短時高低溫脅迫對南亞實蠅生長發育及繁殖的影響
2021-03-12 03:29:18黃禹禹顧祥鵬彭孝琴陳國華齊國君張曉明陶玫
植物保護 2021年1期
黃禹禹 顧祥鵬 彭孝琴 陳國華 齊國君 張曉明 陶玫
摘要 :南亞實蠅Bactrocera tau (Walker)是一種重要的入侵性農業害蟲。主要分布在我國云南、廣東、四川和河南等地,南亞實蠅的為害嚴重降低了蔬菜和水果的品質。短時極端溫度是影響南亞實蠅種群的重要因子。為了明確短時高低溫對南亞實蠅的影響,通過選擇12 h的短時低溫(-4、0、4、8℃)及12 h的短時高溫(34、38、42、46℃),在實驗室條件下系統研究短時高低溫脅迫對南亞實蠅種群生長發育和繁殖的影響。結果表明,在短時高/低溫處理后,南亞實蠅各蟲態的存活率隨著處理溫度的升高/降低逐漸下降;隨著處理短時高/低溫的升高/降低,南亞實蠅發育歷期、產卵前期和后代性比逐漸增加;隨著短時高溫處理溫度的升高,雌雄成蟲的壽命先延長后縮短,單雌產卵量逐漸減少;隨著短時低溫處理溫度的降低,雌雄成蟲的壽命逐漸縮短,單雌產卵量先增加后減少。短時高溫和短時低溫均對南亞實蠅的生長發育和繁殖有著不利的影響,但一定的低溫(8℃)處理后對雌成蟲繁殖力有一定的促進作用。
關鍵詞 :南亞實蠅; 高溫; 低溫; 生長發育; 繁殖
中圖分類號:
S 433文獻標識碼: A
DOI: 10.16688/j.zwbh.2020113
Effects of shortterm high and low temperature on the development and reproduction of Bactrocera tau (Walker)
HUANG Yuyu1, GU Xiangpeng1, PENG Xiaoqin1, CHEN Guohua1,QI Guojun2, ZHANG Xiaoming1*, TAO Mei1*
(1. College of Plant Protection, Yunnan Agricultural University, National Key Laboratory for Conservation and
Utilization of Biological Resources in Yunnan, Kunming 650201, China; 2. Guangdong Provincial Key
Laboratory of High Technology for Plant Protection, Research Institute of Plant Protection,
Guangdong Academy of Agricultural Sciences, Guangzhou 510640, China)
Abstract :Bactrocera tau (Walker) is an important agricultural invasive insect pest. It is mainly distributed in Yunnan, Guangdong, Sichuan and Henan provinces of China. The serious damage of B.tau can reduce the quality of vegetables and fruits. The shortterm extreme temperature is an important factor affecting the population of B.tau. In order to determine the effect of shortterm high and low temperature on the growth, development and reproduction of B.tau, the effects of shortterm low temperature (-4, 0, 4, 8℃) for 12 hours and shortterm high temperature (34, 38, 42, 46℃) for 12 hours were studied under laboratory conditions. The results showed that the survival rates of all stages gradually decreased with the increase and decrease of the treatment temperature after shortterm high and low temperature treatments. The developmental period, preoviposition period, single female oviposition, longevity and the sex ratio of offspring were significantly affected by the treatments. The developmental period, sex ratio and preoviposition period gradually increased with increasing/decreasing temperature; the longevity of male and female adults initially increased and then decreased, while the single female oviposition gradually decreased with increasing treatment temperature. Meanwhile, their longevity gradually decreased and the single female oviposition increased and then decreased with decreasing treatment temperature. Overall, the shortterm high and low temperature had negative effects on the growth, development and reproduction of B.tau. This could slightly promote the reproduction of females in a certain range of shortterm low temperature (8℃).
Key words :Bactrocera tau; high temperature; low temperature; growth; development; reproduction
南亞實蠅Bactrocera tau (Walker),隸屬雙翅目Diptera實蠅科Tephritidae寡鬃實蠅屬Dacus Fabricus,1992年寡鬃實蠅屬被修訂為果實蠅屬Bactrocera Macquart,南亞實蠅也稱為南亞果實蠅和南亞寡鬃實蠅,俗名瓜蛆、蹦蹦蟲、黃蜂子。南亞實蠅于1849年在福建省首次報道,但在很長一段時間內沒有造成較大為害[1]。至今,南亞實蠅已在亞洲的熱帶和亞熱帶以及南太平洋地區有分布,近年在我國隨著水果蔬菜等農產品的頻繁貿易和調運,迅速擴散,目前為止已經在云南、廣東、四川和河南等多個省分布和為害[25]。南亞實蠅以雌成蟲產卵于果實內,卵孵化為幼蟲后取食果肉組織,被害后的組織在一定條件下會變畸形或直接腐爛,輕者降低果實的品質和產量,重者造成絕收,給農業生產帶來了巨大的損失[68]。南亞實蠅危害大,擴散快的主要原因之一就是它的寄主范圍廣,目前報道的寄主植物種類達16科91種,包括葫蘆科、杜英科和茄科的多種重要農作物[910]。南亞實蠅的為害給多種農作物造成較大的經濟損失,如在重慶地區5月-9月南亞實蠅對南瓜的為害率最高可達73.10%[4];對海南省瓜菜種植園中的葫蘆科、茄科、番木瓜科和西番蓮科等作物均能造成嚴重危害和減產[11]。此外,南亞實蠅的大量野生寄主種類能促進南亞實蠅的擴散和為害,導致防治困難[1213]。在南亞實蠅發生的地區,其成蟲常年可見,且一年內有兩次高峰期,分別為1月-3月和6月-7月,且夏季的種群數量大于春季[14]。
昆蟲屬于典型的變溫動物,其生長發育易受到溫度的影響[1516]。當環境溫度超出昆蟲正常生長發育的適溫范圍,即使是短暫的也會對昆蟲的生長或存活造成巨大影響甚至導致種群滅亡[16]。依據溫度變化高低及作用時間的長短,可將溫度變化對昆蟲的影響分為長時高低溫脅迫和短時高低溫脅迫,由于脅迫類型的不同,對昆蟲產生的影響也不一致[17]。在氣候變暖的趨勢下,高溫的范圍、頻率和持續時間持續增加[1820]。過去幾十年以來,全球平均氣溫呈逐步上升趨勢[21],
與持續性有規律的溫度變化相比,早春天氣不穩定,日氣溫往往與正常氣溫相差甚遠,溫度的突然短暫變化對昆蟲的生長發育和繁殖可能產生更大影響。在短時高溫處理后,亞洲玉米螟 Ostrinia furnacalis卵的孵化率、幼蟲的化蛹率、蛹的羽化率和成蟲存活率都隨處理溫度的升高而逐漸降低[22];徐天楊的研究表明,玉米螟赤眼蜂Trichogramma ostriniae經過42℃的短時高溫處理3 h和4 h后,雌蜂的壽命顯著縮短[23];羅敏等研究表明在經過2 h的2~14℃的低溫脅迫后,廣聚螢葉甲Ophraella communa除蛹以外的其他蟲態的發育及存活率受到顯著影響,且成蟲壽命及產卵量也顯著降低[24];當東亞飛蝗Locusta migratoria manilensis (Meyen) 受到≤5℃的低溫脅迫時,其各蟲態的取食行為基本停止,當處理溫度為0℃以下時,1齡幼蟲只能存活18 h[25]。南亞實蠅種群增長的最適溫區為25~31℃,在此溫度區間內其種群增加最快,若外界溫度偏離了南亞實蠅的最適溫區,則會對南亞實蠅的繁殖和發育帶來積極或消極的影響,從而影響到南亞實蠅種群的發展[26]。
在我國多數地區,南亞實蠅全年均有發生,必然會經受到由于初春氣候的不穩定及夏季溫度的驟升而出現偏離正常溫度較大的短時高低溫,這樣不正常的短時低溫和高溫可能對南亞實蠅的種群造成影響。本研究在實驗室模擬以上情景,研究了12 h的短時低溫(-4、0、4、8℃)脅迫及12 h的短時高溫(34、38、42、46℃)脅迫對南亞實蠅生長發育和生殖的影響。為探討在早春和夏季的田間控制南亞實蠅種群提供基礎,最終為南亞實蠅的田間綜合防治提供參考。
1 材料與方法
1.1 試驗材料與溫度設定
供試南亞實蠅于2017年采自云南省紅河州蒙自市南郊菜園,后置于養蟲室中,在溫度(24±1)℃、RH(70±5)%條件下繼代飼養5代。試驗中的短時高低溫處理均在人工氣候箱及冰箱中進行,RH(70±5)%,光周期L∥D=14 h∥10 h。處理的溫度設定為-4、0、4、8、24、34、38、42、46℃,其中對照組溫度為(24±1)℃,0℃以下的短時低溫試驗在冰箱進行。每個處理設4個重復,每個溫度處理12 h。處理后立即轉入養蟲室,在溫度(24±1)℃、RH(70±5)%條件下飼養。
試驗儀器:BIC300人工氣候箱,上海博迅醫療設備廠。SZX7體視顯微鏡,日本奧林巴斯株式會社。BCD201STPA冰箱,海爾集團。
西葫蘆Cucurbita pepo和南瓜C.moschata均購買自沃爾瑪超市的有機食品區,洗凈晾干后保存于4℃冰箱冷藏室中備用。
1.2 試驗方法
1.2.1 短時高低溫對卵、幼蟲和蛹生長發育的影響
選取1日齡卵為試驗材料,每組10粒卵,為1次重復,放置于事先準備好的1 cm厚西葫蘆切片上,并置于7.5 cm培養皿中。將盛有卵的培養皿放置于人工氣候箱中進行短時高溫或短時低溫處理處理12 h,0℃以下的短時低溫處理在冰箱進行。處理后的卵立即放入養蟲室進行飼養,每隔24 h利用體視顯微鏡觀察記錄卵的發育進度及存活情況,直至供試卵孵化或死亡(卵發黑變色或失水干癟視為死亡)。每個溫度為1個處理,每個處理設置4次重復(即4組卵)。
選取1日齡幼蟲作為試驗材料,每組10頭幼蟲,為1次重復,放置于切成4 cm厚成塊的西葫蘆上,后放入特制的紙杯中,紙杯中含有人工飼料和帶有幼蟲的西葫蘆,紙杯放入透明玻璃罐中,用紗網袖套封口。將盛有幼蟲紙杯的玻璃罐放入提前開啟設置好的人工氣候箱進行短時高溫或短時低溫處理處理12 h,0℃以下的短時低溫處理在冰箱進行。處理后的幼蟲立即放入養蟲室進行飼養,定時更換補充西葫蘆確保幼蟲有充足的食物,同時觀察幼蟲發育進度、存活情況及其發育歷期。幼蟲即將化蛹時,將紙杯置于底部鋪有3 cm厚細沙土的沙盆里,西葫蘆上蓋一浸濕3層的紗布補充水分,每隔24 h觀察記錄1次幼蟲的化蛹情況,直至供試幼蟲化蛹或死亡[24]。每個溫度為1個處理,每個處理設置4次重復。
選取1日齡的蛹為試驗材料,每組為10頭蛹,為1次重復,將蛹放置于鋪有浸潤濾紙的直徑7.5 cm培養皿中,將培養皿放入透明玻璃罐中,玻璃罐口用紗網袖套封口。將盛有蛹的玻璃罐放入提前開啟設置好的人工氣候箱進行短時高溫或短時低溫處理處理12 h,0℃以下的短時低溫處理在冰箱進行。處理完成后立即將蛹放入養蟲室,定時噴水補充水分,同時每隔24 h觀察記錄蛹的發育進度、存活情況及其發育歷期,直至供試蛹羽化或死亡(蛹發黑變色或干癟破損,則視為死亡)。每個溫度為1個處理,每個處理設置4次重復。
1.2.2 短時高低溫對成蟲存活、繁殖、壽命及后代性比的影響
選取24 h內羽化的同一批成蟲作為試驗材料。把1頭雌蟲與1頭雄蟲放置于1個透明玻璃罐中,并用2個7.5 cm培養皿分別盛放水和雀巢果珍粉供其補充水和食物,盛水的培養皿內放置脫脂棉,在其上鋪蓋濕潤濾紙,防止實蠅溺亡。玻璃罐用紗網袖套封口,便于添加水和食物,將盛有成蟲的玻璃罐放入提前開啟設置好的人工氣候箱進行短時高溫或短時低溫處理處理12 h,其中的0℃以下短時低溫處理在冰箱進行。處理完成后將成蟲立即放入養蟲室,每罐每日放入1塊1 cm3的南瓜塊用來收集卵,每隔24 h更換1次并觀察記錄產卵前期、日產卵數、單雌產卵量和成蟲壽命,直至供試成蟲全部死亡。將每日收集到的卵分組放入養蟲籠中,并以西葫蘆飼養,待成蟲羽化后分別統計不同處理下成蟲的后代雌性比。每10對南亞實蠅為1個重復,每個溫度下的處理設置4次重復。
1.2.3 計算公式
相關參數計算公式如下:
校正死亡率=[(處理死亡率-對照死亡率)/(1-對照死亡率)]×100%[27];
校正存活率=1-校正死亡率[27];
某蟲態的發育歷期=(Σ存活的某蟲態的發育歷期)/某蟲態存活數量[2829];
LT50(半致死溫度):某一特定的處理時間內,受試動物死亡一半時所需要的溫度。
S(x)=[exp(a-bx)]/[1+exp(a-bx)]
式中:S(x)為昆蟲種群在一定時間或低溫下的死亡率(%),x為溫度T,a和b為模型參數。當死亡率S(x)為50%時,即a-bx=0,x=a/b,x值即致死中溫度(LT50)[30]。
1.2.4 數據處理
數據分析采用SPSS 22.0軟件,使用Duncan氏法在α=0.05水平上比較不同蟲態對不同溫度耐熱差異顯著性,不同性別成蟲在同一溫度壽命差異顯著性通過t檢驗(P<0.05)進行分析[28]。
2 結果與分析
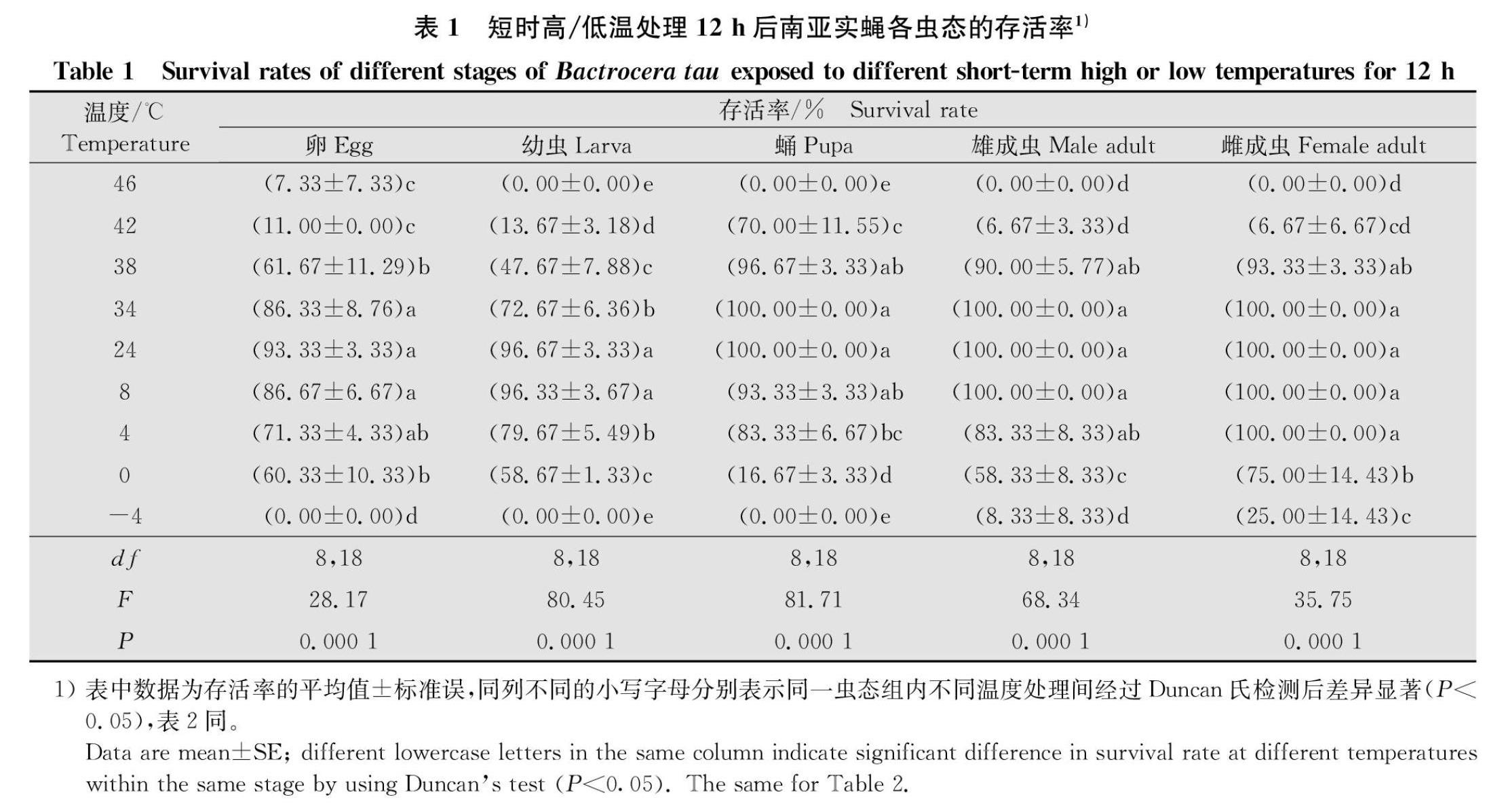
2.1 短時高/低溫度處理對南亞實蠅不同蟲態存活率的影響
不同短時高/低溫度處理對不同南亞實蠅蟲態的存活率有著顯著的影響(卵:F8,18=28.17,P=0.000 1;幼蟲:F8,18=80.45,P=0.000 1;蛹:F8,18=81.71,P=0.000 1;雌成蟲:F8,18=35.75,P=0.000 1;雄成蟲:F8,18=68.34,P=0.000 1)(表1)。短時高溫處理后,卵、幼蟲、蛹和雌雄成蟲的存活率隨著處理溫度的升高逐漸降低,當處理溫度上升至42℃時,蛹和雌雄成蟲的存活率顯著低于對照溫度下的存活率(蛹:F1,4=7.808 0,P=0.049 1;雌:F1,4=196.000 0,P=0.000 2;雄:F1,4=784.000 0,P=0.000 1),且蛹的存活率仍達到70.00%,但雌雄成蟲的存活率僅為6.67%;卵
在38℃和幼蟲在34℃時的存活率分別
顯著低于對照(卵:F1,4=7.977 0,P=0.047 6;幼蟲:F1,4=11.172 0,P=0.028 8),且存活率分別為61.67%和72.67%。在短時低溫處理后,隨著處理溫度的降低,卵、幼蟲、蛹和雌雄成蟲的存活率逐漸降低。當處理溫度下將至0℃時,卵和雌雄成蟲的存活率為60.33%,75.00%和58.33%,均顯著低于對照溫度下的存活率(卵:F1,4=9.238 0,P=0.038 4;雌:F1,4=8.308 0,P=0.044 9;雄:F1,4=25.000 0,P=0.007 5);在-4℃處理溫度下,雄雌成蟲仍有較低的存活率,存活率分別為8.33%和25.00%;4℃低溫下,幼蟲和蛹的存活率顯著低于對照溫度下的存活率(幼蟲:F1,4=8.151 0,P=0.046 2;蛹F1,4=9.308 0,P=0.038 0)(表1),分別為79.67%和83.33%。綜合不同短時高低溫脅迫下南亞實蠅各蟲態的存活率結果表明,隨著處理溫度的升高/降低,南亞實蠅各蟲態的存活率均降低,在極端高溫46℃下,幼蟲、蛹和雌雄成蟲均不能存活;在極端低溫-4℃時,僅雌雄成蟲有較低的存活率。
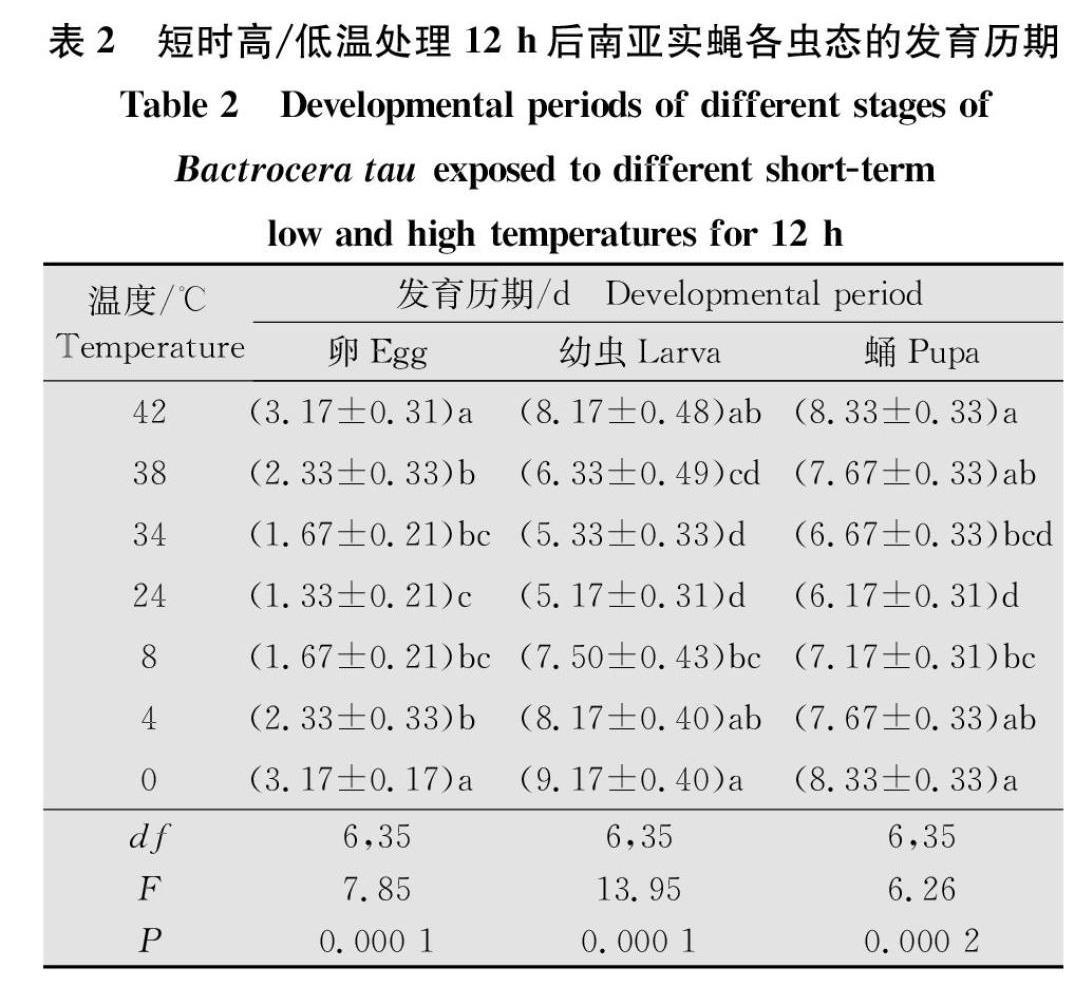
2.2 短時高/低溫對南亞實蠅不同蟲態發育歷期的影響
不同的短時高/低溫度處理南亞實蠅卵、幼蟲和蛹,結果顯示短時高/低溫對不同蟲態的發育歷期有顯著影響(卵:F6,35=7.85,P=0.000 1;幼蟲:F6,35=13.95,P=0.000 1;蛹:F6,35=6.26,P=0.000 2)(表2)。隨著處理溫度的升高/降低,卵、幼蟲和蛹的發育歷期逐漸升高,其中,在對照溫度下卵的發育歷期最短,為1.33 d,處理溫度為42℃和0℃時均為最長,達3.17 d;當處理溫度上升至38℃或下降至4℃時,卵的發育歷期都顯著高于對照組(38℃:F1,10=6.429 0,P=0.029 6;4℃:F1,10=14.412 0,P=0.003 5);幼蟲的發育歷期在對照溫度下最短,為5.17 d,42℃與0℃處理下的發育歷期最長,分別達到8.17 d和9.17 d;當處理溫度上升至42℃或下降至8℃時,該溫度下幼蟲的發育歷期顯著高于對照組(42℃:F1,10=27.931 0,P=0.000 4;8℃:F1,10=19.600 0,P=0.001 3);蛹的發育歷期在對照溫度下最短,為6.17 d,42℃與0℃處理下的發育歷期最長,達8.33 d;當處理溫度為38℃或8℃時,蛹的發育歷期顯著長于對照組(38℃:F1,10=10.946 0,P=0.007 9;8℃:F1,10=5.249 0,P=0.044 2)(表2)。綜合不同短時高低溫處理南亞實蠅各蟲態的發育歷期結果表明,在短時高溫(38~42℃)和短時低溫(0~8℃)處理時,隨著溫度的升高/降低,3種蟲態的發育歷期逐漸升高,且均在42℃和0℃時歷期為最長。
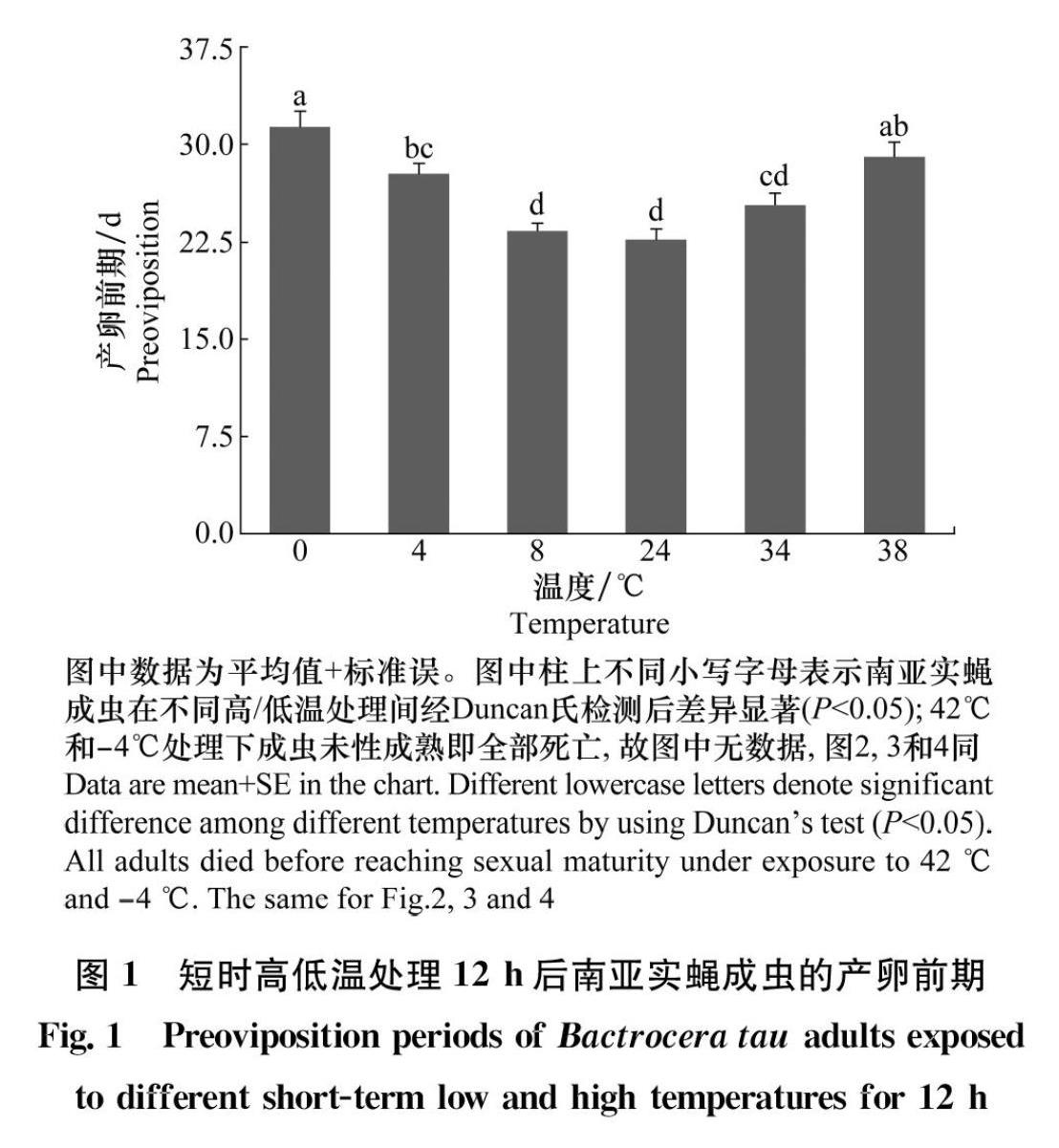
2.3 短時高/低溫對南亞實蠅成蟲產卵前期的影響
分別在不同短時高/低溫條件下進行處理,南亞實蠅成蟲的產卵前期之間存在顯著性差異(F5,12=12.320 0,P=0.000 2)(圖1)。隨著處理溫度的升高/降低,產卵前期逐漸延長,其中,當處理溫度上升至38℃和下降至4℃時的產卵前期顯著高于對照組(38℃:F1,4=19.000 0,P=0.012 1;4℃:F1,4=16.071 0,P=0.016 0)(圖1)。綜合不同短時高/低溫處理下南亞實蠅成蟲的產卵前期結果表明,在短時高溫處理后(34~38℃),成蟲的產卵前期隨著溫度的升高逐漸延長,對照溫度處理下產卵前期最短,同樣的,在低溫處理時(0~8℃)產卵前期隨著溫度的降低,逐漸延長,0℃處理下的產卵前期最長。
2.4 短時高/低溫對南亞實蠅成蟲單雌產卵量的影響
不同短時高/低溫處理對南亞實蠅成蟲的單雌產卵量有著顯著的影響(F5,12=31.126 0,P=0.000 1)(圖2)。其中,在短時高溫處理后,隨著處理溫度的升高,單雌產卵量逐漸降低,當處理溫度達到38℃時的單雌產卵量顯著低于對照組(F1,4=73.144 0,P=0.001 0),且在38℃時的單雌產卵量最少;在短時低溫處理后,隨著處理溫度的降低,單雌產卵量先增加后降低,8℃處理成蟲后單雌產卵量顯著高于對照組(F1,4=12.498 0,P=0.024 1),后隨著處理溫度的降低,單雌產卵量逐漸減少,在0℃時最少。
2.5 短時高/低溫對南亞實蠅成蟲壽命的影響
成蟲壽命也與溫度密切相關,不同短時高/低溫處理對南亞實蠅雌雄成蟲的壽命有著顯著的影響。分別在短時高/低溫條件下處理,不同溫度下同一性別成蟲的壽命之間存在顯著性差異(雌:F7,16=16.475 0,P=0.000 1;雄:F7,16=16.437 0,P=0.000 1)(圖3)。其中,在短時高溫處理后,雌雄成蟲的壽命都隨著處理溫度的升高呈現先增加后減少的趨勢,雌雄成蟲的壽命在34℃時先增加,后隨著處理溫度的逐漸升高,壽命逐漸縮短,在42℃時最短,且在42℃時雌雄壽命均顯著低于對照組的壽命(雌:F1,4=44.929 0,P=0.002 6;雄:F1,4=41.411 0,P=0.003 0);短時低溫處理之后,雌雄成蟲的壽命都隨著處理溫度的降低而縮短,當處理溫度下降至4℃時,雌雄成蟲的壽命顯著低于對照組(雌:F1,4=8.278 0,P=0.045 1;雄:F1,4=8.123 0,P=0.046 4)。同一處理溫度下雌成蟲的壽命略長于雄成蟲,但兩者差異未達到顯著水平(圖3)。
2.6 短時高/低溫對南亞實蠅后代雌性比的影響
分別在短時高/低溫條件下處理成蟲,南亞實蠅的后代雌雄性比之間存在顯著性差異(F5,12=24.513 0,P=0.000 1)(圖4)。隨著處理溫度的升高/降低,南亞實蠅后代雌性比逐漸增加,其中,當處理溫度上升至34℃/下降至4℃時的后代雌性比顯著高于對照組(34℃:F1,4=9.829 0,P=0.035 0;4℃:F1,4=8.887 0,P=0.040 7)。在高溫處理后(34~38℃),后代性比隨著溫度的升高而增加,對照溫度處理下最低,同樣的,在低溫處理時(0~8℃)后代性比隨著溫度的降低,逐漸升高。
3 結論與討論
許多研究表明,溫度對昆蟲的生長發育起著至關重要的作用。每種昆蟲都有其特定的溫度適應范圍,如果溫度超過這種昆蟲的耐受范圍,它的生命活動就會受到抑制,嚴重的會導致昆蟲死亡[31]。本研究結果表明,南亞實蠅各個蟲態的存活率與溫度密切相關,隨著短時高溫或低溫處理中溫度的升高或降低,各個蟲態的存活率逐漸降低,當處理溫度≥42℃時,除蛹以外的其余各蟲態的存活率均低于50%,當處理溫度≤-4℃時,除了雌雄成蟲外,其余各蟲態均死亡,且該極端溫度下,雌成蟲的存活率高于雄成蟲。褚世海等的研究表明在35℃高溫處理黑點切葉野螟Herpetogramma basalis幼蟲16 h后,幼蟲存活率小于25%,但隨著處理溫度的升高,幼蟲的發育歷期逐漸延長[32];瓜實蠅B.cucurbitae被短時高溫處理后,各蟲態存活率降低,發育歷期延長,在24~42℃溫度范圍內,成蟲產卵量逐隨著短時處理溫度的升高逐漸減少[28];隨著處理溫度從14℃降至2℃持續2 h,廣聚螢葉甲Ophraella communa除蛹之外所有蟲態的發育和存活都受到顯著影響,隨著處理溫度的降低,成蟲存活率顯著降低[24];櫟黃枯葉蛾Trabala vishnou gigantina越冬卵在被≤-30℃的低溫處理后,無論處理時間的長短,均能導致越冬卵的存活率大幅度下降[33]。此外,在一定的溫度范圍內,意大利蝗Calliptamus italicus通過調節抗逆物質和保護酶的含量,能夠在較高的溫度下存活,但由于抗逆物質和保護酶的含量不同,雌性具有更快的反應速率和更強的耐高溫性[34]。這與本文結果相一致。在初春和夏季,當南亞實蠅遭遇短時高低溫后,對南亞實蠅各蟲態的存活率均有著不利影響。
昆蟲的不同發育階段可以分為可活動階段和非活動階段,成蟲和幼蟲為可活動階段,卵和蛹為非活動階段,昆蟲能夠改變自己的行為和生理代謝,以避免環境溫度升高時高溫脅迫造成的損害,處于活動階段的昆蟲(成蟲和幼蟲)有可能逃離目前的有害環境,避免高溫損害,然而,在類似情況下處于非活動階段(卵和蛹)的昆蟲只能通過提高其固有的耐熱性來抵御不利影響[35]。因此,非活動階段昆蟲的耐熱性通常高于活動階段[3637]。不同的短時高/低溫度脅迫下,南亞實蠅發育歷期、產卵前期、單雌產卵量、壽命以及后代雌雄性比有著顯著的差異,隨著短時高/低溫處理溫度的升高和降低,發育歷期、產卵前期和后代性比逐漸增加;隨著高溫處理溫度的升高,雌雄成蟲的壽命先增加后減少,雌成蟲的單雌產卵量逐漸減少;隨著低溫處理溫度的降低,成蟲壽命逐漸縮短,但雌成蟲的單雌產卵量呈現先增加后減少。該結果與其他研究結果相似,例如,Ma等的研究表明,當溫度從29℃升高到34℃時,麥無網長管蚜Metopolophium dirhodum 的壽命和繁殖力均顯著降低[38]。在43℃和45℃高溫處理后,懸鈴木方翅網蝽Corythucha ciliata的存活率、壽命和繁殖力均降低[39]。以往的研究認為,環境中過高或過低的溫度都對昆蟲的生殖產生不利影響[40]。但大多數結論均是建立在持續溫度處理的研究結果上,本文則是研究短時溫度變化對生長發育及繁殖的影響,與恒溫處理的結果有差異,相對較高的短時低溫(8℃)處理成蟲12 h后,對單雌產卵量有顯著促進作用,但并不是所有短時低溫溫度處理均對單雌產卵量有促進作用,過低或過高的短時溫度對其單雌產卵量仍然會造成損傷。從能量角度分析,大多昆蟲在遭遇低溫后會將更多的能量用于生殖過程,從而降低存活率[41]。昆蟲的產卵不僅僅與溫度有關,還與濕度有著密切的關系,于瑩等的研究表明在高溫干旱條件下,褐飛虱Nilaparvata lugens雌成蟲體重、單雌產卵量和卵孵化率均下降[42];楊帥等的研究表明,咖啡豆象Araecerus fasciculatus在相對濕度為90%時其成蟲的壽命和單雌產卵量最高[43];草地螟Loxostege sticticalis在條件為21~22℃和RH60%~80%下,草地螟存活率高,生殖能力強[44]。以上研究表明相對高濕的環境對昆蟲種群的發展也有影響,因此考慮溫度條件的同時也應考慮濕度條件。本研究中,34℃的短時高溫會使得雌雄成蟲壽命略有延長,但與24℃下壽命無顯著差異,隨著處理溫度繼續的上升,則壽命縮短。其他研究也與該結果相似,郭蘇帆的研究表明,韭菜遲眼蕈蚊Bradysia odoriphaga和異遲眼蕈蚊B.difformis成蟲的壽命隨著處理溫度(24~34℃)的升高呈現出先增加后減少[45]。雖然本研究結果反映出短時高溫和低溫對南亞實蠅的繁殖和發育存在著不利的影響,但相對較低的高溫處理對雌雄成蟲的壽命有一定的延長作用,相對較高的低溫處理對繁殖力有一定的促進作用,因此在南亞實蠅發生的地區,在初春出現短時低溫后,應對南亞實蠅種群采取適當的控制措施。本研究系統地研究了短時高/低溫脅迫對南亞實蠅生長發育和繁殖的影響,但短時低溫和短時高溫脅迫后昆蟲體內的生理活性物質的變化有待于進一步研究。
參考文獻
[1] WALKER F. List of the specimens of dipterous insects in collection of the British Museum, part4 [M]. London: British Museum, 1849: 6891172.
[2] SINGH S K, KUMAR D, RAMAMURTHY V V. Biology of Bactrocera (Zeugodacus) tau (Walker) (Diptera: Tephritidae) [J]. Entomological Research, 2010, 40(5): 259263.
[3] 汪興鑒. 東亞地區雙翅目實蠅科昆蟲[J]. 動物分類學報, 1996, 21(S1): 1338.
[4] 李小珍. 南亞果實蠅種群特征及其對食料和熱脅迫的生理調控機制[D]. 重慶: 西南大學, 2007.
[5] 毛紅彥, 丁華鋒, 焦永吉, 等. 2013-2017年河南省南亞果實蠅種群動態監測[J]. 植物檢疫, 2020, 34(1): 8284.
[6] 周鎖奎, 李廣學, 邱仲華, 等. 南亞寡鬃實蠅生物學特性觀察及防治研究[J]. 植物保護, 1993, 19(5): 1112.
[7] HASYIM A, MURYATI, KOGEL W J. Population fluctuation of adult males of the fruit fly, Bactrocera tau Walker (Diptera: Tephritidae) in passion fruit orchards in relation to abiotic factors and sanitation [J]. Indonesian Journal of Agricultural Science, 2008, 9(1): 2933.
[8] 張艷. 海南省三種熱帶果園實蠅害蟲種群動態及誘捕防控技術研究[D]. 海口: 海南大學, 2013.
[9] BAIMAI V, PHINCHONGSAKULDIT J, SUMRANDEE C, et al. Cytological evidence for a complex of species within the taxon Bactrocera tau (Diptera: Tephritidae) in Thailand [J]. Biological Journal of the Linnean Society, 2000, 69(3): 399409.
[10]張艷, 陳俊諭. 南亞果實蠅國內研究進展[J]. 熱帶農業科學, 2018, 38(11): 7077.
[11]林明光, 汪興鑒, 曾玲, 等. 海南果蔬實蠅種類、地理分布及危害調查[J]. 植物檢疫, 2013, 27(5): 8589.
[12]JAMNONGLUK W, BAIMAI V, KITTAYAPONG P. Molecular phylogeny of tephritid fruit flies in the Bactrocera tau complex using the mitochondrial COI sequences [J]. Genome, 2003, 46(1): 112118.
[13]KITTHAWEE S, DUJARDIN J P. The geometric approach to explore the Bactrocera tau complex (Diptera: Tephritidae) in Thailand [J]. Zoology, 2010, 113(4): 243249.
[14]韋淑丹. 廣西南寧市瓜類實蠅的發生為害及瓜實蠅生物學和生態學特性[D]. 南寧: 廣西大學, 2011.
[15]陳瑜, 馬春森. 氣候變暖對昆蟲影響研究進展[J]. 生態學報, 2010, 30(8): 21592172.
[16]馬春森, 馬罡, 趙飛. 氣候變暖對麥蚜的影響[J]. 應用昆蟲學報, 2014, 51(6): 14351443.
[17]王艷敏, 仵均祥, 萬方浩, 等. 昆蟲對極端高低溫脅迫的響應研究[J]. 環境昆蟲學報, 2010, 32(2): 250255.
[18]MEEHL G A. More intense, more frequent, and longer lasting heat waves in the 21st century [J]. Science, 2004, 305(5686): 994997.
[19]HANSEN J, SATO M, RUEDY R. Perception of climate change [J]. Proceedings of the National Academy of Sciences of the United States of America, 2012, 109(37): 1472614727.
[20]SENEVIRATNE S I, DONAT M G, MUELLER B, et al. No pause in the increase of hot temperature extremes [J]. Nature Climate Change, 2014, 4(3): 161163.
[21]EASTERLING D R. Climate extremes: observations, modeling, and impacts [J]. Science, 2000, 289(5487): 20682074.
[22]劉超. 短時高溫對亞洲玉米螟的影響[D]. 沈陽: 沈陽農業大學, 2016.
[23]徐天楊. 高溫對玉米螟赤眼蜂生存和繁殖的影響[D]. 沈陽: 沈陽農業大學, 2017.
[24]羅敏, 郭建英, 周忠實, 等. 短時低溫脅迫對廣聚螢葉甲發育和生殖的影響[J]. 昆蟲學報, 2011, 54(1): 7682.
[25]陳申芝, 張合倫, 高強, 等. 低溫脅迫對東亞飛蝗存活及其生命特征的影響[J]. 亞熱帶農業研究, 2009, 5(4): 261263.
[26]劉麗紅. 溫度及補充營養對南亞實蠅Bactrocera tau(Walker)種群增長的影響[D]. 重慶: 西南農業大學, 2005.
[27]楊福田, 王慶云, 秦卓, 等. 4種殺蟲劑對甜菜龜葉甲幼蟲毒殺效果初探[J]. 中國植保導刊, 2020, 40(5): 8384.
[28]顧祥鵬, 黃禹禹, 張金永, 等. 短時高溫脅迫對瓜實蠅生長發育及繁殖的影響[J]. 環境昆蟲學報, 2020, 42(2): 391399.
[29]范秀娟, 陳丹, 孫志娟, 等. 短時高溫對煙蚜生長發育、繁殖和取食行為的影響[J]. 昆蟲學報, 2014, 57(10): 11881197.
[30]歐陽芳, 戈峰. 昆蟲抗凍耐寒能力的測定與分析方法[J]. 應用昆蟲學報, 2014, 51(6): 16461652.
[31]陳紅松. 廣聚螢葉甲對異常高溫的生殖與生理代謝響應[D]. 長沙: 湖南農業大學, 2012.
[32]褚世海, 叢勝波, 李儒海, 等. 短時高溫處理對黑點切葉野螟生長發育和繁殖的影響[J]. 福建農業學報, 2019, 34(11): 12881293.
[33]劉永華, 閻雄飛, 陸鵬飛, 等. 低溫脅迫對櫟黃枯葉蛾越冬卵發育和存活的影響[J]. 西北農林科技大學學報(自然科學版), 2018, 46(10): 126130.
[34]李爽, 王冬梅, 李娟, 等. 雌雄意大利蝗耐高溫差異及其生理生化響應對策[J]. 應用昆蟲學報, 2015, 52(4): 960967.
[35]HUEY R B, HERTZ P E, SINERVO B. Behavioral drive versus behavioural inertia in evolution: a null model approach [J]. The American Naturalist, 2003, 161(3): 357366.
[36]MARAIS E, CHOWN S L. Beneficial acclimation and the Bogert effect [J]. Ecology Letters, 2008, 11(10): 10271036.
[37]MITCHELL K A, SINCLAIR B J, TERBLANCHE J S. Ontogenetic variation in cold tolerance plasticity in Drosophila: is the Bogert effect bogus? [J]. Naturwissenschaften, 2013, 100(3): 281284.
[38]MA Chunsen, HAU B, POEHLING H. Effects of pattern and timing of high temperature exposure on reproduction of the rose grain aphid, Metopolophium dirhodum [J]. Entomologia Experimentalis et Applicata, 2003, 110(1): 6571.
[39]JU Ruiting, GAO Lei, ZHOU Xuhui, et al. Tolerance to high temperature extremes in an invasive lace bug, Corythucha ciliata (Hemiptera: Tingidae), in subtropical China [J/OL]. PLoS ONE, 2013, 8: e54372. DOI: 10.1371/journal.pone.0054372.
[40]劉安然. 溫度對昆蟲繁殖力的影響及其機制初探[J]. 江西農業, 2020(6): 105107.
[41]PAPAJ D R. Ovarian dynamics and host use [J]. Annual Review of Entomology, 2000, 45(1): 423448.
[42]于瑩, 徐紅星, 鄭許松, 等. 在高溫下干旱脅迫對褐飛虱生態適應性的影響[J]. 植物保護學報, 2013, 40(3): 193199.
[43]楊帥, 張濤, 高玉林, 等. 相對濕度對咖啡豆象生長發育、繁殖及種群增長的影響[J]. 應用昆蟲學報, 2016, 53(1): 121127.
[44]羅禮智, 程云霞, 唐繼洪, 等. 溫濕度是影響草地螟發生為害規律的關鍵因子[J]. 植物保護, 2016, 42(4): 18.
[45]郭蘇帆. 高溫對韭菜遲眼蕈蚊和異遲眼蕈蚊生長發育及生理代謝的影響[D]. 蘭州: 甘肅農業大學, 2018.
(責任編輯:田 喆)
