大黃魚脊椎骨及其早期發育研究
2021-03-11 05:44:02黃偉卿周逢芳阮少江林培華周振平丁建發劉家富楊俊凡
海洋科學 2021年2期
黃偉卿 , 張 藝, 周逢芳, 阮少江, 林培華 周振平 丁建發, 劉家富,楊俊凡
(1. 寧德市鼎誠水產有限公司, 福建 寧德 352100; 2. 寧德師范學院 生命科學學院, 福建 寧德352100; 3. 寧德市水產技術推廣站, 福建 寧德 352100)
大黃魚(Larimichthys crocea)屬于硬骨魚綱(Osteichthyes)、鱸形目(Perciformes)、石首魚科(Sciaenidae)、黃魚屬(Larimichthys), 為暖水性集群洄游魚類, 主要分布在中國南海、東海和黃海南部, 因其肉質鮮美、體色金黃、嘴唇深紅、吉祥象征, 深受消費者喜愛[1-3]。2019 年產量達22.6 萬噸, 居中國海水養殖魚類產量之首, 已成為中國最大規模的海水養殖魚類和八大優勢出口養殖水產品之一[1-5]。目前, 關于大黃魚的研究主要集中在養殖技術[6]、肌肉營養[7]、遺傳育種[8]和病害防治[9]等方向, 并取得了豐碩的成果。
魚類的骨骼作為魚體的基本結構, 能夠支撐魚類的身體、保護內臟器官以及在協調肌肉的運動方面起重要作用。在魚類的長期進化過程中, 其骨骼發育時序、形態構造與其棲息環境相適應, 已經成為研究魚類系統進化和分類的重要依據, 也是深入研究魚類組織器官功能發育的重要途徑[10-12]。于超勇等[12]對 1~60 日齡的大瀧六線魚(Hexagrammos otakii)仔稚魚脊椎骨發育研究表明, 脊柱椎體于28日齡時開始由頭端向尾端骨化, 至50 日齡時脊柱椎體全部完成骨骼元件骨化, 脊柱及附肢骨骼早期發育與其巡游運動模式向機動運動模式的轉變、攝食能力增強密切相關。曹曉穎等[13]對1~35 日齡鱖(Siniperca chuatsi)仔稚魚脊椎骨發育研究發現, 脊柱于15 日齡由前向后骨化, 20 日齡背肋與腹肋由基部向末端骨化, 29 日齡骨化完成。脈弓與脈棘、髓弓與髓棘均由前向后、由基部向末端骨化, 脈棘與髓棘骨化時間晚于相應的椎體, 骼骨化發育與其早期運動、攝食與御敵等行為密切相關。王秋榮等[14]研究青石斑魚(Epinephelus awoara)骨骼發育異常時發現, 脊柱發育異常主要表現在脊柱前凸、脊柱后凸和脊柱側凸以及椎骨的愈合、變形等癥狀。在大黃魚骨骼的研究方面, 劉家富[1]介紹了閩-粵東族大黃魚的脊椎骨有26 個骨節, 由11 個腹椎和15 個尾椎組成。丘書院[15]介紹了大黃魚骨骼標本制作的5種使用方法, 并且提出最后1 個尾椎有6 個尾下骨和上面3 個尾上骨。王秋榮等[16]對人工繁育的大黃魚仔稚魚的脊柱、胸鰭骨骼以及尾鰭骨骼的發育進行探究, 提出中軸骨骼和附肢骨骼的不斷發育完善,能夠增強大黃魚呼吸、攝食、游泳的機能。王映等[17]運用了 Micro CT 技術對大黃魚全魚骨骼系統進行了掃描與三維重建用來區分大黃魚野生群體與養殖群體。但關于大黃魚脊椎骨不同數量骨節發育和異常現象方面的研究尚未見報道, 旨為大黃魚的生物學特征提供理論依據。
1 材料與方法
1.1 大黃魚的來源
2019 年5 月, 從寧德市鼎誠水產有限公司養殖基地2 400 尾閩-粵東族大黃魚中隨機挑選平均體長34.26±1.36 cm, 體質量677.1±9.49 g 成魚48 尾進行骨骼節數和生物特性進行分析。將上述成魚在室內水溫24℃下, 采用生物餌料牡蠣進行營養強化后作為親本進行人工催產, 并從子代5~40 日齡仔稚魚中, 每個日齡隨機取樣30 尾, 進行骨骼節數及生物特性進行研究。
1.2 樣本處理
成魚的處理: 48 尾大黃魚成魚采用日本·米卡薩型號的X 光機(型號: HF4WA)進行骨骼掃描后, 將其煮熟并剔除肌肉, 對脊椎骨骨節數進行統計、測量和拍照分析。
幼魚的處理: 大黃魚幼魚取樣后, 置于 Leica S9i 體視顯微鏡下進行觀察, 用顯微鏡配置的軟件測量全長、體長以及骨骼長度等數據后, 用10%的福爾馬林進行固定保存, 供日后制作骨骼標本時使用。
1.3 軟骨-硬骨雙染色法[10-14]對脊椎骨進行染色
固定: 將固定于10%福爾馬林3 d 后的仔稚幼魚置換于蒸餾水中浸泡1 d。
脫水: 將仔稚魚標本浸泡在梯度酒精(20%→40%→60%→75%)中, 每種梯度浸泡 1 d, 最終在75%酒精中置于冰箱(4℃)中保存, 染色前, 再將仔稚魚置換于95%的酒精中浸泡1 d。
軟骨染色: 將樣品浸泡于配制好的軟骨染色液(20 mL 冰醋酸+80 mL 95%酒精+10 mg Alcian Blue power)中5 h 左右, 直至魚體表面、鰭條末端略有藍色。
中性化: 將上述樣品浸泡于新鮮配制的飽和硼酸鈉溶液中1 d。
漂白: 將上述樣品置換于80 mL(1% KOH)+ 20 mL(3% H2O2)溶液中浸泡1 d。
硬骨染色: 將樣品置換于配制好的硬骨染色液(5 mg Alizarin Red power +100 mL 1% KOH)中浸泡5~9 h(染色時間根據樣品體積大小適當進行調整)。
褪色: 將樣品浸泡于1% KOH 溶液中直到魚體更加透明。最后將樣品置換于甘油和1% KOH 溶液的混合液中, 按照二者不同比例依次更換, 比例如下, 甘油︰1% KOH=1︰3 →甘油︰1% KOH=1︰1→甘油︰1% KOH=3︰1, 各比例分別浸泡1 d, 直到魚體透明, 最后置換于純甘油中保存。
1.4 數據處理
數據采用SPSS17.0 統計分析。用 Duncan’s 多重比較分析進行差異顯著性分析。
2 結果與分析
2.1 大黃魚脊椎骨骨節數及數量性狀分析
閩-粵東族大黃魚成魚脊椎骨骨節數存在著25、26 和27 個3 種情況, 分別占總量的4.17%、83.33%和12.5%(圖1、表1)。
大黃魚脊椎骨不同的骨節數組成情況分別為:25 個骨節的大黃魚是由11 個腹椎和14 個尾椎組成。26 個骨節的大黃魚存在著2 種腹椎與尾椎的組成形式, 一種是由11 個腹椎和15 個尾椎組成,該組成模式占總量的55.6%; 另一種是由12 根腹椎和14 根尾椎組成, 該組成模式占總量的44.4%。27 個骨節的大黃魚是由12 個腹椎和15 個尾椎組成(表1)。
27 骨節的大黃魚的髓棘/椎體、橫突/椎體、橫突/髓棘比值分別為(193.38±0.488)%、(53.41±0.207)%和(24±0.096)%, 顯著高于25 骨節(P<0.05); 26 骨節大黃魚的脈棘/椎體比值為(185.47±1.875)%, 顯著高于25 和27 骨節(P<0.05); 25 骨節大黃魚的脈棘/髓棘的比值為(125.44±0.21)%, 顯著高于 26 和 27 骨節(P<0.05)(表2)。

圖1 大黃魚的脊椎骨Fig. 1 Vertebrae of a large yellow croakera. 25 骨節; b. 26 骨節; c. 27 骨節
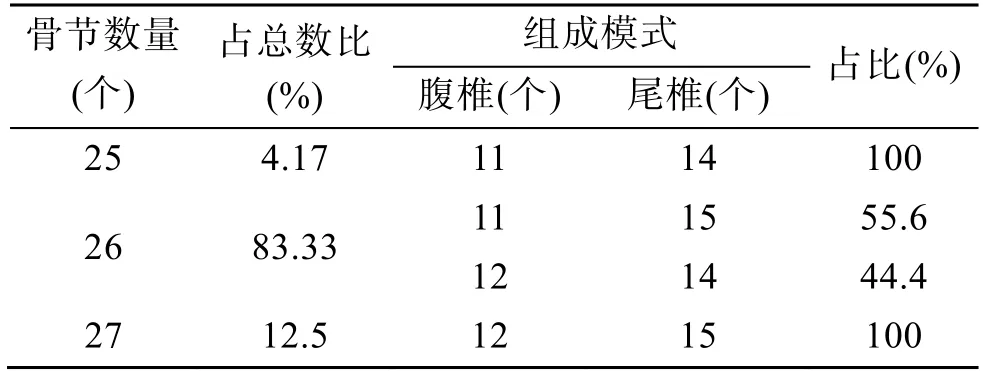
表1 大黃魚骨節數和組成模式Tab. 1 Number of vertebra and the pattern of bone composition of a large yellow croaker
2.2 大黃魚脊椎骨的發育和發育異常現象
大黃魚在5 日齡的脊索呈線狀未分節(圖2a); 8日齡時脊索出現明顯分節, 脊柱前端出現兩個髓弓(圖2b); 15 日齡時尾下骨以及脈弓發育明顯, 以軟骨組織形式出現(圖2c); 16 日齡時隨著尾下骨的增多, 著生在尾部脊索上的脈弓、髓弓隨之出現(圖2d), 發育過程中脈弓比髓弓的生長速度快, 但在此期間仔稚魚的脊柱尚未骨化。隨著仔稚魚脊柱的椎體從頭部開始沿著尾部的方向開始骨化, 21 日齡仔稚魚清晰可見21~22 個脊椎骨, 髓棘、脈棘分別由髓弓、脈弓末梢延長形成(圖 2e)。24 日齡發育出22~23 個脊椎骨(圖2f), 26 日齡仔稚魚的尾桿骨開始骨化(圖2g), 38 日齡仔稚魚脊柱完成骨化(圖2h)。脊椎骨發育的骨節數有25、26 以及27 個之分。以26 個骨節為主, 占比總數73.08%, 其次是27 個骨節, 占比總數 15.38%, 25 個骨節最少, 占比總數11.54%(表3)。

表2 3 種骨節大黃魚椎體、脈棘、橫突以及髓棘的關系Tab. 2 Relationship among vertebral body, pulse-spines, transverse processes, and myeloid spines of three kinds of a large yellow croaker with different vertebrae

表3 大黃魚子代脊椎骨骨節數量占比Tab. 3 Proportions of the number of vertebral vertebrae in the larvae of a large yellow croaker
大黃魚仔稚魚脊柱畸形主要表現在椎體分裂和增生。椎體分裂的現象, 主要出現在倒數第二個椎骨,椎體下端分裂, 但不分離。椎體增生常見于在第24~25 個椎骨, 多出的一個椎體, 其體積小、結構不完整、有脈棘、沒有髓棘(圖3)。
大黃魚成魚存在椎體融合以及脈棘和髓棘冗余、分叉的畸形現象: 椎體融合的現象出現在27 個脊椎骨的大黃魚成魚中, 兩個椎體融合為一, 其椎體體積比正常的大, 有2 個髓棘和脈棘對側分布; 脈棘分叉和冗余25、26、27 個脊椎骨的大黃魚成魚都存在, 主要出現在尾椎的位置, 分叉是從脈棘的基部或者中部開始一分為二, 脈棘冗余存在相鄰處或者相近處多一根脈棘, 髓棘分叉現象出現較少, 主要表現在髓棘出現樹枝狀分叉(圖4)。
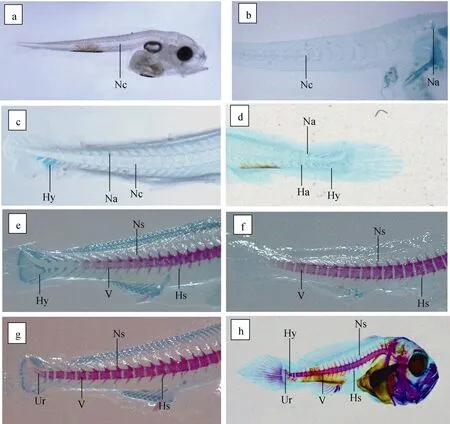
圖2 大黃魚仔稚魚脊椎骨發育Fig 2 Development of vertebrae in a juvenile large yellow croaker

圖3 大黃魚仔稚魚骨骼發育異常Fig. 3 Development of abnormal vertebrae of a juvenile large yellow croaker
2.3 大黃魚仔稚魚脊椎骨的生長
測量 5~40 日齡的大黃魚脊椎骨的生長情況:脊椎骨(y)與日齡(x1)符合二項式關系式為y=0.009 5x12– 0.174 1x1+ 4.260 4(R2= 0.960 7), 平均增長率為0.295 mm/d (圖5a); 與全長(x2)的關系, 符合二項式關系式為y= 0.012 3x22+0.127 3x2+ 2.672 1(R2=0.986 7)(圖5b); 與體長(x3)的關系, 符合二項式關系式為y= 0.001 3x32– 0.624 2x3+0.59(R2= 0.960 7) (圖5c)。

圖4 大黃魚成魚骨骼發育異常Fig. 4 Development of abnormal vertebrae of an adult large yellow croaker
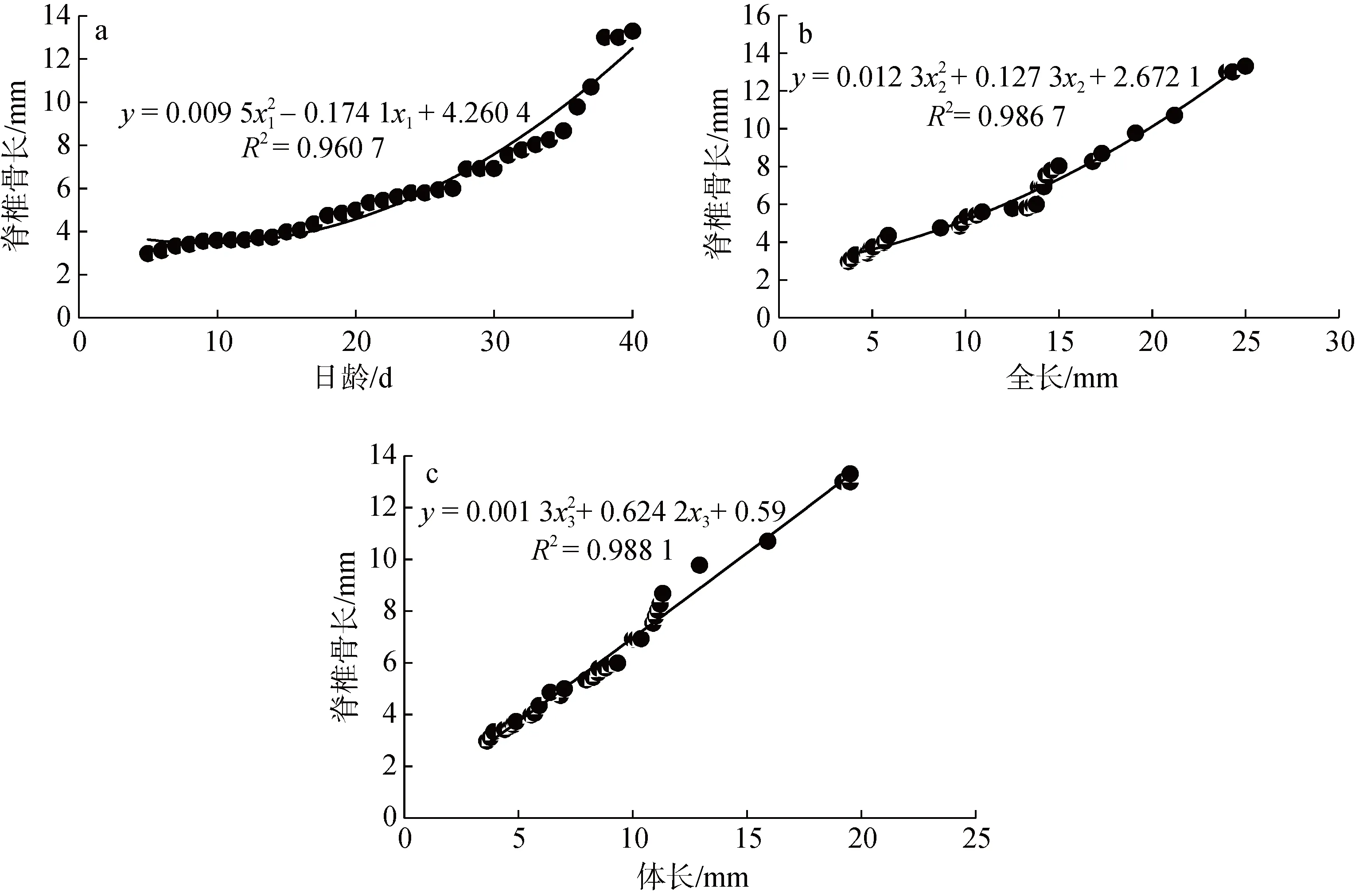
圖5 5~40 日齡大黃魚仔稚骨骼生長Fig. 5 Growth of bones of juvenile fish aged 5-40 days of a large yellow croaker
3 討論
3.1 大黃魚脊椎骨骨節數
本研究發現閩-粵東族成品大黃魚存在25、26和27 個3 種大黃魚脊椎骨骨節情況, 以26 個脊椎骨骨節為主, 主要由11 個腹椎和15 個尾椎組成, 但也存在著12 根腹椎和14 根尾椎組成形式, 這一結果與劉家富[1]和丘書院[15]發現岱衢族、閩-粵東族和硇洲族3 個種群大黃魚中僅岱衢族存在脊椎骨骨節27 個,閩-粵東族與硇洲族大黃魚群體的脊椎骨骨節數存在25 和26 個有所差異, 造成這一差異主要原因可能是由于閩-粵東族已混有岱衢族所致。同時本研究還發現,大黃魚仔稚幼魚中也存在著25、26 和27 個3 種大黃魚脊椎骨骨節, 這一結果與王秋榮[16]等對3~28 日齡閩-粵東族大黃魚仔稚魚骨骼發育研究結果表明16 日齡仔稚魚全部出現了27 個脊椎骨存在差異。
3.2 大黃魚脊椎骨的發育
本文結果與王秋榮[16]等研究相同, 大黃魚的脊柱骨的發育最初從髓弓、脈棘和尾下骨開始的, 椎體的順序是從頭部往尾部方向。王秋榮[16]等還發現發育至16 日齡仔稚魚尾下骨的增多, 著生在脊索上的脈弓、髓弓以軟骨組織的形式清晰呈現, 發育至28 日齡時脊椎骨骨化現象出現。這一結果與本研究發現發育至21 日齡時才開始發生骨化有所差異, 這一差異筆者認為與養殖水溫等水環境的差異相關。
大黃魚骨節的骨化順序與其他一些魚類不同,如 尖 吻 鱸(Lates calcarifer)[11]、 美 洲 鰣(Alosa sapidissima)[18]等, 骨節的骨化則由頭尾兩端向中間進行; 刀鱭(Coilia nasus)[19]則是以體前中部位置為起點向前、后兩端發育。鄧平平[20]等提出魚類的早期發育過程中, 形態結構與功能需求是相互影響的,這可能與其生態習性的形成有關, 但其機理還需要進一步地分析驗證。
3.3 大黃魚脊椎骨骨節發育異常
造成魚類骨骼發育異常的因素有很多, 包括遺傳、營養因素和環境條件等[16]。本研究, 遺傳、培育條件不適以及餌料營養缺陷可能是誘導骨骼發育異常的主要原因。
李凱彬[21]等研究表明骨骼發育異常現象的產生與遺傳有關, 不同的信號通路基因會對骨骼發育過程進行精細調控。本研究, 成魚均發育正常、子代卻出現發育異常的現象, 可能與某些發育異常的表現型受到隱性基因控制或者控制的基因發生突變有關。有研究表明, 缺乏微量元素也會導致脊椎骨骨節發育異常的現象, 如缺乏維生素K 會導致底鳉(Fundulus grandis)椎體融合、髓弓畸形的發生, 缺乏常量礦物質P 會致使大西洋鮭(Salmo salar)脊柱棘部畸形的發生, Ca 元素的攝入不均衡會導致魚類骨骼發育不良以及骨骼代謝異常的現象[14,16,22]。餌料中必需脂肪酸、微量礦物質等營養物質配比不平衡都也會引起魚類脊柱發育異常[23]。本試驗期間主要以輪蟲、豐年蟲以及橈足類等生物餌料為主, 輪蟲營養強化不足、豐年蟲投喂過多或者橈足類來源不同等均可能導致脊椎骨發育異常, 也可能是由于養殖桶偏小導致養殖溫差和溶解氧以及水流速變化大亦或是近岸海域取水造成鹽度差異大等原因[14]造成本次幼魚骨節發育異常的現象。
