不同性別和交配狀態(tài)的馬鈴薯塊莖蛾對馬鈴薯揮發(fā)物的觸角電位反應
2021-03-09 00:58:58李祥張小嬌肖春董文霞
中國農(nóng)業(yè)科學 2021年3期
李祥,張小嬌,肖春,董文霞
不同性別和交配狀態(tài)的馬鈴薯塊莖蛾對馬鈴薯揮發(fā)物的觸角電位反應
李祥,張小嬌,肖春,董文霞
云南農(nóng)業(yè)大學植物保護學院/云南生物資源保護與利用國家重點實驗室,昆明 650201
【】探明不同性別和交配狀態(tài)的馬鈴薯塊莖蛾()對寄主植物(馬鈴薯)揮發(fā)物的電生理反應的差異,為利用化學生態(tài)手段防治馬鈴薯塊莖蛾提供參考。利用觸角電位技術(shù)(electroantennogram,EAG),測定未交配雌蛾、交配雌蛾、未交配雄蛾3種不同生理狀態(tài)的馬鈴薯塊莖蛾對馬鈴薯塊莖和植株釋放的16種揮發(fā)性化合物(包括6種脂肪族化合物:1-辛烯-3-醇、丁醇、庚醛、辛醛、壬醛、癸醛;4種芳香族化合物:苯乙醇、苯乙醛、苯乙酮、水楊酸甲酯;6種萜類化合物:-蒎烯、對-聚傘花素、反--羅勒烯、桉葉油醇、反--法尼烯、茉莉酸甲酯)的電生理反應及對其中6種化合物(1-辛烯-3-醇、庚醛、壬醛、苯乙酮、水楊酸甲酯、反--羅勒烯)的劑量反應。馬鈴薯塊莖蛾對這16種化合物均能產(chǎn)生觸角電位反應,其中未交配雌蛾和雄蛾對脂肪族醇類化合物的EAG反應值較高,交配雌蛾對脂肪族醛類化合物的EAG反應值較高,3種不同生理狀態(tài)的馬鈴薯塊莖蛾對萜類化合物的EAG反應值均較低。當刺激劑量為1 000 μg時,不同性別和交配狀態(tài)的馬鈴薯塊莖蛾對同種化合物的反應差異較大,未交配雌蛾對辛醛、水楊酸甲酯、反--羅勒烯的反應強于交配雌蛾和未交配雄蛾,交配雌蛾對庚醛、壬醛、-蒎烯的反應強于未交配雌蛾和雄蛾,未交配雄蛾對1-辛烯-3醇、苯乙醇、苯乙醛的反應強于雌蛾。在測試劑量范圍內(nèi)(1、10、100、1 000 μg),3種生理狀態(tài)的馬鈴薯塊莖蛾對6種化合物的EAG反應值隨刺激劑量的增加呈現(xiàn)增大的趨勢,均未達到飽和閾值,交配雌蛾對同一化合物不同劑量的反應差異更為明顯。馬鈴薯塊莖蛾未交配雌蛾和雄蛾對脂肪族醇類化合物較敏感,交配雌蛾對脂肪族醛類化合物較敏感,雌、雄蛾對萜類化合物均不敏感;對于單個化合物的電生理反應,交配雌蛾對庚醛、壬醛、-蒎烯更為敏感;對同一化合物不同劑量的電生理反應,交配雌蛾同樣更為敏感。
馬鈴薯塊莖蛾;馬鈴薯揮發(fā)物;觸角電位反應
0 引言
【研究意義】馬鈴薯塊莖蛾()是危害馬鈴薯和煙草等茄科植物最嚴重的害蟲之一,目前在亞洲、歐洲、美洲、非洲、大洋洲的100多個國家均有分布[1-2]。在中國,隨著馬鈴薯種植面積的逐步擴大,馬鈴薯塊莖蛾現(xiàn)已廣泛分布于云南、貴州、四川、廣西、湖南等多個省區(qū),且危害日益加重,其中以云南、貴州、四川三省受害最為嚴重,馬鈴薯塊莖蛾在大田時期造成的危害可使馬鈴薯減產(chǎn)30%,對倉儲期馬鈴薯塊莖的危害高達100%[3-4]。由于馬鈴薯塊莖蛾幼蟲具有鉆蛀危害的習性,因此防治困難[1]。近幾年,一些研究者試圖通過利用植物揮發(fā)物組分調(diào)控其行為,尋找防治馬鈴薯塊莖蛾的新途徑[5-11]。因此,研究不同性別和交配狀態(tài)的馬鈴薯塊莖蛾對植物揮發(fā)物敏感性的差異,對于馬鈴薯塊莖蛾行為調(diào)節(jié)劑(包括引誘劑和驅(qū)避劑)的精準開發(fā)和應用具有重要意義。【前人研究進展】同種昆蟲由于性別和生理狀態(tài)的差異,植物揮發(fā)物組分在其行為反應中所起的作用不同,因而對植物揮發(fā)物各組分的敏感性也不同。不同生理狀態(tài)的番石榴實蠅()雌、雄蟲對寄主番石榴氣味具有不同的敏感性和選擇性,已交配雌蟲反應最強,其次是性成熟未交配的雄蟲,性未成熟的雄蟲反應最弱[12]。交配狀態(tài)直接影響蛾類昆蟲對植物揮發(fā)物的行為反應,例如煙草天蛾()雌蟲交配后對寄主植物的反應增強[13],蘋果蠹蛾()交配雌蟲對蘋果揮發(fā)物的反應強于未交配雌蛾[14],只有交配后的白蠟麗細蛾()才對寄主植物(白蠟樹)葉片揮發(fā)物發(fā)生定向反應[15]。蛾類昆蟲的觸角電生理反應也因其性別和交配狀態(tài)而異。羽化后1—3 d未交配的美國白蛾()雌、雄蛾對寄主植物揮發(fā)物的觸角電位(electroantennogram,EAG)反應不同[16],羽化后2—4 d的未交配茶尺蠖()雌、雄蛾對非寄主迷迭香()揮發(fā)物組分的EAG反應也存在差異[17]。冷杉梢斑螟()和海灰翅夜蛾()雌蟲交配后對寄主植物揮發(fā)物的EAG反應增強[18-20]。Das等測定了印度馬鈴薯塊莖蛾羽化后2—3 d的雌、雄成蟲對27種植物揮發(fā)物組分的EAG反應,發(fā)現(xiàn)其對大部分供試化合物的電生理反應因性別而異[21],但有關(guān)雌蛾在交配前后對寄主揮發(fā)物的敏感性差異未見報道。另外,昆蟲對植物揮發(fā)物的敏感性也因地域不同而存在差異,例如粉蝶盤絨繭蜂()荷蘭種群和中國種群對寄主揮發(fā)物的EAG反應存在差異[22]。【本研究切入點】馬鈴薯塊莖蛾的交配狀態(tài)是否影響其對寄主植物揮發(fā)物的敏感性,中國的馬鈴薯塊莖蛾種群對寄主植物揮發(fā)物的電生理反應是否與印度種群存在差異?為了回答這些問題,本文選取馬鈴薯釋放的16種揮發(fā)性化合物,測定馬鈴薯塊莖蛾未交配雌蛾、交配雌蛾、未交配雄蛾對這些化合物的EAG反應。【擬解決的關(guān)鍵問題】通過分析不同生理狀態(tài)的馬鈴薯塊莖蛾對不同化合物及同種化合物不同劑量電生理反應的差異,探明影響馬鈴薯塊莖蛾對植物揮發(fā)物反應的因素,為利用化學生態(tài)手段防治馬鈴薯塊莖蛾提供參考。
1 材料與方法
試驗于2014—2017年在云南生物資源保護與利用國家重點實驗室昆蟲電生理室完成。
1.1 材料
1.1.1 供試昆蟲 馬鈴薯塊莖蛾采自云南農(nóng)業(yè)大學植物病理實驗大棚,室內(nèi)連續(xù)飼養(yǎng)10代以上,幼蟲以馬鈴薯塊莖(合作88)飼養(yǎng),成蟲飼以10.0%的蜂蜜水。室內(nèi)飼養(yǎng)條件:溫度為(27.0±2.0)℃,相對濕度為(70.0±5.0)%,光周期為L﹕D=14﹕10。試驗時選取羽化后3 d的健康已交配雌蛾、未交配雌蛾、未交配雄蛾。其中,已交配雌蛾通過將剛羽化的雌、雄蛾以2﹕3的比例混合飼養(yǎng)獲得,試驗后解剖雌蛾進一步確認其交配狀態(tài),如果交配囊內(nèi)有精苞,則確定是已交配雌蛾;如果無精苞,則剔除相關(guān)數(shù)據(jù)。
1.1.2 氣味化合物 測定化合物包括6種脂肪族化合物:1-辛烯-3-醇(98.0%,J&K)、丁醇(99.5%,J&K)、庚醛(95.0%,TCI)、辛醛(98.0%,TCI)、壬醛(95.0%,TCI)、癸醛(95.0%,TCI);4種芳香族化合物:苯乙醇(99.0%,Acr?s)、苯乙醛(98.0%,Acr?s)、苯乙酮(98.0%,J&K)、水楊酸甲酯(99.0%,J&K);6種萜類化合物:-蒎烯(98.0%,Sigma- Aldrich)、對-聚傘花素(96.0%,TCI)、反--羅勒烯(90.0%,Sigma-Aldrich)、桉葉油醇(99.0%,Acr?s)、反--法尼烯(98.0%,Sigma-Aldrich)、茉莉酸甲酯(95.0%,Sigma-Aldrich)。其中,馬鈴薯植株揮發(fā)物組分有庚醛、辛醛、水楊酸甲酯、桉葉油醇,馬鈴薯塊莖揮發(fā)物組分有茉莉酸甲酯、1-辛烯-3-醇、對-聚傘花素、丁醇、苯乙酮,植株和塊莖中均存在的揮發(fā)物組分有壬醛、癸醛、苯乙醛、苯乙醇、-蒎烯、反--羅勒烯、反--法尼烯[5,23-25]。以正己烷(色譜純,Merck)作為溶劑,將上述樣品配成5×10-2g·ml-1的溶液,并將1-辛烯-3-醇、庚醛、壬醛、苯乙酮、水楊酸甲酯、反--羅勒烯稀釋成濃度為5×10-3、5×10-4、5×10-5g·ml-1的正己烷溶液,存于-20℃的冰箱待用。
1.2 方法
1.2.1 EAG的測定 EAG反應測定方法和EAG反應相對值的計算方法同文獻[11]。以反-2-己烯醛為參照化合物,其濃度為5×10-2g·ml-1。每個樣品重復4根觸角,每根觸角刺激5次以上,樣品測試前后用參照化合物各刺激一次。
1.2.2 數(shù)據(jù)處理 數(shù)據(jù)采用Excel和SPSS17.0軟件進行分析,利用Origin2017繪圖軟件繪圖。利用-檢驗(<0.05)分析處理與對照之間的差異顯著性。不同處理之間的差異顯著性分析采用單因素方差分析,利用Tukey檢驗分析其差異顯著性(<0.05)。
2 結(jié)果
2.1 馬鈴薯塊莖蛾對1 000 μg的16種揮發(fā)性化合物EAG反應
馬鈴薯塊莖蛾未交配雌蟲、交配雌蟲、未交配雄蟲對1 000 μg的參照化合物反-2-己烯醛的EAG反應值分別為(0.149±0.003)、(0.150±0.006)、(0.152±0.008)mV,三者之間無顯著差異(>0.05)。
當刺激劑量為1 000 μg時,馬鈴薯塊莖蛾未交配雌蛾、交配雌蛾、未交配雄蛾對16種化合物EAG反應絕對值均顯著高于對照(<0.05)。但是,這3種生理狀態(tài)的馬鈴薯塊莖蛾對這16種化合物的EAG相對反應值差異較大。未交配雌蛾和雄蛾對醇類化合物的EAG相對反應值較高,交配雌蛾對醛類化合物的EAG相對反應值較高,3種不同生理狀態(tài)的馬鈴薯塊莖蛾對萜類化合物的EAG相對反應值均較低。未交配雌蛾和雄蛾對1-辛烯-3-醇的反應值顯著高于多數(shù)化合物,分別為152.47%、226.42%;交配雌蛾對壬醛和庚醛的反應值顯著高于其他化合物,分別為191.67%、188.87%(圖1)。
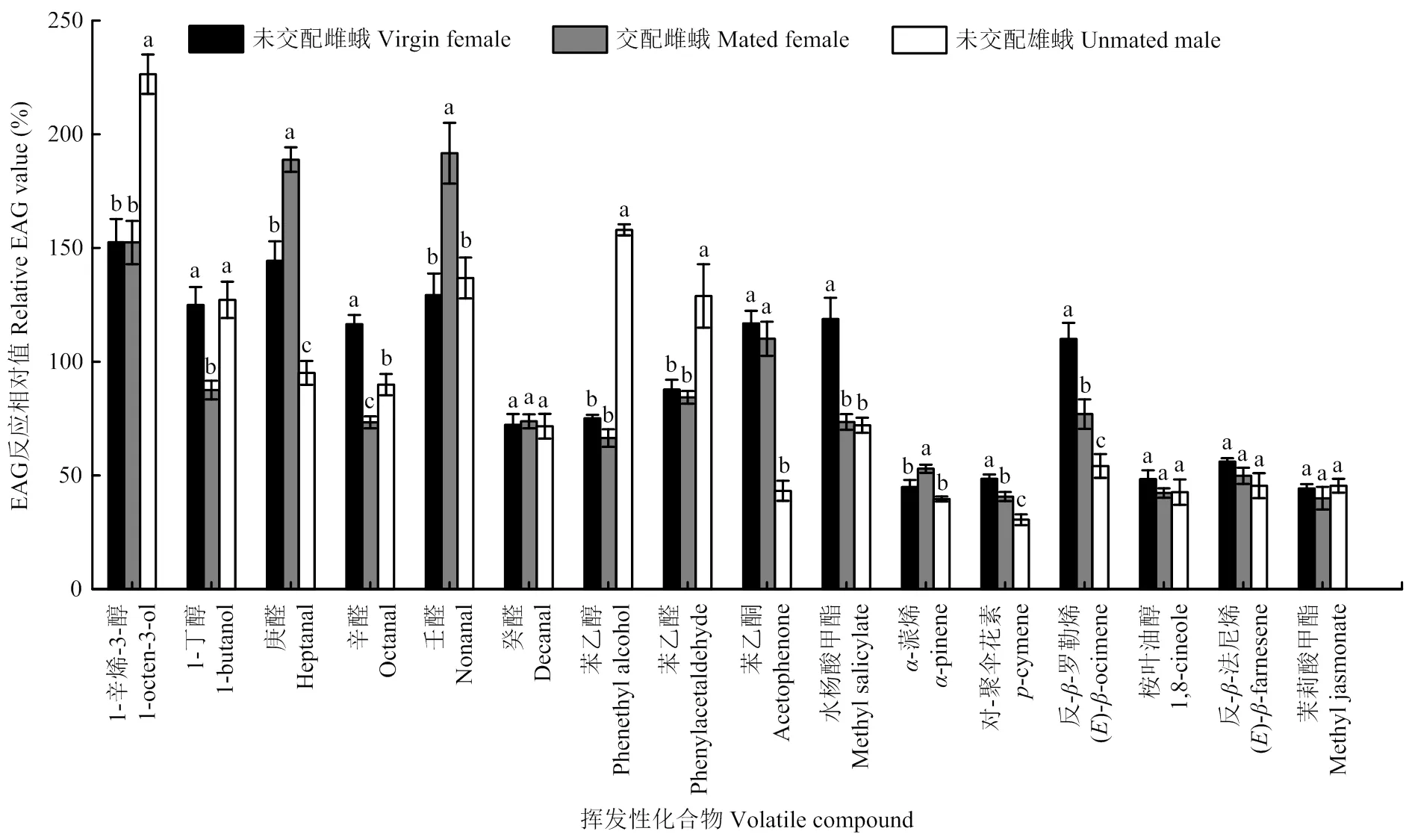
當刺激劑量為1 000 μg時,不同性別和交配狀態(tài)的馬鈴薯塊莖蛾同種化合物的反應差異較大。對庚醛的反應為交配雌蟲>未交配雌蟲>未交配雄蟲,對辛醛的反應為未交配雌蟲>未交配雄蟲>交配雌蟲,對反--羅勒烯和對-聚傘花素的EAG反應均為未交配雌蟲>交配雌蟲>未交配雄蟲。未交配雌蛾對水楊酸甲酯的反應值顯著高于交配雌蛾和未交配雄蛾;交配雌蛾對壬醛、-蒎烯的EAG反應值均顯著高于未交配雌蛾和雄蛾,對丁醇的反應值顯著低于未交配雌蛾和雄蛾;未交配雄蛾對1-辛烯-3-醇、苯乙醇、苯乙醛的反應值顯著高于雌蛾,對于苯乙酮的反應值顯著低于雌蛾(<0.05)(圖2)。
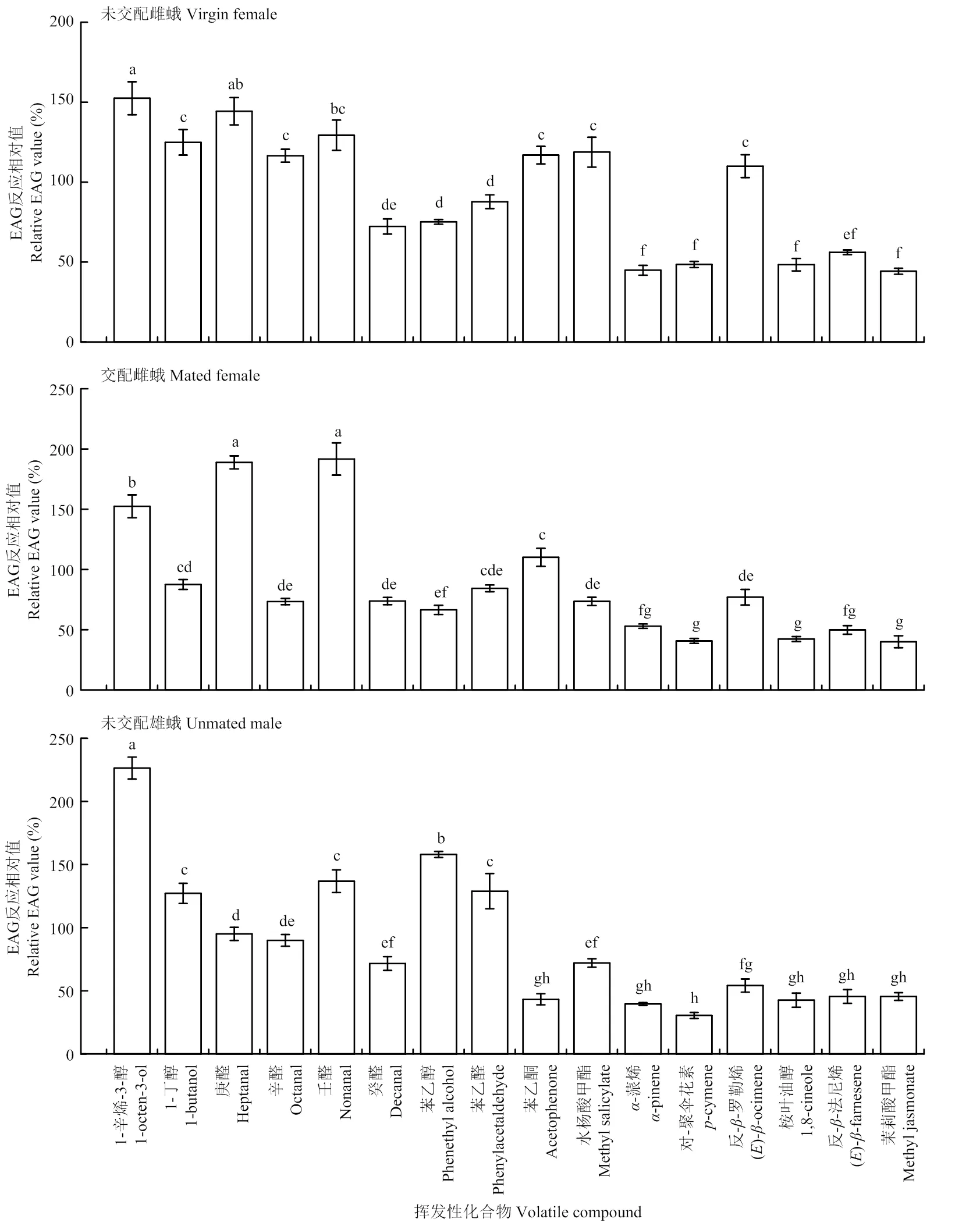
不同小寫字母表示同一生理狀態(tài)的馬鈴薯塊莖蛾對不同化合物的EAG反應相對值經(jīng)Turkey檢驗差異顯著(P<0.05)
2.2 馬鈴薯塊莖蛾對6種揮發(fā)性化合物的EAG劑量反應
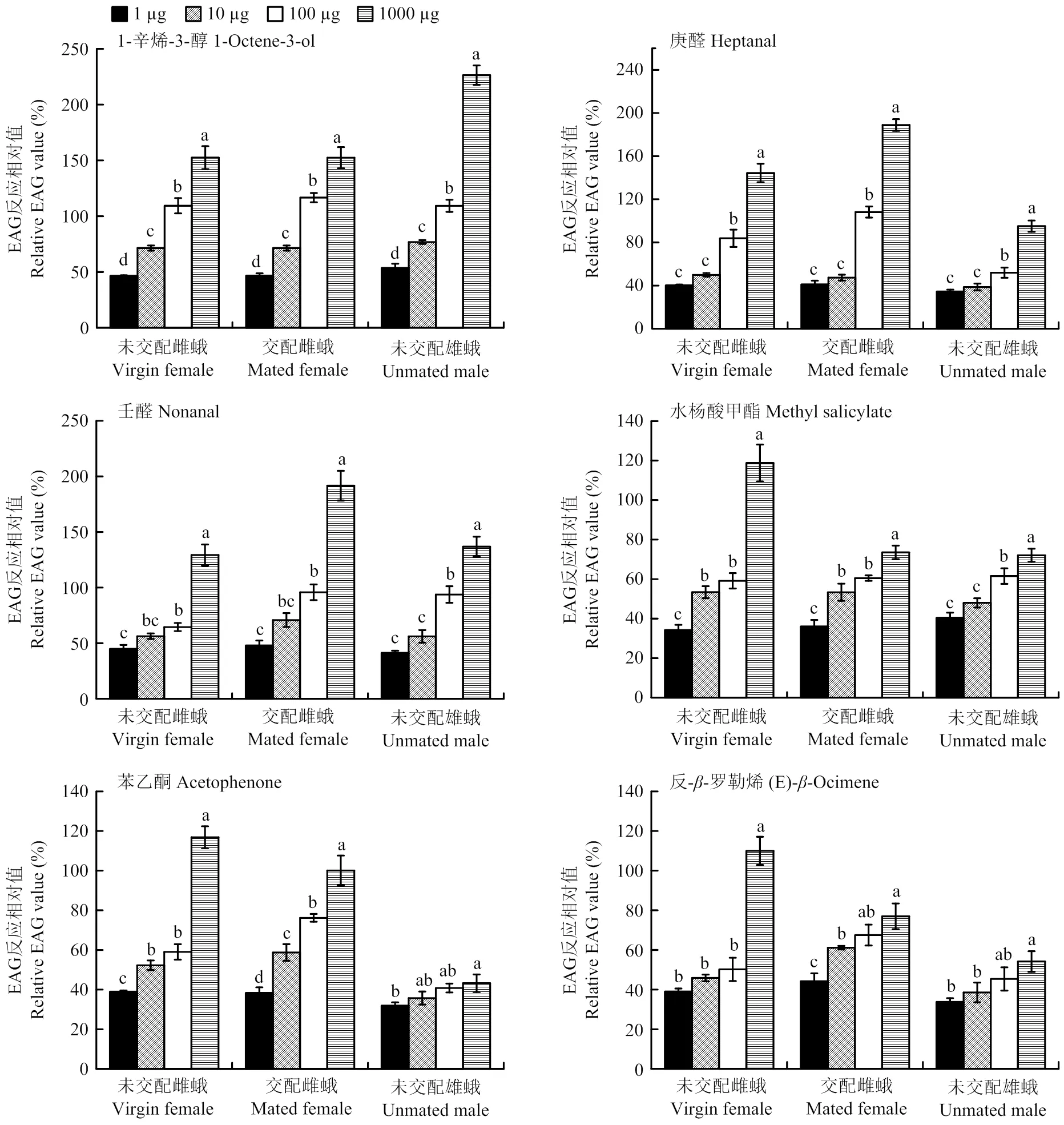
當刺激劑量分別為1、10、100 μg時,馬鈴薯塊莖蛾未交配雌蛾、交配雌蛾、未交配雄蛾對1-辛烯-3-醇、庚醛、壬醛、苯乙酮、水楊酸甲酯、反--羅勒烯的EAG反應值均顯著高于對照(<0.05)。對不同劑量的1-辛烯-3-醇的EAG反應,3種生理狀態(tài)的塊莖蛾均為1 000 μg>100 μg>10 μg>1 μg;對庚醛的EAG反應,則為1 000 μg>100 μg>10、1 μg;對于壬醛、水楊酸甲酯的EAG反應,未交配雌蟲與交配雌蟲對不同劑量的反應趨勢一致;對于苯乙酮、反--羅勒烯的反應,3種生理狀態(tài)的馬鈴薯塊莖蛾對不同劑量的反應趨勢各不相同(圖3)。總之,在測試劑量范圍內(nèi),3種生理狀態(tài)的馬鈴薯塊莖蛾對6種化合物的EAG反應強度隨刺激劑量的增加呈現(xiàn)增強的趨勢,且均未達到飽和閾值。
不同小寫字母表示未交配雌蛾、交配雌蛾、未交配雄蛾對同一化合物的EAG反應相對值經(jīng)Turkey檢驗差異顯著(P<0.05)
3 討論
本試驗所選取的16種馬鈴薯揮發(fā)物1-辛烯-3-醇、丁醇、庚醛、辛醛、壬醛、癸醛、苯乙醇、苯乙醛、苯乙酮、水楊酸甲酯、-蒎烯、對-聚傘花素、反--羅勒烯、桉葉油醇、反--法尼烯、茉莉酸甲酯在一定的劑量下均會引起馬鈴薯塊莖蛾的EAG反應,表明馬鈴薯塊莖蛾觸角中的嗅覺外周神經(jīng)系統(tǒng)對馬鈴薯氣味的反應具有廣譜性,這可以使馬鈴薯塊莖蛾通過辨別環(huán)境中的不同氣味,從而尋找到適合的寄主植物。但是,對于同種化合物,雌、雄馬鈴薯塊莖蛾敏感性表現(xiàn)出不同程度的差異,這可能是由于雌蛾通常擁有的感受寄主植物揮發(fā)物的嗅覺感受神經(jīng)元數(shù)量比雄蛾高[26]。本研究結(jié)果與歐洲玉米螟()雌、雄蟲對寄主植物揮發(fā)物的EAG反應相似[27],但與蘋果蠹蛾和女貞細卷蛾()對寄主植物揮發(fā)物EAG反應的結(jié)果不同,后者雌、雄蟲的敏感性不存在差異[28-29]。馬鈴薯塊莖蛾雌蟲對寄主揮發(fā)物組分的敏感性因交配狀態(tài)而異,在其他蛾類昆蟲上也有類似結(jié)果[18-20]。
在馬鈴薯塊莖蛾對劑量為1 000 μg馬鈴薯揮發(fā)性化合物的EAG反應中,交配雌蛾對于庚醛、壬醛、-蒎烯的電生理反應值顯著高于未交配雌蛾和雄蛾。這3種化合物在低劑量下對馬鈴薯塊莖蛾產(chǎn)卵具有引誘作用,庚醛在高劑量下對產(chǎn)卵具有驅(qū)避作用,壬醛對羽化3 d的交配雌蛾定向兼具引誘作用,庚醛和壬醛對未交配雄蛾的定向行為均不起作用[7,11,30]。因此,交配的雌蛾對這3種化合物更為敏感。未交配雄蛾對1-辛烯-3醇、苯乙醇、苯乙醛的EAG反應值顯著高于未交配雌蛾和交配雌蛾,交配雌蛾與未交配雌蛾之間無顯著差異。有研究表明,高劑量(4、40、400 mg)的1-辛烯-3醇具有干擾馬鈴薯塊莖蛾在馬鈴薯塊莖上產(chǎn)卵在作用[31],苯乙醇則是低濃度下對產(chǎn)卵具有引誘作用,高濃度下具有驅(qū)避作用[8],但是對于雄蛾的作用目前還不清楚。筆者推測,在雄蛾為了找到配偶而向寄主植物定向的過程中,寄主植物釋放的這3種化合物(低劑量)有可能起著重要作用。未交配雌蛾對辛醛、水楊酸甲酯、反--羅勒烯的EAG反應值顯著高于交配雌蛾和未交配雄蛾。辛醛對羽化后1 d的未交配雄蛾的定向具有驅(qū)避作用,對羽化后2—3 d的已交配雌蛾產(chǎn)卵具有驅(qū)避作用[11,31];水楊酸甲酯在高濃度下對雌蛾產(chǎn)卵具有引誘作用[8]。但辛醛、水楊酸甲酯、反--羅勒烯對未交配雌蟲的作用目前還不清楚。3種生理狀態(tài)的馬鈴薯塊莖蛾對癸醛、桉葉油醇、反--法尼烯、茉莉酸甲酯的EAG反應值之間無顯著差異。癸醛對交配雌蛾定向具有引誘作用,對未交配雄蛾的定向不起作用,對雌蛾產(chǎn)卵具有引誘作用[11];桉葉油醇在低濃度時對馬鈴薯塊莖蛾雌蛾產(chǎn)卵具有引誘作用,而在高濃度時具有驅(qū)避作用[8,30];茉莉酸甲酯在低濃度時(接近完整馬鈴薯塊莖釋放的濃度)對交配雌蛾定向具有引誘作用。總之,馬鈴薯塊莖蛾對某個化合物具有EAG反應只是表明該蟲能夠感受到該化合物,反應值的大小也僅表明其對該化合物的敏感性,由于化合物的揮發(fā)性不同,導致即使是同劑量的不同化合物,最終被馬鈴薯塊莖蛾觸角上的受體結(jié)合蛋白結(jié)合的劑量也不同。因此,部分化合物的功能需要行為和田間實驗進一步驗證。
不同小寫字母表示同一生理狀態(tài)的馬鈴薯塊莖蛾對不同劑量的同一化合物的EAG反應相對值經(jīng)Turkey檢驗差異顯著(P<0.05)
Das等研究發(fā)現(xiàn),同樣是1 000 μg的化合物,馬鈴薯塊莖蛾雌蟲對反-2-己烯醛、苯乙酮、苯乙醇、-蒎烯的EAG反應值顯著高于雄蟲,而對于苯乙醛的反應,雌、雄蟲的反應值無差異[21]。這與本研究的結(jié)果不同。造成這種差異的原因可能有兩個方面:(1)Das等進行EAG測定時,沒有檢查馬鈴薯塊莖蛾的交配狀態(tài),采用的是羽化后2—3 d雌、雄蟲,本研究僅采用羽化后3 d雌、雄蟲,而且嚴格區(qū)分了其交配狀態(tài);(2)Das等測定的是印度種群,本研究測定的為中國種群,昆蟲的地域不同,也會導致其對同種揮發(fā)物的敏感性不同。
另外,馬鈴薯塊莖蛾雌、雄蟲對同劑量的同種化合物反應不同,有可能是由于其觸角上的嗅覺感器對各化合物的感受不同所導致。Anfora等測定了馬鈴薯塊莖蛾雌、雄蛾觸角上的長毛形感受器對辛醛、庚醛、壬醛、1-辛烯-3-醇的單感器反應,發(fā)現(xiàn)在測定的雌蛾觸角上的19個感器中,有17個感器至少對2種或2種以上的化合物具有電生理反應,有一個毛形感器對12種化合物具有電生理反應,分別有12、10、9、7個感器對1-辛烯-3-醇、辛醛、壬醛、庚醛發(fā)生反應;雄蟲觸角上的同種感器(測定的感器數(shù)為16個)卻對辛醛、壬醛、庚醛無單感器反應,僅對1-辛烯-3-醇有反應(2個感器有反應)[31]。同種、同性別的昆蟲在不同時期生命活動不同,導致與之密切相關(guān)的嗅覺蛋白(例如氣味結(jié)合蛋白、嗅覺受體等)的表達量不同[32-33]。馬鈴薯塊莖蛾未交配雌蟲尋找寄主的目的是在寄主上求偶,交配雌蟲尋找寄主的目的是在寄主上產(chǎn)卵。因此,未交配雌蟲和交配雌蟲對同種化合物EAG反應的差異,可能主要源于觸角上嗅覺感器內(nèi)嗅覺蛋白表達量的差異。
4 結(jié)論
馬鈴薯塊莖蛾未交配雌蟲、交配雌蟲、未交配雄蟲基于不同的生理需求和目的,對不同類別化合物的電生理反應表現(xiàn)出不同的敏感性,未交配雌蛾和雄蛾對脂肪族醇類化合物較敏感,交配雌蛾對脂肪族醛類化合物較敏感,3種不同生理狀態(tài)的馬鈴薯塊莖蛾對萜類化合物均不敏感。交配狀態(tài)直接影響馬鈴薯塊莖對寄主揮發(fā)物電生理反應,交配雌蛾對于庚醛、壬醛、-蒎烯的敏感性高于未交配雌蛾和雄蛾。對同一化合物不同劑量的電生理反應,交配雌蛾同樣更為敏感。
[1] RONDON S I. The potato tuber worm: a literature review of its biology, ecology and control. American Journal of Potato Research, 2010, 87(2): 149-166.
[2] RONDON S I. Decoding(Lepidoptera: Gelechiidae) in the new age of change. Journal of Integrative Agriculture, 2020, 19(2): 316-324.
[3] GAO Y. Potato tuberworm: A threat for China potatoes. Entomology, Ornithology and Herpetology: Current Research, 2018, 7(2): 1000e132.
[4] 高玉林, 徐進, 劉寧, 周倩, 丁新華, 詹家綏, 成新躍, 黃劍, 魯宇文, 楊宇紅. 我國馬鈴薯病蟲害發(fā)生現(xiàn)狀與防控策略. 植物保護, 2019, 45(5): 106-111.
GAO Y L, XU J, LIU N, ZHOU Q, DING X H, ZHAN J S, CHENG X Y, HUANG J, LU Y W, YANG Y H. Current status and management strategies for potato insect pests and diseases in China. Plant Protection, 2019, 45(5): 106-111. (in Chinese)
[5] ARAB A, TRIGO J R, LOURENC?O A L, PEIXOTO A M, RAMOS F, BENTO J M S. Differential attractiveness of potato tuber volatiles to(Gelechiidae) and the predator(Anthocoridae). Journal of Chemical Ecology, 2007, 33(10): 1845-1855.
[6] SHARABY A, ABDEL-RAHMAN H, MOAWAD S. Biological effects of some natural and chemical compounds on the potato tuber moth,Zeller (Lepidoptera: Gelechiidae). Saudi Journal of Biological Sciences, 2009, 16(1): 1-9.
[7] 馬艷粉, 胥勇, 肖春. 10種寄主植物揮發(fā)物對馬鈴薯塊莖蛾產(chǎn)卵的引誘作用. 中國生物防治學報, 2012, 28(3): 448-452.
MA Y F, XU Y, XIAO C. Oviposition attraction effect of ten host-plant volatiles on potato tuber moth,. Chinese Journal of Biological Control, 2012, 28(3): 448-452. (in Chinese)
[8] 馬艷粉, 張曉梅, 胥勇, 肖春. 滇楊揮發(fā)物成分對馬鈴薯塊莖蛾產(chǎn)卵選擇的影響. 植物保護, 2016, 42(2): 99-103.
MA Y F, ZHANG X M, XU Y, XIAO C. Effects of volatiles fromon oviposition preferece of potato tuber moth,. Plant Protection, 2016, 42(2): 99-103. (in Chinese)
[9] 劉燕, 謝冬生, 熊焰, 王春婭, 任智強, 肖春. 庚醛與桉葉油醇組合對馬鈴薯塊莖蛾產(chǎn)卵選擇的影響. 植物保護, 2016, 42(3): 99-103.
LIU Y, XIE D S, XIONG Y, WANG C Y, REN Z Q, XIAO C. Effects of combination of cineole with heptanal on oviposition choices of the potato tuber moth,. Plant Protection, 2016, 42(3): 99-103. (in Chinese)
[10] DEKEBO A, ARYAL S, JUNG C. Olfactory responses of adult potato tuber moth,(Zeller) measured by attraction relative to the tomato leaf volatiles. Journal of Asia-Pacific Entomology, 2019, 22(2): 611-618.
[11] LI X, ZHANG X G, XIAO C, GAO Y L, DONG W X. Behavioral responses of potato tuber moth () to tobacco plant volatiles. Journal of Integrative Agriculture, 2020, 19(2): 325-332.
[12] 張小嬌, 錢誠, 楊六三, 劉洪翠, 李成云, 董文霞. 不同生理狀態(tài)的番石榴實蠅對寄主氣味的行為反應. 應用昆蟲學報, 2020, 57(1): 166-172.
ZHANG X J, QIAN C, YANG L S, LIU H C, LI C Y, DONG W X. Behavioral responses of sexually immature, mature virgin, and mated, guava fruit flies to host plant odor.Chinese Journal of Applied Entomology, 2020, 57(1): 166-172. (in Chinese)
[13] MECHABER W L, CAPALDO C T, HILDEBRAND J G. Behavioral responses of adult tobacco hornworms,, to host plant volatiles change with age and mating status. Journal of Insect Science, 2002, 2(1): 5.
[14] YAN F, BENGTSSON M, WITZGALL P. Behavioral response of female codling moths,, to apple volatiles. Journal of Chemical Ecology, 1999, 25(6): 1343-1351.
[15] LEMMEN-LECHELT J K, WIST T J, EVENDEN M L. State-dependent plasticity in response to host-plant volatiles in a long-lived moth,(Lepidoptera: Gracillariidae). Journal of Chemical Ecology, 2018, 44(3): 276-287.
[16] TANG R, SU M W, ZHANG Z N. Electroantennogram responses of an invasive species fall webworm () to host volatile compounds. Chinese Science Bulletin, 2012, 57(35): 4560-4568.
[17] ZHANG Z, BIAN L, SUN X, LUO Z, XIN Z, LUO F, CHEN Z. Electrophysiological and behavioural responses of the tea geometrid(Lepidoptera: Geometridae) to volatiles from a non-host plant, rosemary,(Lamiaceae). Pest Management Science, 2015, 71(1): 96-104.
[18] SHU S, GRANT G G, LANGEVIN D, LOMBARDO D A, MACDONALD L. Oviposition and electroantennogram responses of(Lepidoptera: Pyralidae) elicited by monoterpenes and enantiomers from eastern with pine. Journal of Chemical Ecology, 1997, 23(1): 35-50.
[19] MARTEL V, ANDERSON P, HANSSON B S, SCHLYTER F. Peripheral modulation of olfaction by physiological state in the Egyptian leaf worm(Lepidoptera: Noctuidae). Journal of Insect Physiology, 2009, 55(9): 793-797.
[20] SAVEER A M, KROMANN S H, BIRGERSSON G, BENGTSSON M, LINDBLOM T, BALKENIUS A, HANSSON B S, WITZGALL P, BECHER P G, IGNELL R. Floral to green: mating switches moth olfactory coding and preference. Proceedings of Royal Society B:Biological Sciences, 2012, 279(1737): 2314-2322.
[21] DAS P D, RAINA R, PRASAD A R, SEN A. Electroantennogram responses of the potato tuber moth,(Lepidoptera; Gelichiidae) to plant volatiles. Journal of Biosciences, 2007, 32(2): 339-349.
[22] 王國紅, 劉勇, 戈峰, 吳開拓. 粉蝶盤絨繭蜂中國和荷蘭種群學習行為及EAG反應的比較. 生態(tài)學報, 2012, 32(2): 351-360.
WANG G H, LIU Y, GE F, WU K T. Comparative study on learning behavior and electroantennogram responses in two geographic races of. Acta Ecologica Sinica, 2012, 32(2): 351-360. (in Chinese)
[23] BOLTER C J, DICKE M, VAN LOON J J A, VISSER J H, POSTHUMUS M A. Attraction of Colorado potato beetle to herbivore-damaged plants during herbivory and after its termination. Journal of Chemical Ecology, 1997, 23(4): 1003-1023.
[24] AGELOPOULOS N G, CHAMBERLAIN K, PICKETT J A. Factors affecting volatile emissions of intact potato plants,: Variability of quantities and stability of ratios.Journal of Chemical Ecology, 2000, 26(2): 497-511.
[25] KARLSSON M F, BIRGERSSON G, PRADO A M C, BOSA F, BENGTSSON M, WITZGALL P. Plant odor analysis of potato: Response of Guatemalan moth to above- and background potato volatiles. Journal of Agricultural and Food Chemistry, 2009, 57(13): 5903-5909.
[26] BRUCE T J A, PICKETT J A. Perception of plant volatile blends by herbivorous insects—finding the right mix. Phytochemistry, 2011, 72(13): 1605-1611.
[27] SOLE J, SANS A, RIBA M, GUERRERO A. Behavioural and electrophysiological responses of the European corn borerto host-plant volatiles and related chemicals. Physiological Entomology, 2010, 35: 354-363.
[28] ANESBO L, CORACINI M D A, BENGTSSON M, LIBLIKAS I, RAMIREZ M, BORG-KARLSON A K, TASIN M, WITZGALL P. Antennal and behavioural response of codling mothto plant volatiles. Journal of Applied Entomology, 2004, 128(7): 488-493.
[29] SCHMIDT-BUSSER D, VON ARX M, CONNETABLE S, GUERIN P M. Identification of host-plant chemical stimuli for the European grape berry moth. Physiological Entomology, 2011, 36: 101-110.
[30] MA Y F, XIAO C. Push-pull effects of three plant secondary metabolites on oviposition of the potato tuber moth,. Journal of Insect Science, 2013, 13(1): 128.
[31] ANFORA G, VITAGLIANO S, LARSSON M C, WITZGALL P, TASIN M, GERMINARA G S, DE CRISTOFARO A. Disruption of(Lepidoptera: Gelechiidae) oviposition by the application of host plant volatiles. Pest Management Science, 2014,70: 628-635.
[32] 張治科, 吳圣勇, 雷仲仁. 西花薊馬氣味結(jié)合蛋白的cDNA克隆、序列分析及時空表達. 中國農(nóng)業(yè)科學, 2016, 49(6): 1106-1116.
ZHANG Z K, WU S Y, LEI Z R. Cloning, sequence analysis and expression profile of an odorant binding protein gene in western flower thrips ().Scientia Agricultura Sinica, 2016, 49(6): 1106-1116. (in Chinese)
[33] 陳麗慧, 李梅梅, 陳秀琳, 仵均祥, 許向利. 梨小食心蟲普通氣味受體基因的克隆及表達分析. 昆蟲學報, 2019, 62(4): 418-427.
CHEN L H, LI M M, CHEN X L, WU J X, XU X L. Cloning and expression profiling of general odorant receptor genethe oriental fruit moth,(Lepidoptera: Tortricidae). Acta Entomologica Sinica, 2019, 62(4): 418-427. (in Chinese)
Electroantennogram Responses ofof Different Sexes and Mating States to Potato Volatiles
LI Xiang, ZHANG XiaoJiao, XIAO Chun, DONG WenXia
College of Plant Protection/State Key Laboratory for Conservation and Utilization of Bio-Resources in Yunnan, Yunnan Agricultural University, Kunming 650201
【】The objective of this study is find out the difference of electrophysiological responses of potato tuberworm () with different sexes and mating states to potato volatiles, and to provide a reference for the chemical ecological control of.【】Electroantennogram (EAG) responses of virgin female, mated female and unmated maleto 16 volatile compounds (including 6 aliphatic compounds viz.1-octen-3-ol,1-butanol,heptanal,octanal, nonanal and decanal, 4 aromatic compounds viz. phenethyl alcohol, phenylacetaldehyde, acetophenone and methyl salicylate, and 6 terpenoids viz.-pinene,-cymene, ()--ocimene, 1,8-cineole, ()--farnesene and methyl jasmonate) released from potato tubers and plants were recorded, and the dose-responses to 6 compounds (including 1-octen-3-ol, heptanal, nonanal, acetophenone, methyl salicylate and ()--ocimene)were also conducted.【】All the tested compounds could elicit EAG responses of. Among them, the aliphatic alcohols elicited significantly greater EAG values in the virgin females and the unmated males, and aliphatic aldehydes elicited significantly greater EAG values in the mated females. However, the terpenoids elicited lower EAG values in all the tested. At the dose of 1 000 μg,the same compound elicited varied antennal responses from thewith different sexes and mating states. Octanal, methyl salicylate and ()--ocimene elicited stronger responses in virgin females than those in the mated females and the unmated males, while heptanal, nonanal and-pinene elicited stronger responses in the mated females than those in the others. The compounds 1-octene-3-ol, phenethyl alcohol and phenylacetaldehyde elicited stronger responses in the males than those in the females. At the tested dose series (1, 10, 100 and 1 000 μg), EAG responses ofat 3 physiological states to 6 individual compounds increased with the increase of stimulation dose, and were far from saturation thresholds. For the same compound at different doses, EAG responses of the mated females varied significantly.【】The virgin females and the unmated males ofare more sensitive to aliphatic alcohols, and the mated females are more sensitive to aliphatic aldehydes. However, both the females and males are less sensitive to terpenoid compounds. Among the responses to the individual compound, the mated females are more sensitive to heptanal,nonanal and-pinene than the virgin females and the unmated males, so are their responses to the same compound at different doses.
potato tuberworm (); potato volatile;electroantennogram response

10.3864/j.issn.0578-1752.2021.03.009
2020-06-30;
2020-07-28
國家自然科學基金(31560607)、國家重點研發(fā)計劃(2018YFD0200802)
李祥,E-mail:lixiang0217@126.com。通信作者董文霞,E-mail:dongwenxia@163.com
(責任編輯 岳梅)
