蓮(B.sexangulavar.Rhynchopetala)、蓮葉桐(Hernandianymphaeifolia)、水芫花(Pemphisacidula)、木果楝(Xylocarpusgranatum)、水椰(Nypafruticans)、銀葉樹(Heritieralittoralis)、鈍葉臭黃荊(Premnaobtusifolia)和玉蕊(Barringtoniaracemosa)等10種[5,17,20-22](表1)。但從世界紅樹植物瀕危等級來看,以上大多數瀕危紅樹植物屬于無危(Least Concern, LC)等級,除卵葉海桑屬于近危等級,而海南海桑、擬海桑、海濱貓尾木、蓮葉桐、水芫花、木果楝、鈍葉臭黃荊和玉蕊未進行瀕危定級。但IUCN瀕危物種紅色植物名錄中海南海桑為極危等級[9]。瀕危紅樹植物分布最集中的省份是我國的海南,其瀕危物種所占比例高達50%,同時海南也是我國紅樹植物物種最豐富的省,我國分布的37種[7]紅樹植物在海南均有自然分布,可見海南紅樹林的保護對瀕危紅樹植物資源挽救具有重要意義。其次是廣東和廣西。
我國紅樹植物中,瀕危種數量最多的屬為海桑科的海桑屬(Sonneratia),現有6種海桑屬紅樹植物[15-16]中,海南海桑、卵葉海桑和擬海桑處于極度瀕危狀態。從世界范圍來看,瀕危物種所占比例最高的屬是銀葉樹屬(Heritiera),占據了3個瀕危物種中的2種,分別為小葉銀葉樹(H.fomes)和卵葉銀葉樹(H.globosa)[3],而另一個瀕危物種是海桑屬的格氏海桑(S.griffithii),該物種分布于馬來西亞和印度等亞洲沿岸和西太平洋群島地區[8,17],表現出數量少、部分原生地滅絕和種子活力低[3]的特點。海南海桑的研究報導始于1985年我國學者高蘊璋的研究[27]。除在1996年,Muller 等對婆羅洲(Borneo)海桑屬的一個雜交類群S.alba×S.ovata進行過形態學和細胞學研究的相關報導外(未正式命名),海南海桑在我國以外未見自然分布報導[28]。因此,海南海桑被部分學者稱為我國海南特有紅樹植物[29-30],但由于國外存在其親本卵葉海桑和杯萼海桑的交叉分布現象,本研究認為海南海桑不是我國特有種。使君子科欖李屬(Lumnitzera)瀕危紅樹植物紅欖李為我國國家二級重點保護植物,其野生種目前只有唯一的分布地(三亞鐵爐港),同時野生種生存數量僅9株[31],屬我國瀕危紅樹植物中瀕危狀態最為嚴峻的真紅樹植物。其被列入《中國物種紅色名錄》[13]、《中國植物紅皮書》[11]和《中國生物多樣性保護行動計劃》[23]。但從世界范圍來講,紅欖李的分布范圍卻較為廣泛,包括馬來西亞、泰國、印度尼西亞、斯里蘭卡、澳大利亞以及我國海南。除我國外,紅欖李在世界其他分布地均未見瀕危報道,開花結實狀況正常,具備自然更新能力[24,32-34],其瀕危等級被列為無危等級。水椰亞科是棕櫚科最古老與特殊化的一個類群,瀕危紅樹植物水椰十分珍稀,1984年被列為第一批國家三級保護植物[15,17],同時也被列入《中國植物紅皮書》[11]、《中國生物多樣性保護行動計劃》[23]和《海南省省級重點保護野生植物名錄》[25]。水椰在我國僅天然分布于海南島的海口、文昌、瓊海、萬寧和三亞等地,已有文獻證明三亞分布的群落已被破壞[15]。根據其現有野生種數量及生長狀態,水椰在我國處于易危狀態,但在全球范圍內水椰的天然分布范圍廣泛,從亞洲東部(琉球群島)、南部(斯里蘭卡、印度的恒河三角洲和馬來西亞)至澳大利亞、所羅門群島等地[3,8,16],屬于無危等級的物種[3]。
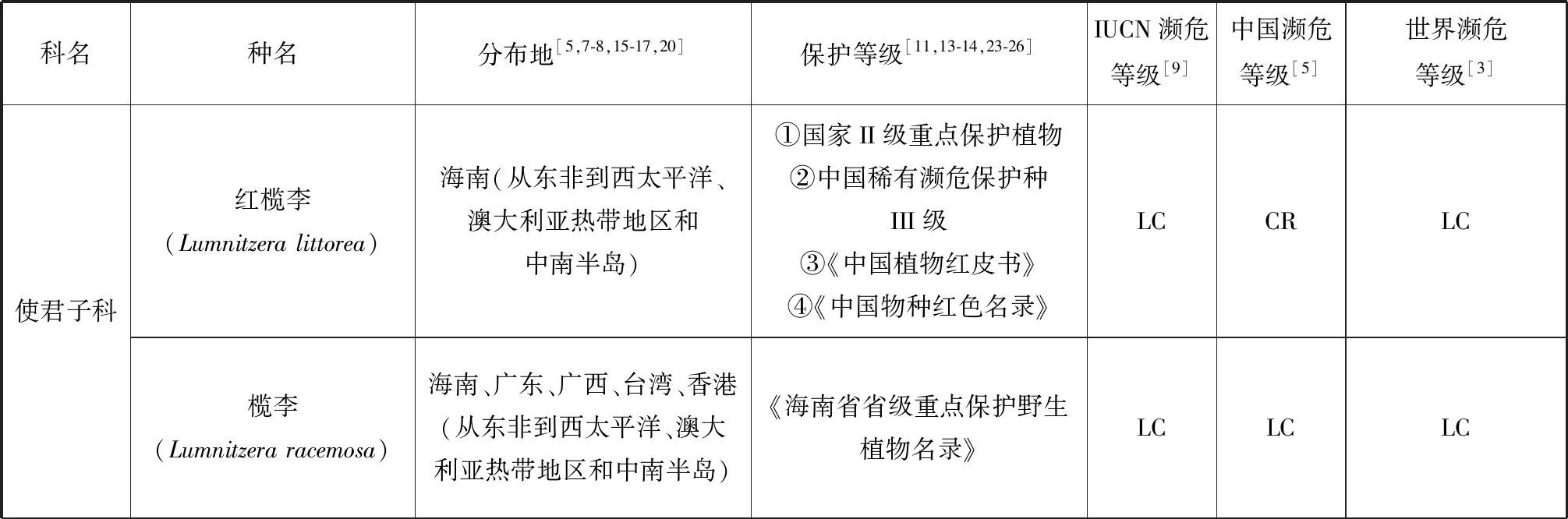
表1 瀕危紅樹植物分布現狀及保護等級
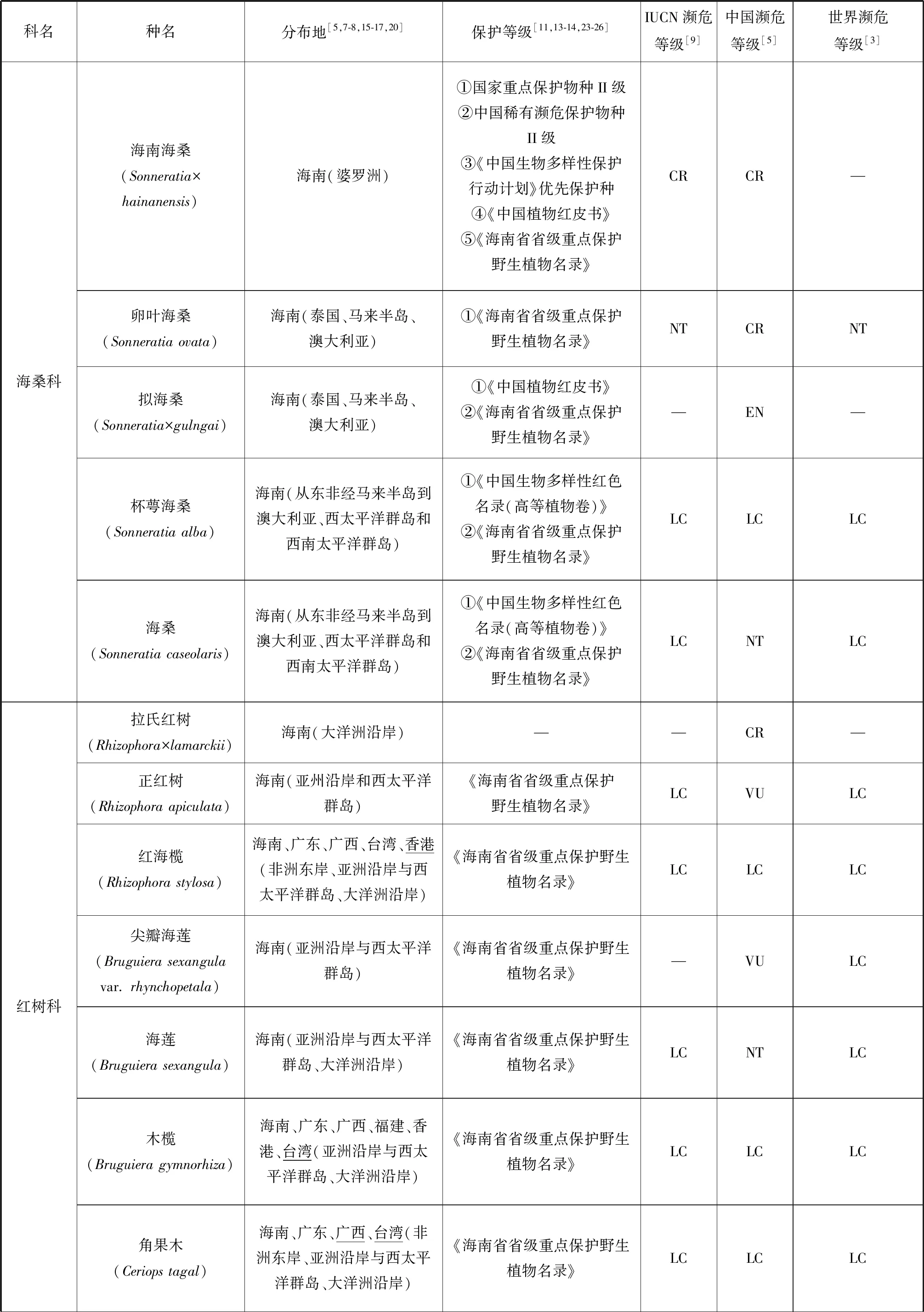
續表1
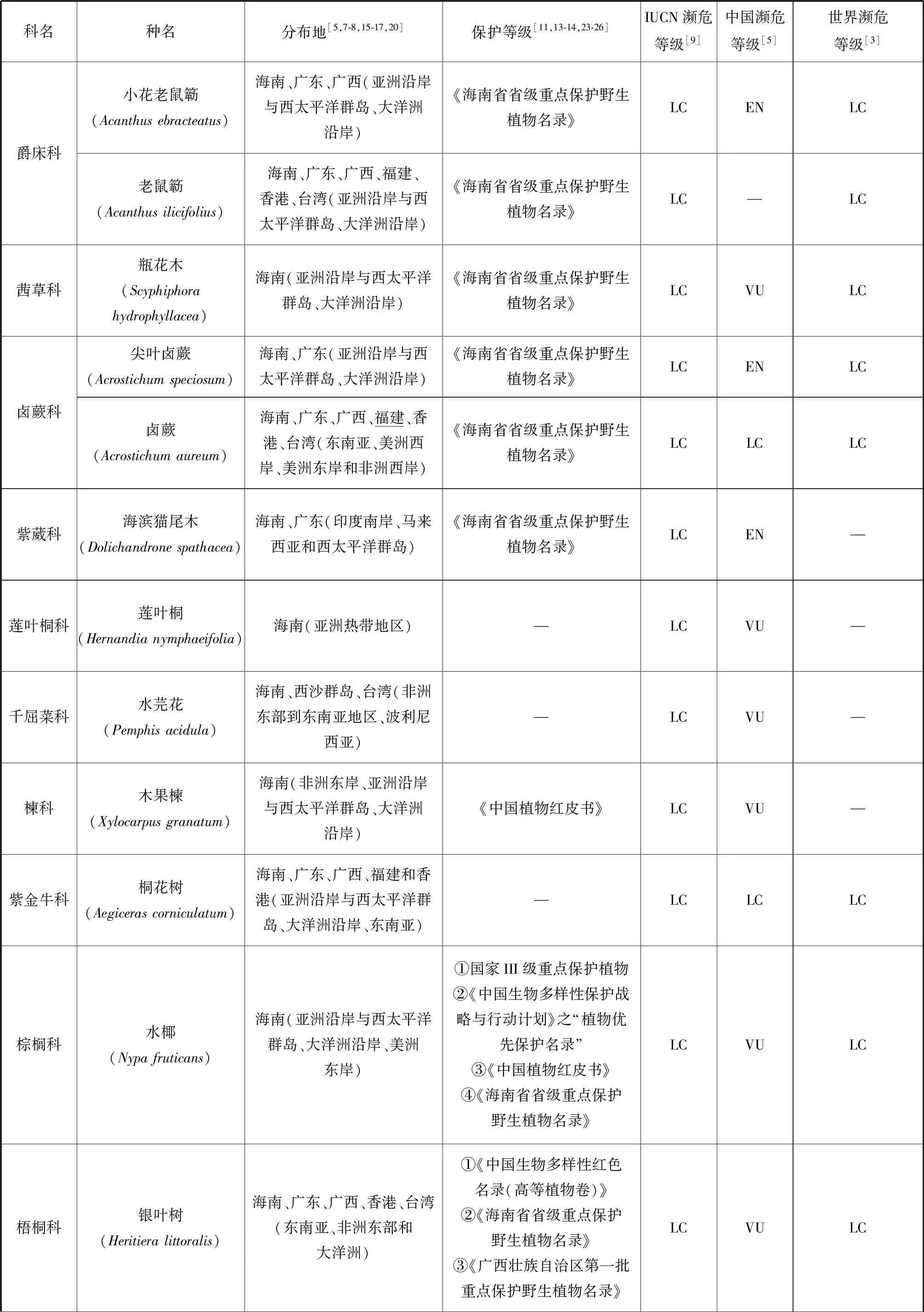
續表1
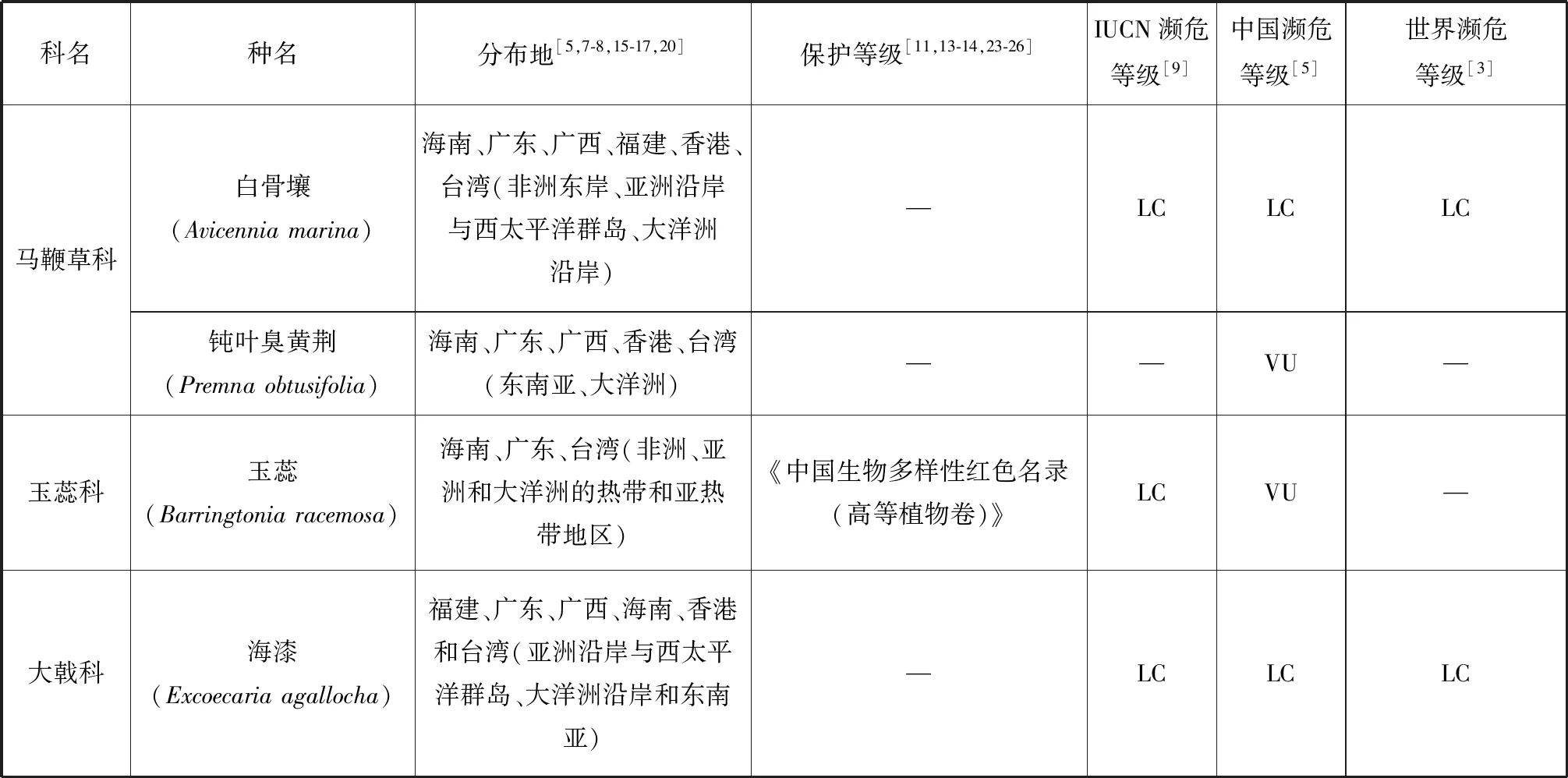
續表1
2 瀕危紅樹植物致危原因
瀕危紅樹植物的保護性研究主要集中在瀕危種的分布地和現有分布數量[5,9-10,16,35-36]的調查上。對于瀕危機制和致危因素的研究則主要集中在少數幾種瀕危紅樹植物上,如:紅欖李[24,32-34]、蓮葉桐[37]、海南海桑[38-41]、卵葉海桑[42]、擬海桑[40]、水椰[21]和水芫花[22]。瀕危紅樹植物的致危因素包括內因和外因兩個方面。
內因主要有:①相對較低的遺傳多樣性和較低的基因流,種群遺傳分化較小。屬于此類的瀕危紅樹植物有紅欖李[43]、海南海桑[36]、擬海桑[4,27]和卵葉海桑[39]等。遺傳多樣性水平決定了物種對環境的適應能力,由于個體數量有限、分布區域狹窄,這些瀕危紅樹植物表現出近交衰退和遺傳漂變作用,從而導致后代種群的遺傳多樣性降低,但同時降低的遺傳多樣性也會使瀕危植物對環境選擇的壓力更為敏感,從而增大了物種滅絕的風險[44]。②分布地狹窄,種群數量少且破碎化,野生種個體數量少,為小種群植物。屬于此類的瀕危紅樹植物有紅欖李[23,31-33]、海南海桑[40]、卵葉海桑[27]和擬海桑[4,36]。還有一些瀕危紅樹植物近年來出現部分自然棲息地滅絕現象[7,15,20],如海南三亞的尖葉鹵蕨和小花老鼠簕,廣西、臺灣和香港分布的角果木,福建的海漆和鹵蕨,香港的紅海欖[7],以及廣東的海濱貓尾木等,進一步加劇了瀕危植物種群的破碎化和瀕危程度。③存在不同程度的有性生殖障礙。一類為種子量大,但空胚率高,如:紅欖李[32,35]和海濱貓尾木,其中紅欖李有胚種子的萌發率亦很低(<15%)。紅欖李由于野生數量不足10株,雖然花期可長達半年以上,但不同植株花期分散;自然條件下的花粉生活力不超過10%[33]。同時紅欖李在花蕾期易受到害蟲的侵害,2013年監測到近58%的果實受害而空胚[45]。自然條件下,紅欖李的傳粉媒介易受到同期開花的伴生紅樹植物的競爭,降低了傳粉效率。由于異交授粉途徑受阻而產生被迫自交現象,自交衰退也是造成空胚的主要原因之一[24]。海濱貓尾木每莢果種子數量超過400粒,其種子空胚率高的原因至今未見報導。另一類為種子產量少或極少,種子萌發率較高但不易成苗,如:海南海桑[40]、擬海桑和瓶花木等。海南海桑花粉平均敗育率為54.34%,落果率高達70%~80%[28];海南海桑大部分種子有自然退化跡象,果粒和種子質量比擬海桑好,但遠差于其他海桑科植物[42]。海南海桑和擬海桑由于母樹數量少,果實空粒率高,加大了種子收集的難度。海南海桑種子空殼粒高達53.33%,但適宜環境條件下種子萌發率可超過80%[33]。自然狀態下,海南海桑和擬海桑母樹周圍未見幼樹存在,可見人工輔育是保護瀕危紅樹植物的必要手段。
外因主要表現在:①由于養殖業、旅游業或房地產業的不當開發和利用造成棲息地被破壞,受此影響的瀕危紅樹植物[5,7,17,20,31,35,37]有:紅欖李、卵葉海桑、木果楝、三亞的尖葉鹵蕨[15]、文昌的水芫花、銀葉樹、玉蕊和鈍葉臭黃荊等。瀕危紅樹植物的瀕危現狀調查不夠細致,瀕危物種的生態價值和意義被低估,造成了瀕危物種分布地的保護工作不到位。②盜伐也是加速紅欖李、老鼠簕、水椰、蓮葉桐、玉蕊、水芫花和木果楝等瀕危植物瀕危狀態的重要因素[5,7,15,17]。
3 瀕危紅樹植物的種群恢復現狀
3.1 瀕危紅樹植物育苗技術
瀕危紅樹植物對種子繁育和苗木培育技術的研究僅局限于紅欖李[34]、蓮葉桐[46]、海南海桑[38]、卵葉海桑[42]、擬海桑[47]、海濱貓尾木[48]、水椰[49]和木果楝[50]等少數物種。瀕危紅樹植物的育苗技術涵蓋了種子收集、種子處理、種子萌發、煉苗和野外適應等步驟。瀕危紅樹植物大多存在不同程度的有性生殖障礙,為得到有胚種子,人工輔助授粉是較為有效的手段,通過對紅欖李人工輔助授粉,紅欖李的種子萌發率由0.2‰提高至5.6‰[37]。對結果實率較低的瀕危紅樹植物,要在果實成熟期提前掛網套袋,減少果實落地后被螃蟹啃實或被潮汐帶走的損失;及時采收,使種子盡量保持充足的水份[24,38,42]。獲得的種子需要在低鹽濃度的鹽水或稀釋海水中浸泡,輔助以殺菌劑進行殺菌。部分瀕危物種需要較長時間浸泡以去除種皮,如紅欖李[51]和蓮葉桐[52]。為了打破種子的休眠作用,適宜濃度的激素處理能產生較好的效果,如擬海桑[47]和水椰[49]。為提高種子萌發率,育苗基質、溫度、濕度和光照條件[47,49-52],以及播種方式都是重要的影響因素。木果楝種子在播種時需要保持種子胚軸水平,種子一半埋于土中,一半暴露于空氣中,胚芽部分位于土壤和空氣的交界處,才有利于種子扎根生長[50]。在苗期養護過程中,部分瀕危紅樹植物對光照十分敏感,蓮葉桐幼苗在遮光條件下,光照強度為5 000~6 000 lx范圍內能有效提高成活率[51],苗期適當遮陽對海濱貓尾木幼苗的成活也是至關重要的[52]。由于紅樹植物生長在潮間帶的特殊生境,受到高光照、高鹽和生理干旱脅迫,培育出的瀕危種幼苗能否在野外成活,煉苗和野外馴化是必不可少的步驟。煉苗期要加強鹽濃度適應、高光強適應和耐貧瘠適應。而野外適應階段需選擇瀕危物種原生地相近的潮汐環境建立苗圃,同時在前期間斷供給淡水,而后逐漸減少頻次,以便使幼苗能夠逐漸適應自然環境[47,49-53]。
3.2 瀕危紅樹植物的種群恢復現狀
我國瀕危紅樹植物的種群恢復工作相對滯后,僅局限于極少數物種,且恢復的內容僅局限于引種回歸實驗[31]和少數瀕危紅樹植物的引種馴化實驗上[46,54-55](表2)。在擴大種質資源數量的試驗中,半紅樹植物如:楊葉肖槿、銀葉樹、水黃皮和海濱貓尾木(圖1)表現出較好的環境適應能力,生長狀況良好,能夠正常開花、結實,能夠形成種群,但自然更新能力不強,還未見種群擴大。半紅樹植物蓮葉桐與其他瀕危真紅樹植物則表現出對引種地不同程度的不適應。除尖葉鹵蕨、海蓮、尖瓣海蓮、陵水和三亞引種的紅欖李(圖2)、海南引種的正紅樹、木果楝、海南海桑和卵葉海桑能夠正常生長、開花、結果外,其他真紅樹瀕危種在引種地表現出不同程度的生長抑制現象,或未能成活或不能正常繁育。引種的瀕危真紅樹植物中能夠形成種群的只有海蓮、尖瓣海蓮和東寨港國家級紅樹林保護區引種的木果楝、海南海桑和卵葉海桑。除了文昌清瀾港的卵葉海桑外都不能實現自然更新,均未見種群擴大。
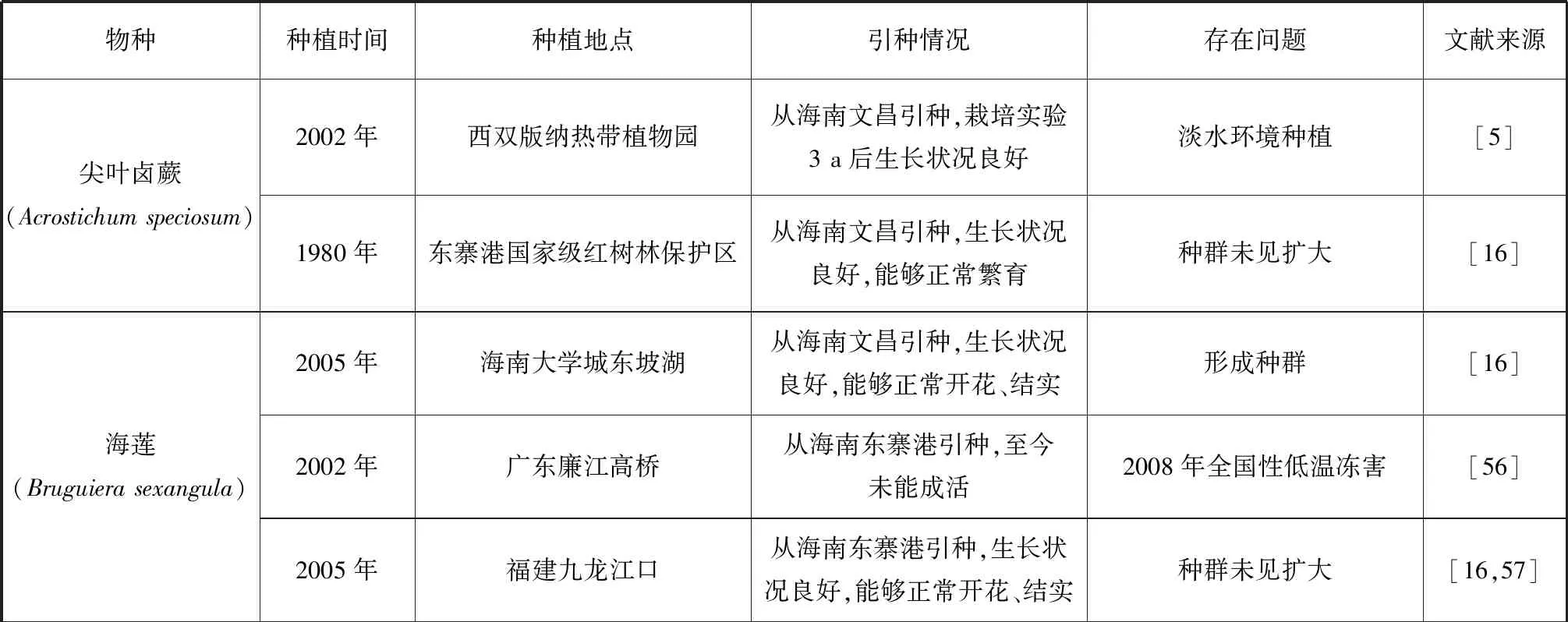
表2 瀕危紅樹植物的種群恢復
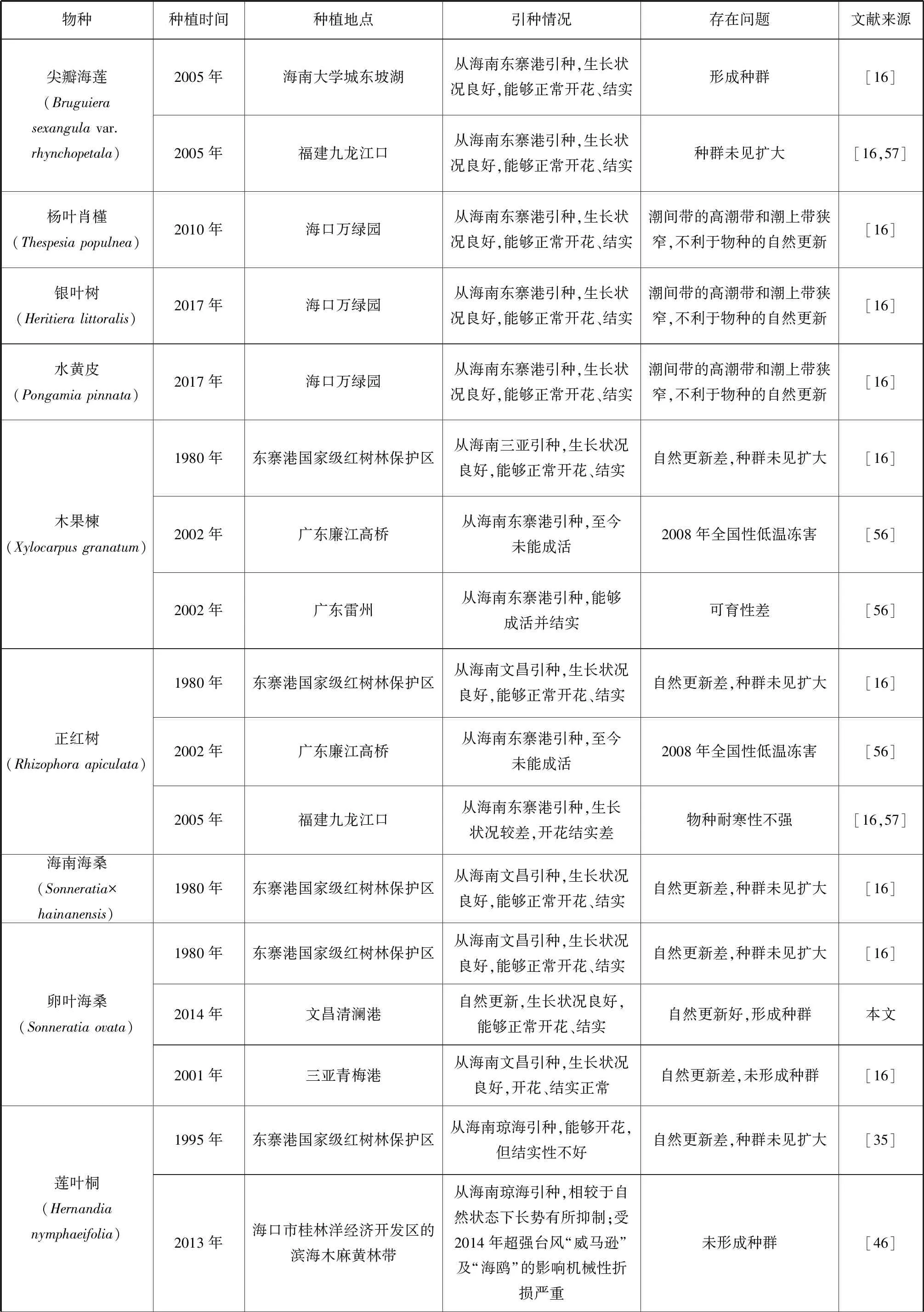
續表2
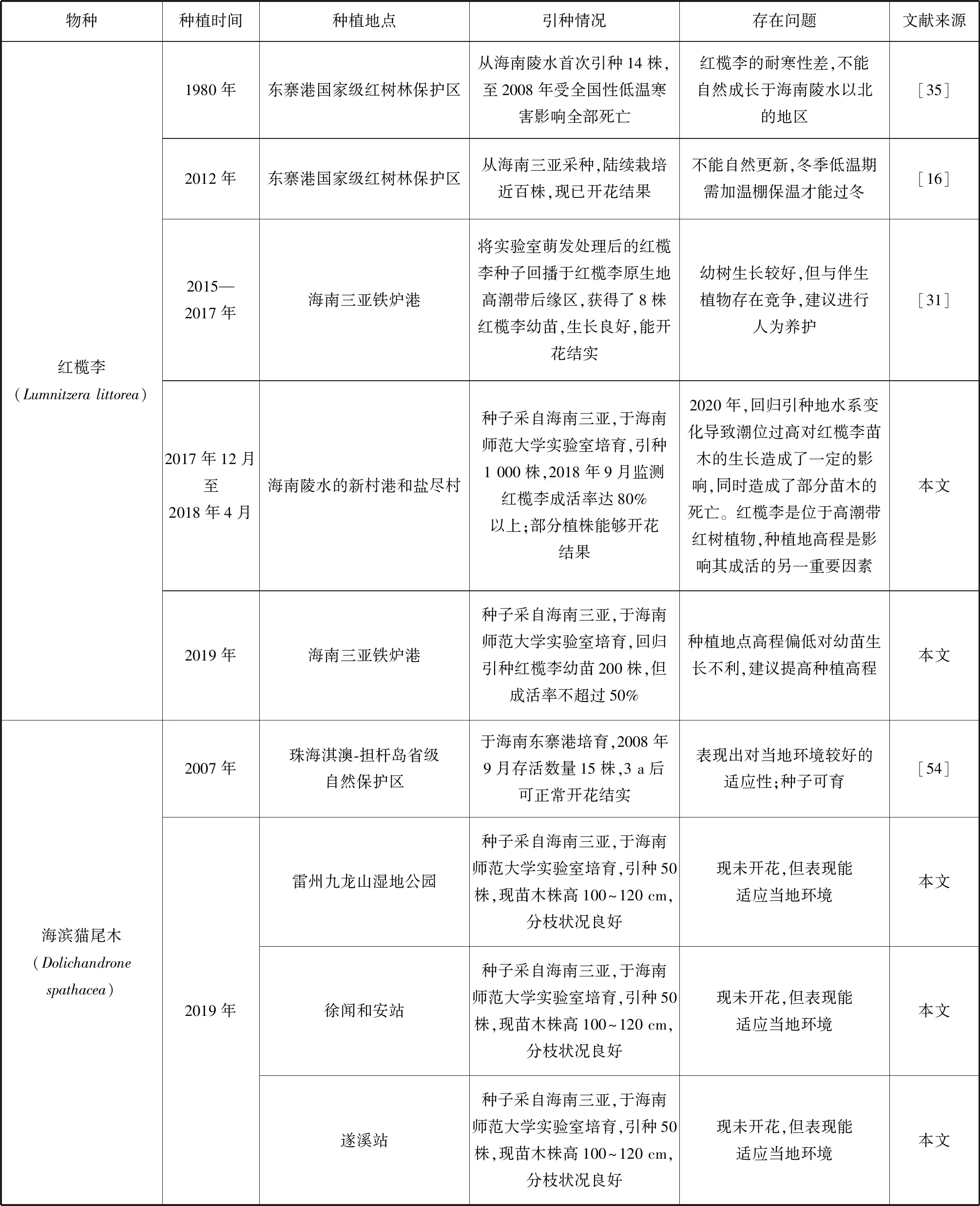
續表2
現有引種試驗表明對瀕危紅樹植物宜林地的選擇缺少科學依據,瀕危紅樹植物原生地生長因子本底調查數據不足,在選擇引種地時未能充分考慮植物對生境條件的要求進行選址,種植后缺少持續監測和管護,使為數稀缺的瀕危紅樹植物種源在引種地上無法成活,造成了巨大的資源浪費。原生地的引種回歸工作需要重視,瀕危紅樹植物對原生地能表現出較好的適應能力,紅欖李種子回播于紅欖李自然分布區(海南三亞鐵爐港)高潮帶后緣地,現已獲得了8株紅欖李幼苗,其中一株高達2 m,觀測到開花結實。文昌清瀾港一廢棄漁塘,面積約為3 333 m2,無人為干擾,5 a間新增的卵葉海桑均為自然更新,同時新增的伴生物種還有杯萼海桑、海桑、欖李、桐花樹和鹵蕨等紅樹植物。建議對該恢復地進行科學跟蹤監測和重點保護。

圖1 瀕危紅樹植物海濱貓尾木引種回歸實驗Fig. 1 Reintroduction experiment of endangered mangrove D.spathacea

圖2 瀕危紅樹植物紅欖李引種回歸實驗Fig. 2 Reintroduction experiment of endangered mangrove L. littorea
4 瀕危紅樹植物的種群恢復建議
在全球紅樹林持續減少的態勢下,中國紅樹林面積增至2.9萬公頃,20 a來中國紅樹林面積增加7 000公頃,得益于紅樹林資源保護修復力度的持續加大[58]。據1980—2010年的研究成果表明,世界范圍內的紅樹林生態系統出現同質性的趨勢。紅樹林保護的重心偏重于紅樹林分布地的保護,對瀕危紅樹植物單個物種的保護工作不足[59]。在我國,瀕危紅樹植物的恢復目前是由保護區來主導的,主要的技術手段是由保護區引種和移植。一方面能夠實現對瀕危樹種的精心管護,另一方面使保護區物種數量增加,種類差異變小,生物多樣性豐度持平或增加[60]。但瀕危紅樹植物的種群恢復僅靠保護區的力量是遠遠不夠的。物種瀕危機制的研究需要大量科研力量和科研經費的投入,要突破育種難題才能從根本上解決瀕危紅樹植物的種源問題。因此鼓勵各級紅樹林保護區聯合科研單位共同開展瀕危紅樹植物種質資源的保護和進行種質資源的人工培育工作,建立適當規模的濱海珍稀植物苗圃,以保障紅樹林生態修復中所需瀕危紅樹植物苗木的供給。
綜合瀕危紅樹植物形成的內外因素,重點開展珍稀瀕危紅樹植物小種群生境地的保護,同時科學選擇宜林地。引種苗木要持續跟蹤監測環境因子,及時調控,減小致瀕危因子的影響。要在“培育得出苗、種得下去”的基礎上,“養護得當、監測得好”,從而最終實現瀕危紅樹植物自然更新能力的恢復。
猜你喜歡
歌海(2024年6期)2024-03-18 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
兒童故事畫報·自然探秘(2022年3期)2022-04-27 00:17:10
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
瘋狂英語·初中天地(2018年6期)2018-11-24 02:39:24
汽車觀察(2018年10期)2018-11-06 07:05:26
歌海(2018年4期)2018-05-14 12:46:15
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
