我國(guó)草地貪夜蛾年發(fā)生世代區(qū)劃分
2021-03-12 03:29:18謝殿杰唐繼洪張蕾程云霞江幸福
植物保護(hù) 2021年1期
謝殿杰 唐繼洪 張蕾 程云霞 江幸福


摘要 :為明確草地貪夜蛾在我國(guó)不同地區(qū)的年發(fā)生世代數(shù),為提高其預(yù)測(cè)預(yù)報(bào)和綜合防治水平提供科學(xué)依據(jù),本研究采用不同恒溫(20、24、28、32、36℃)條件,以玉米苗為幼蟲(chóng)食物測(cè)定了草地貪夜蛾不同發(fā)育階段及全世代的發(fā)育歷期,計(jì)算出草地貪夜蛾不同發(fā)育階段、全世代發(fā)育起點(diǎn)溫度和有效積溫。在此基礎(chǔ)上,結(jié)合1985年-2014年30年全國(guó)605個(gè)氣象站點(diǎn)的氣象數(shù)據(jù)預(yù)測(cè)了草地貪夜蛾在我國(guó)不同地區(qū)的年發(fā)生世代數(shù),并利用地理信息系統(tǒng)對(duì)其年發(fā)生世代區(qū)進(jìn)行了劃分。結(jié)果表明:草地貪夜蛾卵、1~6齡幼蟲(chóng)、蛹、雌成蟲(chóng)和雄成蟲(chóng)的發(fā)育起點(diǎn)溫度分別為11.20、9.63、13.03、12.88、11.09、13.04、10.73、13.90、13.98℃和16.68℃,有效積溫分別為41.32、41.32、18.71、28.23、45.87、60.33、55.90、90.97、166.86日·度和121.90日·度;全世代發(fā)育起點(diǎn)溫度為12.6℃,有效積溫為584.0日·度。根據(jù)全國(guó)不同地區(qū)氣象數(shù)據(jù)推測(cè)草地貪夜蛾在我國(guó)的理論年平均發(fā)生世代數(shù)在0~7.41代之間,總體上發(fā)生世代數(shù)隨緯度和海拔的升高而減少。我國(guó)1月份12.6℃等溫線以南地區(qū)全年最低溫度高于草地貪夜蛾發(fā)育起點(diǎn)溫度,為周年繁殖區(qū)域,主要位于云南南部(全年發(fā)生世代數(shù)最高可超過(guò)5代)、廣西南部、廣東中部以南、福建東南部(全年世代數(shù)最高可超過(guò)6代)、臺(tái)灣大部(全年世代數(shù)最高可超過(guò)7代)以及海南省全境(全年可發(fā)生7代以上)。江南和江淮遷飛過(guò)渡區(qū)主要為4~5代區(qū)和3~4代區(qū)。黃淮海及北方重點(diǎn)防范區(qū)主要為2~3代區(qū)和1~2代區(qū),部分為3~4代區(qū)。
關(guān)鍵詞 :草地貪夜蛾; 發(fā)生世代; 有效積溫; 發(fā)育起點(diǎn)溫度
中圖分類號(hào):
S 435.132文獻(xiàn)標(biāo)識(shí)碼: A
DOI: 10.16688/j.zwbh.2020213
Annual generation numbers prediction and division of fall armyworm,
Spodoptera frugiperda in China
XIE Dianjie1#, TANG Jihong2#, ZHANG Lei1, CHENG Yunxia1, JIANG Xingfu1*
(1. State Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection,
Chinese Academy of Agricultural Sciences, Beijing 100193, China; 2. Institute of Environment and
Plant Protection, Chinese Academy of Tropical Agricultural Sciences, Haikou 571101, China)
Abstract :In order to investigate the annual generation numbers of fall armyworm in different regions of China, and provide scientific basis for fall armyworm forecast and integrated control, the developmental durations of each developmental stage and whole generation of fall armyworm were determined at constant temperatures (20,24,28,32,36℃), and the effective accumulated temperature and the developmental threshold temperature were calculated. And then the annual generation numbers of fall armyworm in different regions of China were predicted based on the 30 years (1985-2014) meteorological data of 605 meteorological stations. Then the whole country was divided into eight regions according to the annual generation numbers of fall armyworm using geographic information system. The developmental threshold temperatures of eggs, 1st6th instar larvae, pupae, female and male adults were 11.20, 9.63, 13.03, 12.88, 11.09, 13.04, 10.73, 13.90, 13.98 and 16.68℃, respectively, and the effective accumulated temperatures were 41.32, 41.32, 18.71, 28.23, 45.87, 60.33, 55.90, 90.97, 166.86 d·℃ and 121.90 d·℃, respectively. The developmental threshold temperature and effective accumulated temperature of the whole generation were 12.6℃ and 584.0 d·℃, respectively. The theoretical average annual generation numbers of fall armyworm ranged from 0 to 7.41. The theoretical average annual generation numbers decreased with the increase of latitude and altitude. In January, the annual minimum temperature in the south of 12.6℃ isotherm was above 12.6℃, which was higher than the developmental threshold temperature of fall armyworm, and the fall armyworm could develop normally all year round. These areas include southern of Yunnan (annual generation numbers can exceed five generations at most), southern of Guangxi, south of Guangdong, southeast of Fujian (annual generation numbers can exceed six generations at most), most of Taiwan (annual generation numbers can exceed seven generations at most) and all of Hainan province (more than seven generation numbers throughout the year). The fall armyworm could occur four to five generations and three to four generations in migration transition zones of Jiangnan and Jianghuai areas. In Huanghuaihai and the key prevention areas (northern China), the fall armyworm could occur two to three or one to two generations in most areas, and three to four generations in some areas.
Key words :Spodoptera frugiperda; generation number; effective accumulated temperature; developmental threshold temperature
草地貪夜蛾Spodoptera frugiperda (J.E.Smith)屬鱗翅目Lepidoptera,夜蛾科Noctuidae,灰翅夜蛾屬Spodoptera,原產(chǎn)于美洲熱帶和亞熱帶地區(qū)[1]。草地貪夜蛾寄主范圍廣,在美洲可為害76科227屬的353種自然寄主植物[2];為害嚴(yán)重時(shí)可造成玉米減產(chǎn)20%~40%[3];其繁殖能力強(qiáng),單雌平均產(chǎn)卵量可達(dá)1 449粒[4];對(duì)溫度適應(yīng)性強(qiáng),在中、南美洲熱帶和亞熱帶地區(qū)可周年發(fā)生[56];成蟲(chóng)具有遷飛習(xí)性,在中、南美洲可向南遷飛至阿根廷,向北遷飛至加拿大,氣象條件適宜的情況下,草地貪夜蛾從美國(guó)南部密西西比州遷飛至加拿大南部,共1 600 km的距離僅需30 h[710]。2016年1月,草地貪夜蛾首次入侵非洲,并迅速蔓延到非洲大部分地區(qū)[11]以及亞洲部分國(guó)家[12],成為一種世界性農(nóng)業(yè)害蟲(chóng)。2019年1月,草地貪夜蛾入侵我國(guó)云南,隨后迅速蔓延,到2019年10月,有26個(gè)省(市、區(qū))監(jiān)測(cè)到草地貪夜蛾成蟲(chóng),有22個(gè)省(市、區(qū))查見(jiàn)幼蟲(chóng)為害[1315]。
溫度、濕度、光周期等影響著昆蟲(chóng)的生長(zhǎng)發(fā)育及分布范圍,其中,溫度是最主要的環(huán)境因素[16]。昆蟲(chóng)完成某一發(fā)育階段必須積累一定量的有效溫度,即有效積溫(effective accumulated temperature)。當(dāng)環(huán)境溫度高于昆蟲(chóng)的發(fā)育起點(diǎn)溫度(developmental threshold temperature)時(shí),昆蟲(chóng)才開(kāi)始發(fā)育。昆蟲(chóng)的發(fā)育起點(diǎn)溫度及有效積溫決定了它是否能在某一地區(qū)完成生活史[1719]。因此,研究昆蟲(chóng)的發(fā)育起點(diǎn)溫度和有效積溫有助于預(yù)測(cè)其在某一地區(qū)的發(fā)生期和發(fā)生世代數(shù),為害蟲(chóng)的預(yù)測(cè)預(yù)報(bào)及綜合治理提供依據(jù)。
有關(guān)草地貪夜蛾發(fā)育起點(diǎn)溫度、有效積溫和發(fā)生世代的研究國(guó)內(nèi)已有少量報(bào)道,如何莉梅等研究了草地貪夜蛾的發(fā)育起點(diǎn)溫度和有效積溫[20];張紅梅等根據(jù)試驗(yàn)測(cè)定的草地貪夜蛾發(fā)育起點(diǎn)溫度和有效積溫推測(cè)了其在云南各地區(qū)的年發(fā)生世代數(shù)[21]。秦譽(yù)嘉等、盧輝等預(yù)測(cè)了草地貪夜蛾在我國(guó)的潛在地理分布[2223]。這些結(jié)果對(duì)于闡明草地貪夜蛾在我國(guó)的發(fā)生規(guī)律及提高預(yù)測(cè)預(yù)報(bào)水平有重要的作用,但由于不同作者采用的飼養(yǎng)條件等不同,得到的草地貪夜蛾發(fā)育起點(diǎn)溫度和有效積溫?cái)?shù)據(jù)差異較大,而且目前關(guān)于草地貪夜蛾在我國(guó)不同地區(qū)發(fā)生世代數(shù)的研究尚未見(jiàn)報(bào)道。
鑒于草地貪夜蛾入侵后在我國(guó)迅速蔓延,已經(jīng)對(duì)我國(guó)糧食安全產(chǎn)生了巨大威脅[2425],為摸清其在我國(guó)的發(fā)生規(guī)律,提高其預(yù)測(cè)預(yù)報(bào)和綜合防治水平。本研究在室內(nèi)5種恒溫條件下以玉米苗為幼蟲(chóng)食物,系統(tǒng)測(cè)定草地貪夜蛾不同發(fā)育階段的發(fā)育歷期,計(jì)算出不同發(fā)育階段及全世代發(fā)育起點(diǎn)溫度,在此基礎(chǔ)上,利用1985年-2014年我國(guó)不同地區(qū)605個(gè)氣象站的氣象數(shù)據(jù),并結(jié)合地理信息系統(tǒng)按照預(yù)測(cè)的草地貪夜蛾在各地區(qū)年發(fā)生世代數(shù)對(duì)其發(fā)生區(qū)進(jìn)行了劃分,從而為提高草地貪夜蛾的預(yù)測(cè)預(yù)報(bào)和綜合治理水平提供科學(xué)依據(jù)。
1 材料與方法
1.1 供試蟲(chóng)源及種群飼養(yǎng)技術(shù)
試驗(yàn)用草地貪夜蛾幼蟲(chóng)于2019年4月采自廣西壯族自治區(qū)南寧市(108.37°E,22.82°N),帶回室內(nèi)人工飼養(yǎng)繁殖,建立種群。種群飼養(yǎng)技術(shù)同謝殿杰等的方法[4]。
1.2 草地貪夜蛾不同溫度下飼養(yǎng)與發(fā)育歷期觀察
試驗(yàn)在恒溫培養(yǎng)箱中進(jìn)行,設(shè)置20、24、28、32、36℃共5個(gè)溫度梯度,相對(duì)濕度為(60±5)%、光周期L∥D=16 h∥8 h、光照強(qiáng)度為18 000 lx。幼蟲(chóng)用新鮮玉米苗飼喂,每天更換,直至幼蟲(chóng)化蛹,記錄幼蟲(chóng)歷期及蛹?xì)v期。成蟲(chóng)羽化后記錄其存活歷期,直至成蟲(chóng)全部死亡。
1.3 發(fā)育起點(diǎn)溫度及有效積溫的計(jì)算
試驗(yàn)中不同溫度下草地貪夜蛾世代周期的平均數(shù)和標(biāo)準(zhǔn)誤用SPSS 20軟件計(jì)算,并進(jìn)行方差分析,使用Tukeys HSD法進(jìn)行差異顯著性檢驗(yàn)。根據(jù)有效積溫法則,采用最小二乘法計(jì)算草地貪夜蛾整個(gè)世代的發(fā)育起點(diǎn)溫度(C)和有效積溫(K)。
C=∑V2∑T-∑V∑VTn∑V2-∑V2(1)
K=n∑VT-∑V∑Tn∑V2-∑V2(2)
Sc= ∑(T-T′)2n-21n+2∑(V-)2(3)
Sk= ∑(T-T′)2(n-2)∑(V-)2(4)
其中,V為發(fā)育速率(發(fā)育歷期的倒數(shù)),T為處理試驗(yàn)溫度,T′為理論溫度,T′=C+KV,為發(fā)育速率的平均值,n為試驗(yàn)溫度處理數(shù)(n=5),C為發(fā)育起點(diǎn)溫度,K為有效積溫,Sc和Sk分別是C和K的標(biāo)準(zhǔn)差。
1.4 氣象數(shù)據(jù)來(lái)源與處理
本文所用1985年-2014年氣象數(shù)據(jù)均下載于國(guó)家氣象科學(xué)數(shù)據(jù)中心網(wǎng)站(http:∥data.cma.cn/data/cdcdetail/dataCode/SURF_CLI_CHN_MUL_DAY_V3.0.html);所用氣象數(shù)據(jù)集為中國(guó)地面氣候資料日值數(shù)據(jù)集(V3.0)。在去除數(shù)據(jù)不完整的站點(diǎn)后,得到605個(gè)氣象站點(diǎn)數(shù)據(jù)。
1.5 草地貪夜蛾理論年發(fā)生世代數(shù)計(jì)算
根據(jù)計(jì)算出的草地貪夜蛾發(fā)育起點(diǎn)溫度和有效積溫,按照公式(5)計(jì)算1.4所述605個(gè)氣象站點(diǎn)1985年-2014年每年草地貪夜蛾理論年發(fā)生世代數(shù),并計(jì)算每個(gè)站點(diǎn)這30年理論年發(fā)生世代數(shù)平均值,將其作為對(duì)應(yīng)氣象站點(diǎn)草地貪夜蛾理論年發(fā)生世代數(shù)。從代表性省市隨機(jī)抽出1個(gè)氣象站點(diǎn),將該站點(diǎn)草地貪夜蛾理論年發(fā)生世代數(shù)作為驗(yàn)證數(shù)據(jù)(理論計(jì)算值)不參與模型構(gòu)建(共抽取29個(gè)氣象站點(diǎn)),其余氣象站點(diǎn)數(shù)據(jù)輸入地理信息系統(tǒng)軟件ArcGIS 10.6,利用地統(tǒng)計(jì)分析模塊對(duì)草地貪夜蛾年發(fā)生世代數(shù)進(jìn)行Kriging插值分析,得到全國(guó)草地貪夜蛾年發(fā)生世代數(shù)分布圖;并利用數(shù)值提取工具(GA Layer to Points)將驗(yàn)證數(shù)據(jù)所對(duì)應(yīng)的擬合年發(fā)生世代數(shù)(擬合值)從該圖中提取出來(lái),利用公式6所示卡方檢驗(yàn)分析比較兩者的差異,從而判斷所得年發(fā)生世代數(shù)分布圖的優(yōu)劣。
G=∑ni=0(Ti-C)K
(5)
其中 Ti>C;當(dāng)Ti≤C時(shí),Ti-C=0;式中,G為年發(fā)生世代數(shù),n為全年的天數(shù),Ti為全年中第i日平均氣溫,C為發(fā)育起點(diǎn)溫度,K為世代有效積溫。
SymbolcA@2=∑ki=1(Oi-Ei)2Ei(6)
其中,Oi為第i個(gè)觀測(cè)值(也即是本文中的理論計(jì)算值);Ei為擬合值。如果卡方值SymbolcA@2小于自由度為k-1、α=0.05顯著性水平的臨界值SymbolcA@2(df=k-1,α=0.05),則表示擬合值和理論計(jì)算值無(wú)顯著差異,擬合值能較好地代表理論計(jì)算值。
1.6 1月份12.6℃等溫線提取
由1.5所述605個(gè)氣象站點(diǎn)氣候資料日值數(shù)據(jù)計(jì)算得出1985年-2014年1月份平均氣溫平均值,將該數(shù)據(jù)輸入地理信息系統(tǒng)軟件ArcGIS 10.6,經(jīng)過(guò)Kriging插值分析后得到全國(guó)1月份平均氣溫分布圖,再利用等值線提取工具(GA Layer to Contour)提取12.6℃全世代發(fā)育起點(diǎn)溫度等溫線。
2 結(jié)果與分析
2.1 草地貪夜蛾的世代周期及有效積溫、發(fā)育起點(diǎn)溫度
利用有效積溫和發(fā)育起點(diǎn)溫度的計(jì)算公式得到草地貪夜蛾卵、1~6齡幼蟲(chóng)、蛹、雌成蟲(chóng)和雄成蟲(chóng)的發(fā)育起點(diǎn)溫度分別為11.20、9.63、13.03、12.88、11.09、13.04、10.73、13.90、13.98℃和16.68℃,有效積溫分別為41.32、41.32、18.71、28.23、45.87、60.33、55.90、90.97、166.86日·度和121.90日·度;整個(gè)世代的發(fā)育起點(diǎn)溫度為12.6℃,有效積溫為584.0日·度(表1)。
2.2 草地貪夜蛾理論年發(fā)生世代數(shù)
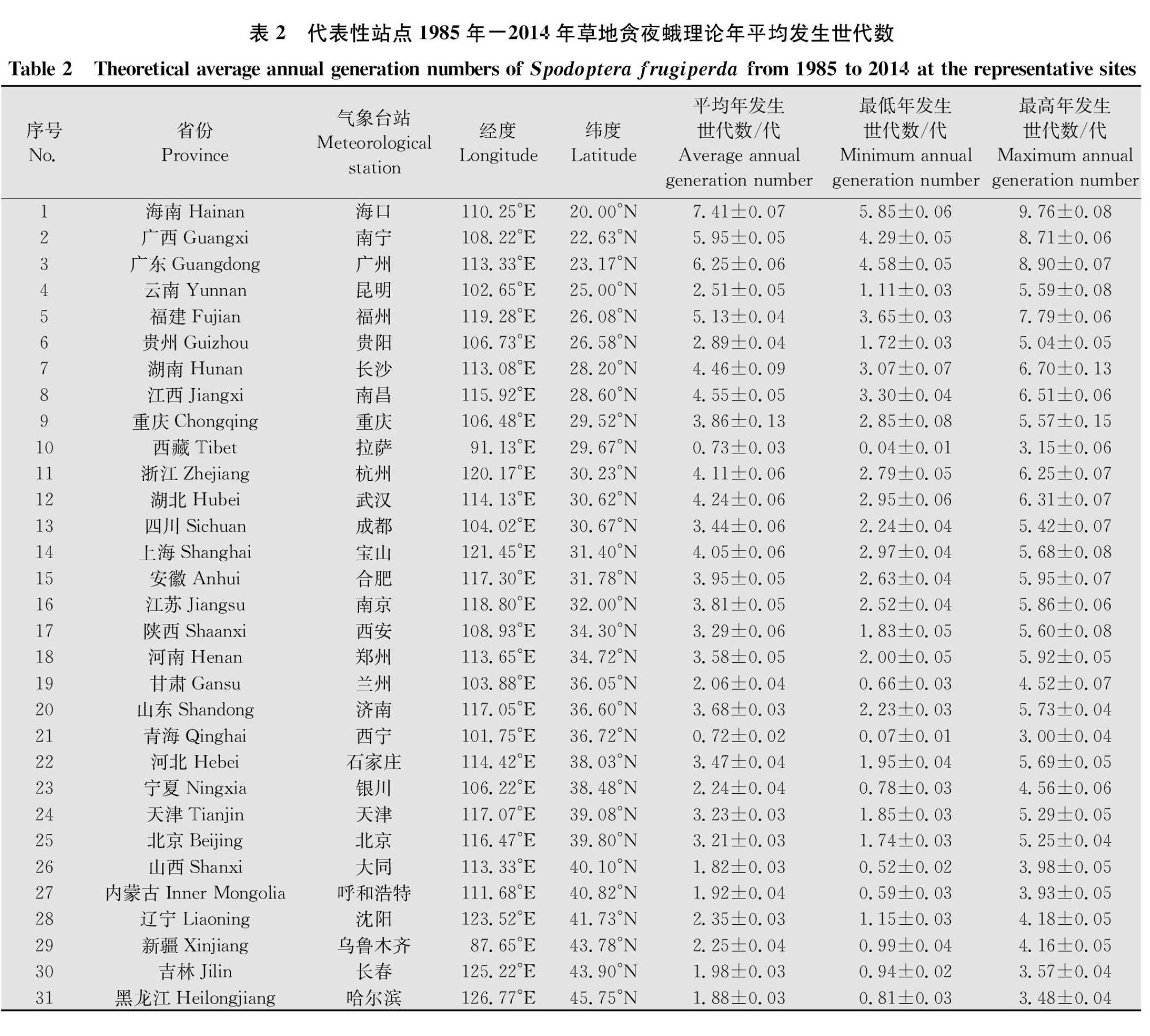
通過(guò)對(duì)605個(gè)氣象站點(diǎn)1985年-2014年的氣象數(shù)據(jù)及草地貪夜蛾的發(fā)育起點(diǎn)溫度和有效積溫計(jì)算得到各站點(diǎn)草地貪夜蛾理論年發(fā)生世代數(shù)。表2為31個(gè)代表性站點(diǎn)1985年-2014年草地貪夜蛾理論平均年發(fā)生世代數(shù),范圍在0~7.41代,理論平均年發(fā)生世代數(shù)隨緯度的升高有減小的趨勢(shì),然而如西藏拉薩、青海西寧等地由于海拔較高,氣溫相對(duì)同緯度其他地區(qū)低,理論平均年發(fā)生世代數(shù)也相對(duì)較少。如表3所示,隨機(jī)選取的作為驗(yàn)證數(shù)據(jù)的29個(gè)氣象站點(diǎn)草地貪夜蛾理論年平均發(fā)生世代數(shù)(理論計(jì)算值)和擬合值無(wú)顯著差異(SymbolcA@2=2.956 0 不足1代區(qū):包括黑龍江西北部及內(nèi)蒙古東北部,青藏高原西藏和青海大部,四川西北部等區(qū)域。 1~2代區(qū):包括黑龍江、吉林大部分區(qū)域,內(nèi)蒙古中東部,河北西北部,新疆東北部,甘肅、寧夏大部分區(qū)域,西藏、青海北部,四川中西部,云南北部等區(qū)域。 2~3代區(qū):包括內(nèi)蒙古、吉林和遼寧三省交界處(包括遼寧大部分區(qū)域),河北大部,北京,天津,山東中部及膠州半島,山西、陜西大部,內(nèi)蒙古中西部,新疆大部,云南、四川及貴州三省交界處等區(qū)域。 3~4代區(qū):包括新疆中部,河北東南部,山東西南部,河南大部,安徽,江蘇,浙江西北部,湖北大部,重慶,四川東部,湖南西北部,貴州大部,云南中部等區(qū)域。 4~5代區(qū):包括湖北東南部,湖南大部,江西大部,浙江大部,福建大部,臺(tái)灣北部,云南中南部,廣西北部等區(qū)域。 5~6代區(qū):包括云南南部,廣西中部,廣東中北部,福建東南部,臺(tái)灣大部等區(qū)域。 6~7代區(qū):包括廣西南部,廣東南部及臺(tái)灣南部等區(qū)域。 7代以上區(qū):海南省。 2.3 1月份12.6℃等溫線 如圖1所示,1月份12.6℃等溫線位于云南省中南部、廣西南部、廣東中部、福建東南部及臺(tái)灣北部。該等溫線在云南省處于4~5代區(qū)內(nèi),在廣西、廣東及福建三省位于5~6代區(qū)內(nèi),而在臺(tái)灣則位于4~5代區(qū)及5~6代區(qū)內(nèi)。該等溫線以南的區(qū)域1月份平均氣溫均在12.6℃以上,高于草地貪夜蛾的全世代發(fā)育起點(diǎn)溫度,草地貪夜蛾可以正常發(fā)育,可能成為草地貪夜蛾的周年繁殖區(qū)域。 3 討論 在昆蟲(chóng)的生長(zhǎng)發(fā)育過(guò)程中,當(dāng)溫度高于其發(fā)育起點(diǎn)溫度時(shí)昆蟲(chóng)才開(kāi)始發(fā)育,并且完成某一發(fā)育階段所需的有效積溫是固定的,因此,外界環(huán)境溫度的高低決定了昆蟲(chóng)是否能夠完成世代發(fā)育以及生長(zhǎng)發(fā)育的快慢[2627]。已有研究表明,草地貪夜蛾的發(fā)育歷期隨溫度降低而顯著延長(zhǎng)[4]。本研究測(cè)定了不同溫度下草地貪夜蛾的發(fā)育歷期,并計(jì)算得出其整個(gè)世代的發(fā)育起點(diǎn)溫度為12.6℃,有效積溫為584.0日·度。Schlemmer[28]曾報(bào)道草地貪夜蛾的發(fā)育起點(diǎn)溫度為12.57℃,有效積溫為391.01日·度;何莉梅等[20]研究表明,草地貪夜蛾的世代發(fā)育起點(diǎn)溫度為9.16℃,有效積溫為680.02日·度。也有研究表明草地貪夜蛾發(fā)育起點(diǎn)溫度為10.9℃,有效積溫需559日·度[29]。這些研究結(jié)果存在一定的差異,可能是由于試驗(yàn)所設(shè)置的溫度、濕度、光照及食料不完全一致,所使用的儀器、數(shù)據(jù)采集和統(tǒng)計(jì)分析方法等也有所不同造成的。例如,Schlemmer利用幼嫩甜玉米棒飼喂幼蟲(chóng),所設(shè)置的光周期為L(zhǎng)∥D=14 h∥10 h;何莉梅等利用人工飼料飼喂幼蟲(chóng)。而本試驗(yàn)考慮到玉米苗期階段受害較重,心葉和葉片都有被害,所以利用新鮮玉米苗飼喂幼蟲(chóng),更接近實(shí)際情況。另外,由于全球氣候的變化,可能使不同地理種群的草地貪夜蛾生物學(xué)特性不同,也會(huì)對(duì)試驗(yàn)結(jié)果產(chǎn)生影響。 張紅梅等利用發(fā)育起點(diǎn)溫度和有效積溫對(duì)草地貪夜蛾在云南的年發(fā)生世代數(shù)進(jìn)行了預(yù)測(cè)[21],但是沒(méi)有涉及草地貪夜蛾發(fā)生的其他地區(qū)。姜玉英等[30]根據(jù)何莉梅等[20]的研究結(jié)果進(jìn)行分析,發(fā)現(xiàn)草地貪夜蛾全世代有效積溫與黏蟲(chóng)接近,并用黏蟲(chóng)在我國(guó)不同緯度的發(fā)生世代數(shù)對(duì)草地貪夜蛾可能的年發(fā)生世代數(shù)進(jìn)行了推測(cè)。本研究表明,理論上草地貪夜蛾在我國(guó)年平均發(fā)生世代數(shù)為0~7.41代。不足1代區(qū)主要是黑龍江西北部及內(nèi)蒙古東北部、青藏高原西藏和青海大部、四川西北部等區(qū)域,主要原因是這些地區(qū)緯度或海拔較高,一年中大部分時(shí)間氣溫都在草地貪夜蛾的世代發(fā)育起點(diǎn)溫度以下;1月份12.6℃等溫線位于云南省中南部、廣西南部、廣東中部、福建東南部及臺(tái)灣北部。該等溫線以南的區(qū)域1月份平均氣溫均在12.6℃以上,全年均高于草地貪夜蛾的世代發(fā)育起點(diǎn)溫度,在這些地區(qū)草地貪夜蛾可以正常發(fā)育,為草地貪夜蛾的周年繁殖區(qū)域。黏蟲(chóng)的發(fā)育起點(diǎn)溫度為9.6℃,全世代有效積溫為685.2日·度[31],在我國(guó)各區(qū)域的發(fā)生代次為:華南(27°N以南) 6~8代,中南(27°N~33°N)5~6代,江淮(33°N~36°N)4~5代,華北(36°N~39°N) 3~4代和東北(39°N以北) 2~3代[33]。與黏蟲(chóng)相比,草地貪夜蛾在各區(qū)域的發(fā)生代數(shù)相對(duì)較低,這可能由于草地貪夜蛾的全世代發(fā)育起點(diǎn)溫度(12.6℃)高于黏蟲(chóng)的全世代發(fā)育起點(diǎn)溫度,其需要較高的環(huán)境溫度才能開(kāi)始生長(zhǎng)發(fā)育,因此全年發(fā)生總世代數(shù)相對(duì)減少。 2019年草地貪夜蛾已經(jīng)侵入我國(guó)除青海、新疆、遼寧、吉林、黑龍江等5省以外的所有區(qū)域,專家預(yù)測(cè)2020年其為害范圍還將進(jìn)一步擴(kuò)大。云南、廣東、廣西、海南、福建等地為草地貪夜蛾的周年繁殖區(qū)[30],這與本研究預(yù)測(cè)的分布范圍及周年繁殖區(qū)域的劃分基本一致。云南、廣西、廣東、海南等地玉米周年種植,特別是該區(qū)域冬玉米的種植,為該蟲(chóng)的周年發(fā)生提供了充足的食物來(lái)源,也為該蟲(chóng)向長(zhǎng)江及淮河流域的遷飛提供了大量蟲(chóng)源。同時(shí),我國(guó)玉米等作物的種植布局隨季節(jié)和緯度變化從南至北遞次推移,時(shí)間和空間上互補(bǔ)的食物資源可促使草地貪夜蛾種群區(qū)域性遷移為害[3334]。隨著草地貪夜蛾在我國(guó)的定殖,該蟲(chóng)可能會(huì)形成與黏蟲(chóng)類似的季節(jié)性往返遷飛規(guī)律,將會(huì)嚴(yán)重威脅我國(guó)的糧食安全;并且草地貪夜蛾大齡幼蟲(chóng)具有捕食習(xí)性,可攻擊取食其他害蟲(chóng)和部分天敵,在生態(tài)位競(jìng)爭(zhēng)中占據(jù)較大的優(yōu)勢(shì)[35],會(huì)對(duì)農(nóng)田生態(tài)系統(tǒng)造成嚴(yán)重的破壞。 值得一提的是,本研究中草地貪夜蛾全國(guó)發(fā)生世代數(shù)的計(jì)算與劃分是基于分布在全國(guó)的605個(gè)區(qū)域級(jí)氣象站點(diǎn)的歷史氣象數(shù)據(jù),并利用ArcGIS軟件的插值功能對(duì)氣象站點(diǎn)以外的區(qū)域進(jìn)行插值而得到的。由于這605個(gè)氣象站點(diǎn)的分布并不均勻,中東部區(qū)域分布較多,而西部如西藏、新疆等地分布相對(duì)較少,這樣計(jì)算出來(lái)的數(shù)值可能和實(shí)際情況存在一些差異,氣象站點(diǎn)較為密集的地域精度相對(duì)較高,而站點(diǎn)少的地域精度相對(duì)偏低。本研究中年發(fā)生世代數(shù)是根據(jù)30年的氣象數(shù)據(jù)計(jì)算得到的平均值,受全球氣候變化的影響,氣溫年際間存在差異,可能會(huì)與將來(lái)田間實(shí)際調(diào)查觀測(cè)到的世代發(fā)生情況產(chǎn)生一定的出入。此外,插值模型本身局限性等因素也可能會(huì)對(duì)田間實(shí)際發(fā)生情況一致性產(chǎn)生影響。盡管如此,本研究結(jié)果將會(huì)為摸清草地貪夜蛾田間發(fā)生規(guī)律以及提高其預(yù)測(cè)預(yù)報(bào)及田間防治水平提供重要依據(jù)。 最后,本研究蟲(chóng)源飼養(yǎng)在恒溫條件下,而自然環(huán)境中的溫度是時(shí)刻發(fā)生改變的,今后有必要進(jìn)一步研究自然變溫條件下草地貪夜蛾世代的發(fā)育起點(diǎn)溫度和有效積溫;寄主植物在不同區(qū)域的分布情況也會(huì)對(duì)草地貪夜蛾的發(fā)生世代數(shù)產(chǎn)生影響;同時(shí)還應(yīng)綜合考慮降水、濕度、光周期等其他環(huán)境因子的影響。因此,下一步應(yīng)加強(qiáng)田間草地貪夜蛾發(fā)生世代數(shù)的調(diào)查工作,結(jié)合理論預(yù)測(cè)結(jié)果,為草地貪夜蛾的精準(zhǔn)預(yù)測(cè)預(yù)報(bào)及田間防治提供科學(xué)依據(jù)。 參考文獻(xiàn) [1] TODD E L, POOLE R W. Keys and illustrations for the armyworm moths of the noctuid genus Spodoptera Guenée from the Western Hemisphere [J]. Annals of the Entomological Society of America, 1980, 73(6): 722738. [2] MONTEZANO D G, SPECHT A, BORTOLIN T M, et al. Immature stages of Spodoptera albula (Walker) (Lepidoptera: Noctuidae): Developmental parameters and host plants[J]. Anais da Academia Brasileira de Ciências, 2013, 85(1): 271284. [3] WYCKHUYS K A G, ONEIL R J. Population dynamics of Spodoptera frugiperda Smith (Lepidoptera: Noctuidae) and associated arthropod natural enemies in Honduran subsistence maize [J]. Crop Protection, 2006, 25(11): 11801190. [4] 謝殿杰, 張蕾, 程云霞, 等. 不同溫度下草地貪夜蛾年齡階段實(shí)驗(yàn)種群兩性生命表的構(gòu)建[J]. 植物保護(hù), 2019, 45(6): 2027. [5] MURA G, MOLINAOCHOA J, COVIELLA C. Population dynamics of the fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae) and its parasitoids in northwestern Argentina [J]. Florida Entomologist, 2006, 89(2): 175183. [6] LUGINBILL P. The fall armyworm [R]. USDA Technology Bulletin, 1928, 34: 91. [7] ROSE A H, SILVERSIDES R H, LINDQUIST O H. Migration flight by an aphid,Rhopalosiphum maidis (Hemiptera: Aphididae), and a noctuid, Spodoptera frugiperda (Lepidoptera: Noctuidae) [J]. The Canadian Entomologist, 1975, 107(6): 567576. [8] SPARKS A N. A review of the biology of the fall armyworm [J]. Florida Entomologist, 1979, 62(2): 8287. [9] JOHNSON C G. Migration and dispersal of insects by flight [M]. London: Methuen, 1969: 745763. [10]JOHNSON S J. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the Western Hemisphere [J]. International Journal of Tropical Insect Science, 1987, 8(4/5/6): 543549. [11]DAY R, ABRAHAMS P, BATEMAN M, et al.Fall armyworm: impacts and implications for Africa [J]. Outlooks on Pest Management, 2017, 28(5): 196201. [12]劉剛. 全國(guó)農(nóng)技中心通報(bào) 我國(guó)及周邊國(guó)家草地貪夜蛾發(fā)生為害情況[J]. 農(nóng)藥市場(chǎng)信息, 2019(9): 59. [13]楊學(xué)禮, 劉永昌, 羅茗鐘, 等. 云南省江城縣首次發(fā)現(xiàn)遷入我國(guó)西南地區(qū)的草地貪夜蛾[J]. 云南農(nóng)業(yè), 2019(1): 72. [14]吳秋琳, 姜玉英, 吳孔明. 草地貪夜蛾緬甸蟲(chóng)源遷入中國(guó)的路徑分析[J]. 植物保護(hù), 2019, 45(2): 16. [15]郭井菲, 趙建周, 何康來(lái), 等. 警惕危險(xiǎn)性害蟲(chóng)草地貪夜蛾入侵中國(guó)[J]. 植物保護(hù), 2018, 44(6): 110. [16]BROUFAS G D, PAPPAS M L, KOVEOS D S. Effect of relative humidity on longevity, ovarian maturation, and egg production in the olive fruit fly (Diptera: Tephritidae) [J]. Annals of the Entomological Society of America, 2009, 102(1): 7075. [17]RIDGWAY R L, GYRISCO GG. Effect of temperature on the rate of development of Lygus lineolaris (Hemiptera: Miridae) [J]. Annals of the Entomological Society of America, 1960, 53(5): 691694. [18]仵均祥. 農(nóng)業(yè)昆蟲(chóng)學(xué)[M].北京: 中國(guó)農(nóng)業(yè)出版社, 2009: 23. [19]BARTEKOVA A, PRASLICKA J.The effect of ambient temperature on the development of cotton bollworm (Helicoverpa armigera Hübner, 1808) [J]. Plant Protection Science, 2016, 42: 135138. [20]何莉梅, 葛世帥, 陳玉超, 等. 草地貪夜蛾的發(fā)育起點(diǎn)溫度、有效積溫和發(fā)育歷期預(yù)測(cè)模型[J]. 植物保護(hù), 2019, 45(5): 1826. [21]張紅梅, 尹艷瓊, 趙雪晴, 等. 草地貪夜蛾在不同溫度條件下的生長(zhǎng)發(fā)育特性[J]. 環(huán)境昆蟲(chóng)學(xué)報(bào), 2020, 42(1): 5259. [22]秦譽(yù)嘉, 藍(lán)帥, 趙紫華, 等. 遷飛性害蟲(chóng)草地貪夜蛾在我國(guó)的潛在地理分布[J]. 植物保護(hù), 2019, 45(4): 4347. [23]盧輝, 唐繼洪, 呂寶乾, 等.草地貪夜蛾的生物防治及潛在入侵風(fēng)險(xiǎn)[J]. 熱帶作物學(xué)報(bào), 2019, 40(6): 12371244. [24]葛世帥, 何莉梅, 和偉, 等. 草地貪夜蛾的飛行能力測(cè)定[J]. 植物保護(hù), 2019, 45(4): 2833. [25]郭井菲, 何康來(lái), 王振營(yíng). 草地貪夜蛾的生物學(xué)特性、發(fā)展趨勢(shì)及防控對(duì)策[J]. 應(yīng)用昆蟲(chóng)學(xué)報(bào), 2019, 56(3): 361369. [26]HOFFMANNA A, SRENSEN J G, LOESCHCKE V. Adaptation of Drosophila to temperature extremes: bringing together quantitative and molecular approaches [J]. Journal of Thermal Biology, 2003, 28(3): 175216. [27]CUI Xuhong, WAN Fanghao, XIE Ming, et al. Effects of heat shock on survival and reproduction of two whitefly species, Trialeurodes vaporariorum and Bemisia tabaci biotype B [J]. Journal of Insect Science, 2008, 8(24): 110. [28]SCHLEMMER M.Effect of temperature on development and reproduction of Spodoptera frugiperda (Lepidoptera: Noctuidae) [D]. Evanston: NorthWest University, 2018. [29]RAMIREZ G, BRAVO M, LLANDERAL C.Development of Spodoptera frugiperda (J. E. Smith)(Lepidoptera: Noctuidae) under different conditions of temperature and humidity [J]. Agrociencia, 1987(67): 161171. [30]姜玉英, 劉杰, 謝茂昌, 等. 2019年我國(guó)草地貪夜蛾擴(kuò)散為害規(guī)律觀測(cè)[J]. 植物保護(hù), 2019, 45(6): 1019. [31]林昌善. 粘蟲(chóng)生理生態(tài)學(xué)[M]. 北京: 北京大學(xué)出版社, 1990: 518526. [32]李光博, 王恒祥, 胡文繡. 粘蟲(chóng)季節(jié)性遷飛為害假說(shuō)及標(biāo)記回收試驗(yàn)[J]. 植物保護(hù)學(xué)報(bào), 1964, 3(2): 101110. [33]楊普云, 朱曉明, 郭井菲, 等. 我國(guó)草地貪夜蛾的防控對(duì)策與建議[J]. 植物保護(hù), 2019, 45(4): 16. [34]吳秋琳, 姜玉英, 胡高, 等. 中國(guó)熱帶和南亞熱帶地區(qū)草地貪夜蛾春夏兩季遷飛軌跡的分析[J]. 植物保護(hù), 2019, 45(3): 19. [35]吳孔明. 中國(guó)草地貪夜蛾的防控策略[J]. 植物保護(hù), 2020, 46(2): 15. (責(zé)任編輯:田 喆)
