施氏獺蛤早期生長性狀遺傳參數和育種值估計
2021-03-11 05:44:14張守都彭慧婧鄭德斌
海洋科學 2021年2期
關鍵詞:生長
鄒 杰, 張守都, 彭慧婧, 鄭德斌
(1. 廣西壯族自治區海洋研究所, 廣西 北海 536000; 2. 青島國家海洋科學研究中心, 山東 青島 266071; 3. 天津渤海水產研究所, 天津 300457)
施氏獺蛤(Lutraria sieboldii)隸屬瓣鰓綱(Lamellibranchia)、異齒亞綱(Heterodonta)、簾蛤目(Veneroida)、蛤蜊科(Mactridae)、獺蛤屬(Lutraria), 俗稱象鼻螺, 為北部灣潮下帶重要經濟貝類之一。施氏獺蛤適合底播增養殖, 部分群體也能適應前中期的筐式養殖, 近年來養殖規模不斷增長, 但養殖群體生長性狀出現衰退, 為此, 施氏獺蛤相關的選擇育種工作隨之開展。生長性狀選擇育種是動物育種傳統且有效的方法, 通過對目標性狀的累代選育, 獲取優良生長性狀的品種。生長性狀受遺傳和環境因素共同影響, 從數量性狀的遺傳機制可知, 影響數量性狀的效應中只有加性效應才能穩定遺傳給后代, 準確估計遺傳參數和育種值是數量性狀育種的核心。約束極大似然法(restricted maximum likelihood, REML)是動物育種遺傳參數估計中較為常用方法[1-2], 最佳線性無偏預測法(best linear unbiased prediction, BLUP)在獲取的個體育種值方面具較高精確性[3], 本研究以施氏獺蛤 2 代選育群體為親本建立全同胞家系, 基于REML 和BLUP 法對早期生長性狀遺傳參數和育種值進行估計和分析, 為施氏獺蛤選擇育種提供基礎資料和參考。
1 材料與方法
1.1 早期家系建立
從施氏獺蛤第二代選育群體(F2)中挑選體型相近的7 個雄性和21 個雌性為親本, 采用巢式設計構建21 個全同胞家系(M1、M2、M3、……、M21)。通過1 雄配3 雌及人工授精方式獲得的幼體分家系分別培養于1 000 L 的水桶中, 用水經紫外線消毒,每3 d 倒桶并調整幼體密度差異, 培育前期以投喂湛江等鞭金藻(Isochrysis zhanjiangensis)為主, 后期以投喂亞心型扁藻(Platymonas subcordiformis)為主, 保持各家系培育操作一致, 附著期保留每桶幼體數4 萬~5 萬個。幼體培育30 d 后將苗種收集以塑料筐裝沙沉水方式進行海上培育, 培育密度0.5 個/cm2, 每個家系留3 000 個體, 海上培育50 d。
1.2 生長性狀測量
用游標卡尺測量殼長、殼高和殼寬, 精確至0.01 mm;用電子天平測量體質量, 精確至0.01 g。每家系取樣30 個體, 30 日齡測量殼長和殼高, 80 日齡測量殼長、殼高、殼寬和體質量。
1.3 數據處理與分析
用SPSS 22.0 統計軟件對測量數據進行正態性檢驗(Kolmogorov-Smirnov 法), 計算各性狀表型值(phenotypic value, PV)與育種值(breeding value, BV)的Pearson 相關系數。
建立混合線性模型(動物模型), 利用ASReml-R V4 軟件采用REML 和BLUP 法對施氏獺蛤進行遺傳參數和育種值估計。單性狀模型估計遺傳力(heritability,h2)和育種值, 兩性狀模型估計表型相關系數(rp)和遺傳相關系數(rg)。
混合線性模型:
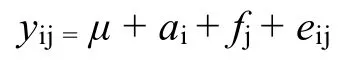
式中,yij為性狀測量值,μ為平均值,ai為加性遺傳效應,fj為全同胞效應,eij為殘差。
性狀1 和性狀2 的表型相關系數和遺傳相關系數計算依據公式:

2 結果與分析
2.1 施氏獺蛤生長性狀描述
施氏獺蛤生長性狀參數統計見表1, 30 日齡和80 日齡的施氏獺蛤生長性狀測量值經Kolmogorov-Smirnov 檢驗均符合正態分布。80 日齡4 個性狀中,殼長、殼高和殼寬性狀的變異系數(coefficient of variation, CV)相近, 體質量變異系數最高, 80 日殼長和殼高性狀變異系數較30 日齡減少, 差異變小。對30 日齡和80 日齡殼長與殼高性狀比較, 極值差比為1.61 和1.89, 均值比為1.53 和2.09, 均表明施氏獺蛤早期發育過程中殼長生長速度較殼高快。
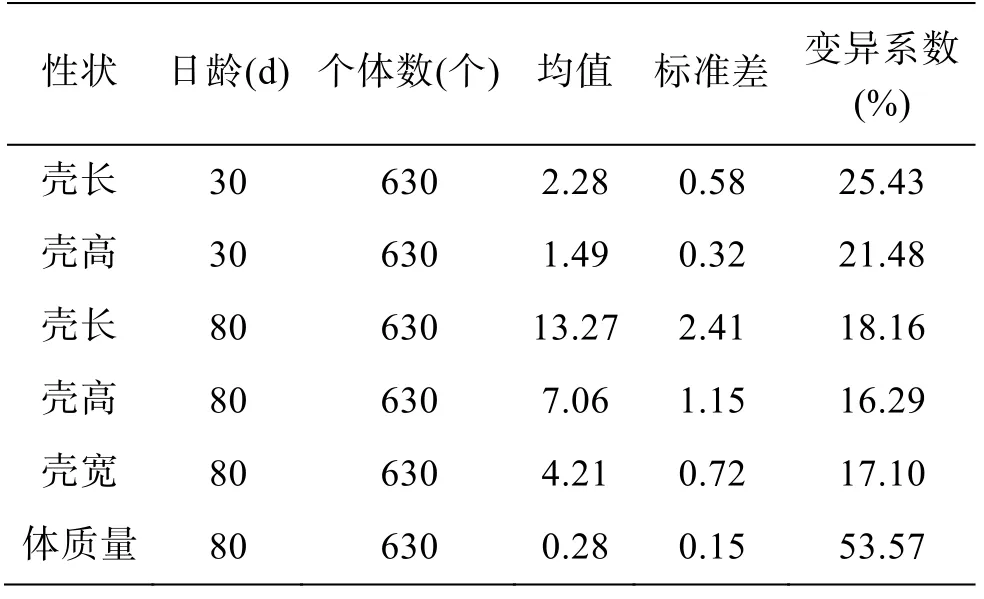
表1 施氏獺蛤生長性狀參數統計Tab. 1 Parametric statistics of the growth traits of Lutraria sieboldii

表2 施氏獺蛤生長性狀方差組分及遺傳力Tab. 2 Variance components estimation of the growth traits and the heritability of Lutraria sieboldii
2.2 遺傳參數估計
單性狀分析施氏獺蛤生長性狀的方差組分和估算遺傳力結果見表2。30 日齡的殼長和殼高性狀遺傳力分別為: 0.31±0.02 和0.25±0.01, 80 日齡的殼長、殼高、殼寬和體質量性狀的遺傳力分別為: 0.35±0.27、0.27±0.20、0.06±0.04 和0.25±0.19, 殼寬性狀遺傳力為低遺傳力, 其余均屬中等遺傳力(h2<0.2 為低遺傳力;0.2≤h2≤0.4 為中等遺傳力;h2>0.4 為高遺傳力[4])。80 日齡的殼長和殼高性狀的遺傳力較30 日齡略有提高, 表明隨時間增長全同胞效應降低。兩性狀分析施氏獺蛤生長性狀間的表型相關和遺傳相關結果見表3。80 日齡4 個性狀間的遺傳相關與表型相關均呈正相關且表現一致性, 遺傳相關系數范圍為 0.96~0.98,相關性均較高, 表型相關系數范圍為0.77~0.96, 殼長、殼高與體質量間表型相關性較高, 結果表明在對早期施氏獺蛤體質量性狀選擇上, 殼長和殼高可作為主要參考性狀。80 日齡殼長和殼高性狀的遺傳相關和表型相關均高于30 日齡, 表明在早期育種性狀參考上, 80 日齡的殼長、殼高性狀參考價值更高。
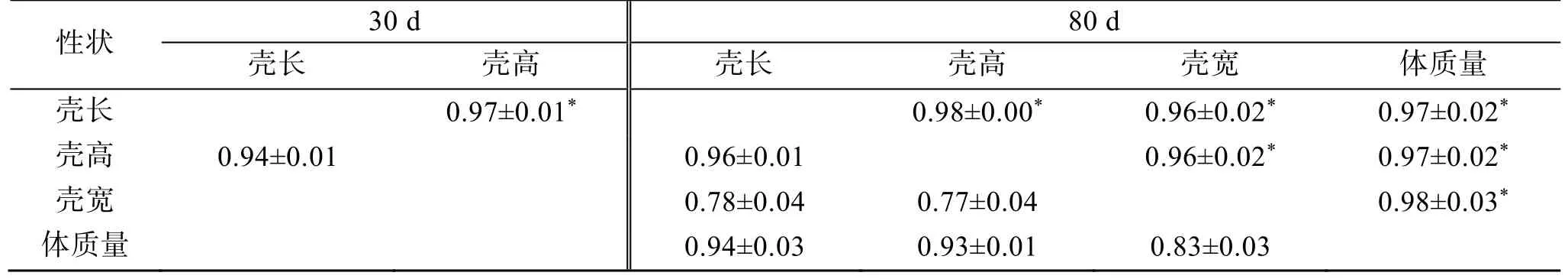
表3 施氏獺蛤生長性狀表型相關和遺傳相關Tab. 3 Genetic correlation and phenotypic correlation of the growth traits of Lutraria sieboldii
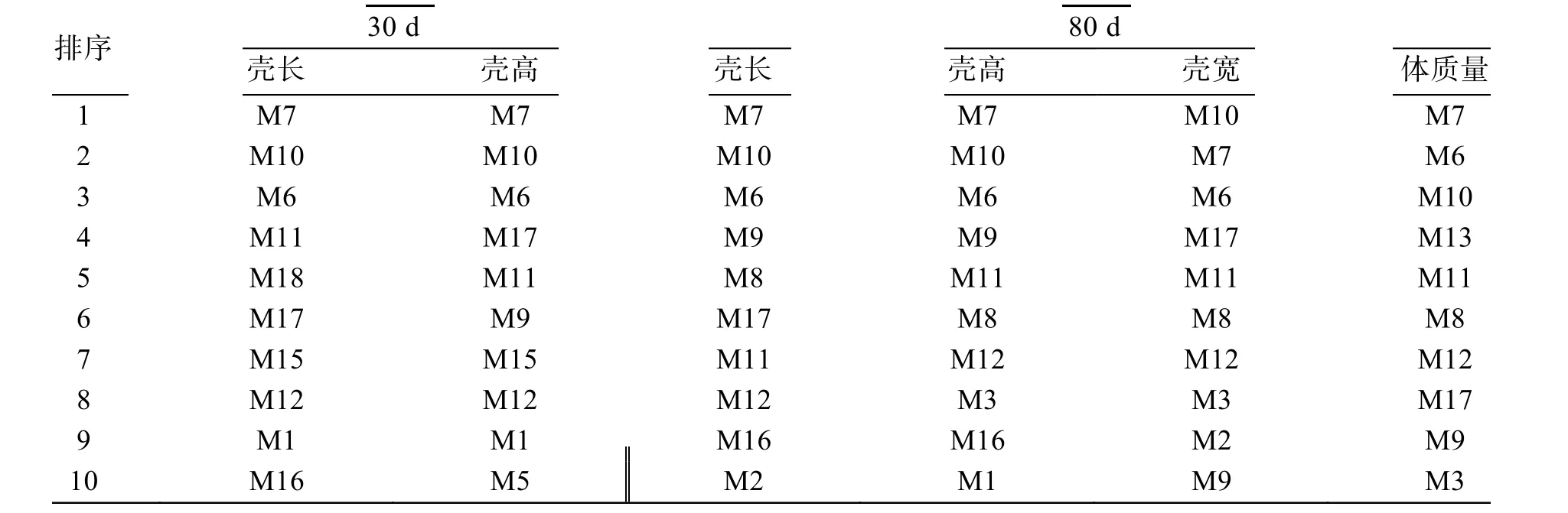
表4 施氏獺蛤不同性狀育種值的家系排序Tab. 4 The ordered families arrangement of breeding values of different traits of Lutraria sieboldii
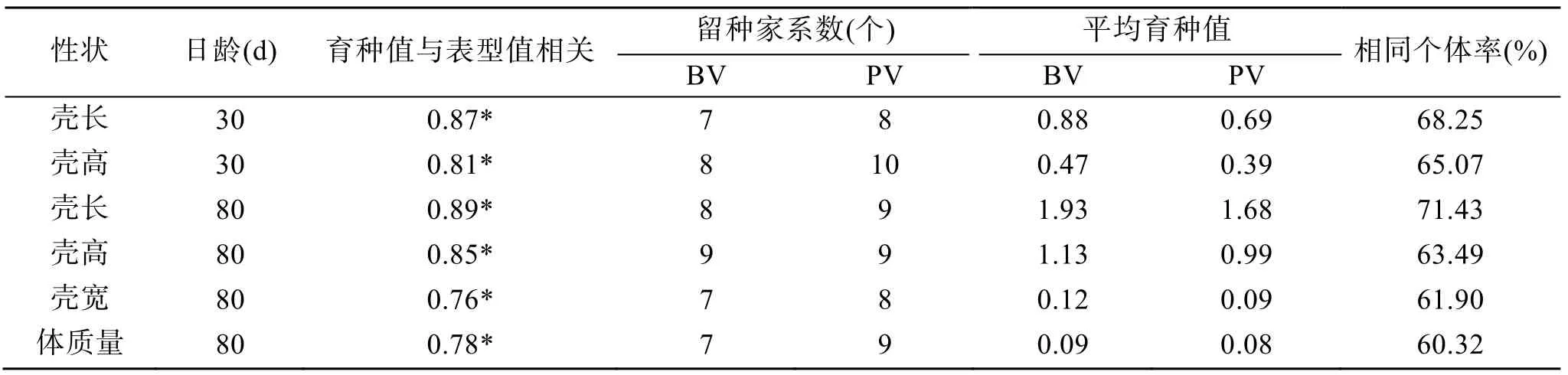
表5 基于表型值和育種值的個體選擇比較(10%留種率)Tab. 5 Comparison of individual selection based on the phenotypic and breeding values (10% selective pressure)
2.3 育種值分析
根據施氏獺蛤不同性狀個體育種值的前10 家系排序結果(表4), 30 日齡殼長、殼高性狀家系相同率為90%, 80 日齡殼長、殼高、殼寬和體質量性狀家系相同率為70%, 對應比較30 日齡和80 日齡的殼長、殼高性狀家系相同率均為70%, 比較結果發現: 不同階段和不同性狀育種值排序存在差異, 但整體表現上呈現一致性, 且家系M7、M10 和M6 于不同階段和不同性狀育種值排序中均排前3, 綜合育種性能突出。個體育種值和表型值的相關性顯著, 基于表型值和育種值的個體留選比較結果(表5, 留種率10%), 留選個體相同率60.32%~71.43%, 根據表型值留選家系數量比根據育種值留選家系數量多0~2個, 根據選留個體的育種值統計比較, 發現兩種方式選擇結果存在差異, 30 日齡施氏獺蛤殼長、殼高育種值選擇效率比其表型值選擇分別提高27%和21%,80 日齡施氏獺蛤殼長、殼高、殼寬和體質量育種值選擇效率比其表型值選擇分別提高16%、14%、30%和13%, 根據育種值選擇效率較高。
3 討論
3.1 遺傳參數
表型相關和遺傳相關是進行間接選擇主要參考依據, 遺傳相關已經剔除了環境影響, 故能比表型相關更確切地反映兩個性狀間的相關程度。陶后全[5]等研究選育馬氏珠母貝(Pinctada martensii)早期遺傳參數得出其表型相關系數(0.747~0.921)高于遺傳相關系數(0.528~0.746), 本研究中施氏獺蛤生長性狀遺傳相關系數(0.96~0.98)高于表型相關系數(0.77~0.96), 與馬氏珠母貝早期性狀相關性表現并不同,說明施氏獺蛤早期選擇參考性狀可靠性較高。
遺傳力是遺傳改良過程中至關重要的遺傳參數,一般認為遺傳力越大選擇效果好。張守都[6]等研究“中科2 號”海灣扇貝(Argopecten irradians)幼蟲期殼長遺傳力時, 6 日齡殼長遺傳力為0.58, 殼長遺傳力較大證明選擇育種對于“中科2 號”早期生長尚具有較大的改良潛力。王慶志[7]等估計了5~25 日齡長牡蠣(Crassostrea gigas)幼體殼高遺傳力(0.161~0.771)和殼長遺傳力(0.139~0.814), 遺傳力表明以殼高或殼長為參數進行選育時, 均可達到改良生長性狀的效果。本試驗中, 30 日齡施氏獺蛤的殼長和殼高遺傳力(0.31, 0.25)為中等遺傳力, 80 日齡的殼長、殼高和體質量遺傳力(0.35, 0.27 和0.25)也為中等遺傳力, 預測施氏獺蛤早期對殼長、殼高和體質量性狀進行選擇時, 可獲得較大的遺傳進度。
養殖環境的不同使估算出的遺傳力產生較大的差異[8-9], 不同遺傳力估計方法也會導致估算結果的不一致。本試驗繁育于秋季進行, 不同家系精卵成熟度存在較大差異, 從而對施氏獺蛤早期的遺傳相關判斷產生非遺傳因素的影響。試驗中施氏獺蛤附著幼體培育密度因存活率不同而存在客觀差異, 造成家系間培育上的環境差異。另外, 試驗所用親本來源于多個半同胞家系混合建立的選育群體, 研究中將選育群體當作無親緣關系進行系譜計算, 實際上部分親本有可能是來自全同胞家系, 從而影響遺傳力的估計準確性。全同胞效應其中包含了顯性效應和母本效應, 全同胞效應在魚類[10-11]中較小, 但在貝類幼體發育過程中, 存在較大的顯性效應和母本效應[12], 根據全同胞家系估計遺傳力時, 將全同胞效應當作加性效應一起估計會增大了遺傳力估計值,ASReml 軟件能較好的處理全同胞效應, 提高遺傳力的估計的準確性, 而試驗遺傳力估計的標準誤偏大則可能與測量樣本數量偏低有關[13]。
3.2 育種值
BLUP 法可較準確地估測育種值, 并在水產動物育種廣泛應用。孟乾[14]等利用BLUP 方法對長牡蠣“海大1 號”的育種值進行估計, 于50%選擇壓力下基于育種值家系選擇效率比基于表型值提高3%~34%, 個體選擇效率提高17%~28%。鄭靜靜[15]等利用BLUP 方法評估60、105 和150 日齡3 個階段中凡納濱對蝦(Litopenaeus vannamei)生長性狀的育種值, 于10%選擇壓力下基于育種值比基于表型值選擇效率提高20.06%、26.03%和12.01%。在本研究中, 于10%選擇壓力下, 試驗30 日齡施氏獺蛤殼長、殼高育種值選擇效率比其表型值選擇分別提高27%和21%, 80 日齡施氏獺蛤殼長、殼高、殼寬和體質量育種值選擇效率比其表型值選擇分別提高16%、14%、30%和13%。結果表明, 在施氏獺蛤育種中利用育種值選擇育種較表型值選擇育種具有更高的優勢。
3.3 生長性狀早期選擇的優劣
早期選擇的主要優勢是能夠早期篩查和選種,從而降低選育成本。施氏獺蛤為開放式底播養殖品種, 養殖群體易混合, 提高了選擇育種難度, 而其培育早期階段因處在人工控制條件下生長, 環境因素可控性較高, 比較容易獲取準確生長性狀表型值并加以分析和家系選擇, 可降低選育工作量。在不同選育策略下, 早期選擇存在相關性優勢, 如Gall 等[16]在羅非魚(Oreochromis mossambicus)改良育種研究分析認為在通過體長的對體質量的間接選育應在幼魚階段進行, 因后期該遺傳相關性降低, 本試驗中80 d 施氏獺蛤殼長、殼高與體質量的表型相關(0.94、0.93)和遺傳相關性(0.97、0.97)均較高, 這一現象有助于施氏獺蛤早期利用生長性狀相關進行綜合選育。貝類生長性狀發育策略并非一成不變[17-18], 不同階段性狀生長呈一致性是早期選擇的重要依據。本試驗前期發育快的家系在早期生長中也一直領先,與彭慧婧[19]等研究整個階段施氏獺蛤速生長家系發育情況相同, 表明對施氏獺蛤早期生長性狀的選擇具有較大可行性。
存活性狀也是影響貝類產量的重要性狀之一,早期選種可提前舍棄那些生長慢的家系或個體, 但無法對整個階段存活性狀進行預測。Langdon 等[20]對長牡蠣進行家系選育時, 發現一些家系雖然早期生長較快, 但生長至成貝時存活率較低, 閆喜武等[21]研究菲律賓蛤仔(Ruditapes philippinarum)家系早期生長發育發現稚貝期存活率趨于穩定且高于幼蟲期,不同階段存活性狀表現不同。本試驗中施氏獺蛤不同家系幼體附著后存活率差異較大(33%~74%), 而海上培育期存活率則相近(72%~91%), 且施氏獺蛤速生家系[19]在存活性狀方面也無明顯優勢。盡管生長性狀遺傳參數和育種值對施氏獺蛤選育參考作用明顯, 但在后續研究中還應綜合存活性狀對育種計劃的影響。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
