枯草芽孢桿菌S-16產抗菌蛋白發酵條件優化及蛋白分離純化
2021-03-22 03:00:12路妍楊鑫吳文慶連紫倩扈景晗周洪友
江蘇農業科學 2021年2期
路妍 楊鑫 吳文慶 連紫倩 扈景晗 周洪友
摘要:以抑菌率為指標,采用單因素試驗和響應面法對枯草芽孢桿菌S-16菌株產抗菌蛋白的發酵條件進行優化。結果表明,以可溶性淀粉為碳源,大豆蛋白胨為氮源,初始pH值為7.5的LB液體培養基,接種量為4.74%,在溫度為34 ℃條件下振蕩培養 53 h,轉速為 200 r/min為菌株最佳發酵條件。在此條件下純化得到的抗菌蛋白分子量約為35 ku,對向日葵核盤菌抑菌率達58.6%,比優化前提高了3.19倍。穩定性試驗結果表明,抗菌蛋白0~80 ℃、pH值為2~10時均有抑菌活性,說明該蛋白具有很好的熱穩定性和酸堿穩定性。
關鍵詞:枯草芽孢桿菌;抗菌蛋白;核盤菌;響應面法;純化;發酵條件優化
中圖分類號: S432.4+4 文獻標志碼: A文章編號:1002-1302(2021)02-0063-08
收稿日期:2020-05-14
基金項目:內蒙古自然科學基金(編號:2017BS0303);內蒙古高等學校科學研究項目(編號:NJZY17069)
作者簡介:路妍(1982—),女,內蒙古呼和浩特人,博士,講師,從事植物真菌病害研究。E-mail:luyan820918@126.com。
通信作者:周洪友,博士,教授,從事植物病害生物防治研究。E-mail:hongyouzhou2002@aliyun.com。
向日葵菌核病是由核盤菌[Sclerotinia sclerotiorum (Lib.) de Bary]引起的真菌病害,是世界范圍向日葵生產上危害重大并且具有毀滅性的病害之一[1-3]。向日葵核盤菌是一種兼性寄生菌,寄主范圍廣,侵染形式多樣,帶菌的種子和菌核為初侵染來源,可侵染向日葵的根、莖、葉、花盤等各個部位,造成植株局部或整體腐爛死亡,在我國發病最嚴重的是盤腐和根莖腐[4]。菌核病的有效控制包括使用以下1種或多種方法的組合:抗病品種、栽培控制和化學控制。種植抗病品種是控制土傳病害最經濟且有效的措施,但至今沒有發現對菌核病免疫和高抗的向日葵材料[1,5]。目前對于該病害的防治主要依靠化學農藥,然而殺菌劑的大量使用導致了潛在危害人類和環境的有毒化合物的積累,也導致了病原體耐藥性的增強[6]。鑒于此,研究和應用生物防治是一種很有前途的方法。根際細菌是控制土傳植物病原菌的優良藥劑[7],細菌種類如芽孢桿菌、假單胞菌、沙雷氏菌和節桿菌等已被證明能控制真菌病害[8]。
枯草芽孢桿菌(Bacillus subtilis)是一種廣泛存在于自然生態系統的非致病根際土壤細菌,具有發達的分泌系統、很強的環境適應能力和抗逆能力,具有促進植物生長和生物防治的雙重功效。能夠產生多種次生代謝產物,包括蛋白質[9]、脂肽類[10]和揮發性物質[11]等。這些物質通過抑制病原菌的生長,抑制孢子的萌發以及誘導激發植物自身的防御系統發揮抑菌作用[12]。抗菌蛋白是枯草芽孢桿菌分泌的主要活性物質,關于抗菌蛋白功能的研究也受到人們廣泛關注。劉伊強等從枯草芽孢桿菌TG26中分離的抗菌蛋白對禾谷鐮孢菌(Gibberella zeae)有較強的抑菌活性[13]。劉永峰等從枯草芽孢桿菌Bs-916中分離的抗菌蛋白對水稻紋枯病病菌(Rhizoctonia solani)和水稻惡苗病病菌(Fusarium moniliforme)具有較強的抑菌活性[14]。彭兵等從枯草芽孢桿菌菌株A發酵液中分離得到一種抗真菌活性蛋白FV,可抑制玉米小斑病病菌(Helminthosporium maydis)孢子萌發和附著胞形成,并影響菌絲細胞的形態結構[15]。研究表明,枯草芽孢桿菌及其分泌物對多種土傳病害的田間防效顯著。例如,Sha等在幾個不同地點進行的2年田間試驗中,使用枯草芽孢桿菌SYX04和SYX20可將水稻葉瘟病發病率降低73.5%~83.5%,穗瘟病發病率降低64.0%~85.6%[16]。馬新等用枯草芽孢桿菌的微囊劑防治番茄立枯病,防效明顯,可達到7276%。枯草芽孢桿菌B908(百抗)對水稻紋枯病的大田防效達70%以上[17]。陳延熙等研制的增產菌,其有效成分是枯草芽孢桿菌,在全國10多個省(市、區)50多種作物中使用增產率達10%~50%,主要用于防治水稻稻瘟病、小麥紋枯病、油菜菌核病等土傳病害[18]。然而,由于抗菌蛋白是枯草芽孢桿菌的次級代謝產物,產量低,使其在生物防治中的應用受到很大限制[17]。
提高抗菌代謝物的產量對后續抗菌機制的研究和抗菌蛋白制劑的研制至關重要。因此,對發酵條件的優化是非常有必要的。響應面法已經成功應用于多種微生物培養條件的優化。王全等采用響應面法對解淀粉芽孢桿菌(Bacillus amyloliquefaciens)的發酵條件進行優化,優化后抗菌蛋白對棉花黃萎病菌(Verticillium dahliae Kleb.)的抑菌圈面積從優化前的171.95 mm2 提高至? 320.3 mm2[19]。楊潔等采用響應面法將枯草芽孢桿菌E1R-j分泌的抗菌脂肽產量提高了2倍[20]。全鑫等利用響應面法優化全蝕病生防菌YB-81的發酵條件,優化后該菌株對全蝕病病菌的抑制率達到872%,較優化前提高了32.5%[21]。
本研究選用從向日葵根際土壤中分離得到的1株枯草芽孢桿菌菌株S-16。前期研究發現,該菌株的無菌培養濾液對向日葵核盤菌具有拮抗作用,而且大田防效明顯。本研究利用單因素法和響應面法對菌株發酵產生抗菌蛋白的條件進行優化,旨在提高抗菌蛋白的產量,并在此條件下分離純化抗菌蛋白,為后續生物農藥的研發和應用提供理論基礎。
1材料與方法
1.1試驗材料
1.1.1供試菌種枯草芽孢桿菌(Bacillus subtilis)S-16和向日葵核盤菌(Sclerotinia sclerotiorum),均由內蒙古農業大學植物病理實驗室提供。
1.1.2培養基LB基礎培養基:5 g/L酵母粉,10 g/L 蛋白胨,10 g/L NaCl,pH值為7.0。
1.2試驗方法
1.2.1種子液制備及其生長曲線繪制用滅菌牙簽挑取枯草芽孢桿菌菌株S-16單菌落,接種于LB固體培養基上,在溫度為28 ℃條件下活化培養24 h后備用。將活化培養好的單菌落接入LB液體培養基(6 mL/100 mL),置于37 ℃搖床培養,轉速為 200 r/min。分別于0、12、24、36、48、72 h取樣測定D600 nm,并繪制生長曲線以確定種子液的最佳接種時間。
1.2.2發酵上清液制備將培養好的種子液按6%(體積比)的接種量接入LB液體基礎培養基(150 mL/250 mL),置于37 ℃,200 r/min搖床培養48 h。發酵液于4 ℃,4 800 r/min離心20 min,然后用細菌過濾器(直徑為0.22 μm)過濾后,收集上清液。
1.2.3發酵液最佳硫酸銨濃度的確定及抗菌蛋白的制備發酵上清液用不同飽和度的(NH4)2SO4分級鹽析,(NH4)2SO4飽和度分別為30%、40%、50%、60%、70%、80%、90%、100%,在4 ℃冰箱中靜置24 h。于4 ℃,12 000 r/min離心 20 min后,棄上清液,將沉淀用磷酸鹽緩沖液(0.02 mol/L,pH值為6.8)溶解,經透析袋透析48 h去鹽,取出樣品,于 4 ℃,12 000 r/min離心20 min后去除沉淀,收集上清液,即為蛋白質粗提液。由于70%飽和度下蛋白質沉淀量最大,因此,后續試驗均選擇70% (NH4)2SO4來沉淀上清液。
1.2.4單因素試驗設計本試驗以王祺等報道的枯草芽孢桿菌S-16的培養條件為基礎[22],選取影響微生物發酵的7個主要因素:培養基碳源、培養基氮源、培養溫度、培養時間、初始pH值、接種量和搖床轉速進行單因素試驗,考察各因素對粗蛋白抑菌活性的影響,從而確定響應面試驗的因素及水平。每個試驗處理設置3次重復,結果采用統計軟件SPSS進行分析。不同因素的梯度條件見表1。
1.2.5響應面法設計基于單因素試驗結果,采用中心組合試驗Box-Behnken設計方案,以發酵溫度(℃)、時間(h)和接種量(%)為自變量因素,以抑菌率(Y)為響應值。自變量分別用X1、 X2、X3 來表示,變量水平用1、0、-1表示(表2)。按方程xi=(Xi-X0)/ΔX 對自變量進行編碼,其中,xi表示變量的編碼值,Xi表示變量的真實值,X0表示試驗中心點變量的真實值,ΔX表示變量的步長變化。
1.2.6穩定性試驗
1.2.6.1溫度對抗菌蛋白活性的影響取4支 10 mL 的離心管,每支離心管加入5 mL抗菌蛋白,分別置于0、10、20、30、40、60、80、100 ℃水浴中處理1 h,以未處理的樣品置于室溫作對照,冷卻后若液體有揮發,用無菌水補液至5 mL,然后采用平板對峙法,以核盤菌為指示菌分別測定其抑菌活性。
1.2.6.2pH值對抗菌蛋白活性的影響取10支離心管,每支管中加入5 mL抗菌蛋白,用HCl(1 mol/L)和NaOH(1 mol/L)將其pH值分別調為2.0、3.0、4.5、5.5、6.5、6.8、7.5、8.0、9.0、10.0,置于室溫培養2 h后,在溫度為 4? ℃條件下過夜,次日再分別將 pH 值調至 7,以未處理的蛋白粗提液為對照,然后采用“1.2.6.1”節中的平板對峙法測定其抗菌活性。
1.2.6.3紫外線對抗菌蛋白活性的影響取5個培養皿,開蓋,向每個培養皿中加入10 mL抗菌蛋白,在距離紫外燈20 cm處分別處理15、30、60、120 min,以日光燈處理為對照,然后采用“1.2.6.1”節中的平板對峙法測定其抗菌活性。
1.3數據分析
試驗數據利用SPSS 17.0統計軟件和Design-Expert( version 8.0.6)軟件進行處理分析。
1.4抗菌蛋白的分離純化
將培養好的種子液按照優化后的培養條件培養,用細菌過濾器(直徑為0.22 μm)過濾后,收集上清液。按照“1.2.3”節的方法提取蛋白質。將獲得的粗蛋白進行十二烷基硫酸鈉聚丙烯酰胺凝膠電泳(SDS-PAGE)(12%分離膠,4%濃縮膠)檢測。預先在透析袋中加入1 mL電洗脫緩沖液(50 mmol/L Tris、50 mmol/L 甘氨酸、0.1% SDS,pH值 8.9),再用刀片切下目的蛋白條帶,放入透析袋中。將收集有目的蛋白條帶膠塊的透析袋放在BioRad 電泳槽的上槽中,上、下槽均加入電洗脫緩沖液,在BioRad小型電泳槽中穩壓100 V電泳 2 h,洗脫目的蛋白。最后取出透析袋中的膠塊,經過透析和凍干,得到純化的目的蛋白,通過SDS-PAGE檢測純化結果。
1.5抗菌蛋白對核盤菌的抑制作用
采用平板對峙法,用7 mm打孔器,將活化好的核盤菌在邊緣打取菌餅,接于馬鈴薯葡萄糖瓊脂(PDA)培養基平板中央。培養至菌落直徑大小為 2 cm 時,在距離中央3 cm處用打孔器(7 mm)打取2個孔,取滅菌后的3片濾紙片蘸取抗菌蛋白,放入其中一個孔中,另取濾紙片3片蘸取磷酸緩沖液,放入等距的另外一個孔中作為空白對照,每個處理重復3次,培養3 d,每天用十字交叉法測定1次抑菌圈直徑,連續測定3 d,取平均值,計算抑菌率。
2結果與分析
2.1種子液生長曲線
由圖1可知,在12~48 h菌體數量增加速度較快,處于對數生長期,48 h后進入穩定期,菌體數量基本保持穩定。因此,選取搖床培養48 h的菌液,用無菌水調D600 nm至1.8作為種子液。
2.2單因素試驗
2.2.1不同發酵溫度對粗蛋白抑菌活性的影響由圖2可知,隨著溫度升高,抑菌率呈先升后降的趨勢,當培養溫度為30 ℃時,抑菌活性最強,抑菌率為53%。說明溫度過低或過高均不適合細菌的生長繁殖及分泌蛋白的產生。顯著性測驗結果表明,發酵溫度對抗菌蛋白的產生影響極顯著(P<0.01)。
2.2.2不同碳源對粗蛋白抑菌活性的影響由圖3可知,各碳源的抑菌率表現為可溶性淀粉>酵母提取物>葡萄糖>蔗糖>乳糖。結果表明,以可溶性淀粉為碳源時,粗蛋白的抑菌能力最強,抑菌率為52.2%,明顯高于其他碳源,因此可溶性淀粉為最佳碳源。顯著性測驗結果表明,不同碳源對粗蛋白的抑菌活性影響不顯著。
2.2.3不同氮源對粗蛋白抑菌活性的影響由圖4可知,各氮源的抑菌率表現為大豆蛋白胨>胰蛋白胨>硝酸銨>硫酸銨。以大豆蛋白胨為氮源時,粗蛋白的抑菌能力最強,抑菌率為513%,明顯高于其他氮源,因此大豆蛋白胨為最佳氮源。顯著性測驗結果顯示,不同氮源對粗蛋白的抑菌活性影響不顯著。
2.2.4不同pH值對粗蛋白抑菌活性的影響由圖5可知,粗蛋白抑菌活性隨著pH值升高呈先增加后下降的趨勢。pH值在5.0~7.5之間,蛋白產量逐漸增加, 且pH值在7.5 時抑菌活性達到最高,
抑菌率為51.7%;隨著pH值的增加,蛋白產量逐漸降低。結果表明,蛋白質分泌在過酸過堿條件下影響較大。顯著性測驗結果表明,pH值在6.0~8.0時對粗蛋白的抑菌活性影響不顯著。
2.2.5不同接種量對粗蛋白抑菌活性的影響由圖6可知,以2%為最低接種量,粗蛋白抑菌活性隨著接種量的增加呈先增加后逐漸降低的趨勢,接種量在4%時抑菌率最大,為54%。結果表明,接種量為4%時,粗蛋白的抑菌活性最大。顯著性測驗結果表明,接種量對粗蛋白的抑菌活性影響極顯著(P<0.01)。
2.2.6不同轉速對粗蛋白抑菌活性的影響由圖7可知,粗蛋白抑菌活性受到搖床轉速的影響,在轉速為200 r/min 時,抑菌率最大,為52.5%。顯著性測驗結果表明,搖床轉速對粗蛋白的抑菌活性影響不顯著。
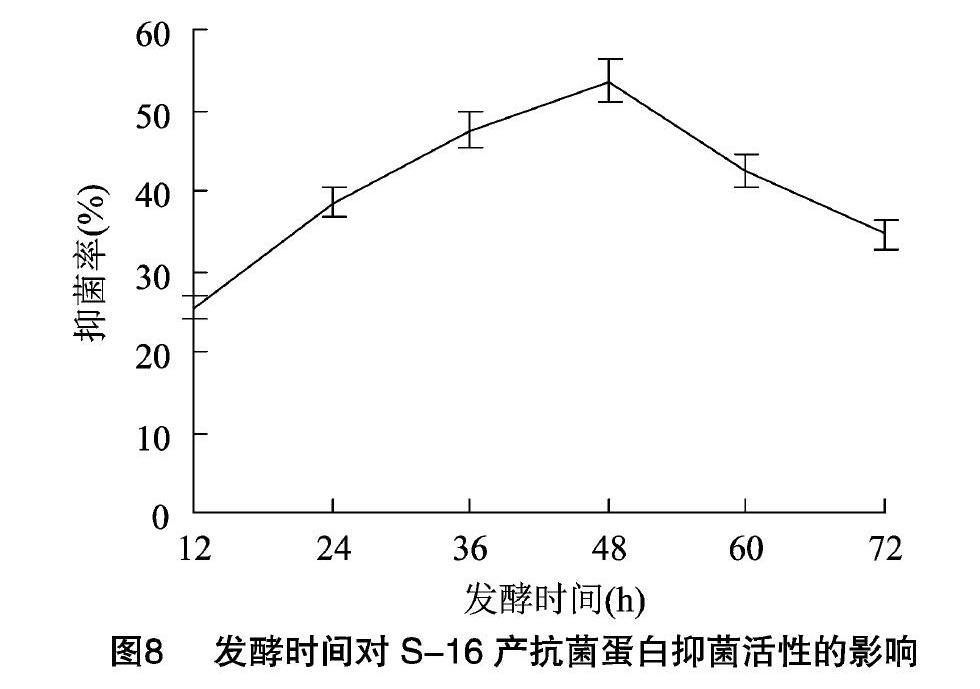
2.2.7不同發酵時間對粗蛋白抑菌活性的影響由圖8可知,在發酵初期,菌株沒有產生大量代謝物。隨著發酵時間的不斷延長,粗蛋白抑菌活性不斷增加,并在48 h時達到最大,抑菌率為53.3%,48 h 后抑菌活性開始降低。顯著性測驗結果表明,發酵時間對粗蛋白的抑菌活性影響極顯著(P<001)。
2.3響應面優化試驗
2.3.1響應面試驗設計與結果分析根據單因素試驗結果,對發酵條件影響顯著的因素有發酵溫度、接種量、發酵時間;因此,選擇上述3個因素進行響應面試驗。
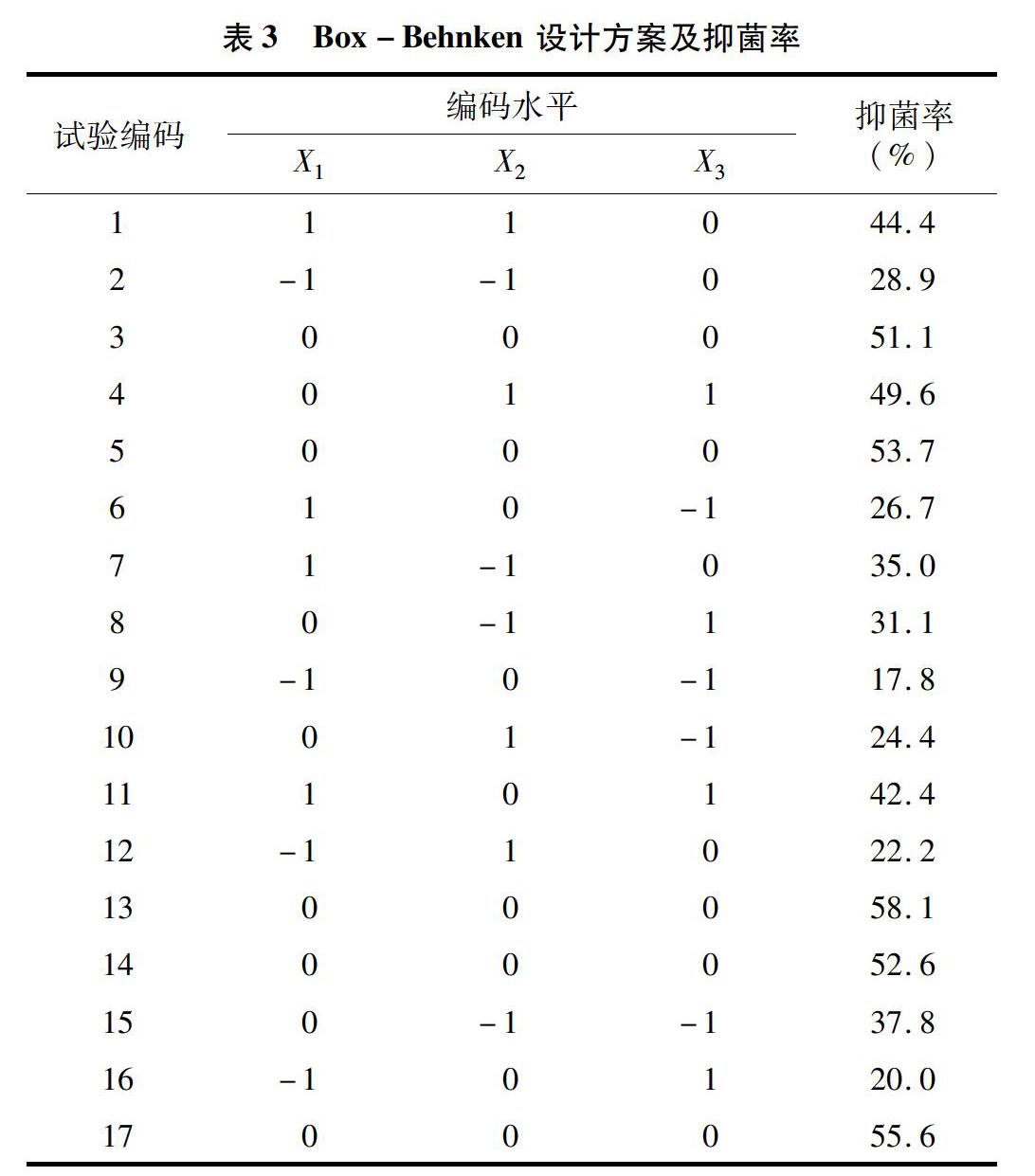
2.3.2模型的建立和方差分析在單因素試驗的基礎上,利用Design-Expert ( version 8.0.6) 軟件對表3中試驗數據進行多元回歸分析,獲得抑菌率(Y)對發酵溫度(X1)、時間(X2)和接種量(X3)的二次多項回歸模型為Y=55.6+10.575×X1+0.4×X2+6.55×X3-2225×X1X2+4.475×X1X3+13075×X2X3-15412 5×X21-1.3125×X22-13.462 5×X23。
從表4中可知,模型F值為72.66,P<0.01,達到極顯著水平,可用此模型預測枯草芽孢桿菌抗菌蛋白發酵條件對抑菌率的影響。回歸方程系數顯著性檢驗結果:X1(溫度)和X3(接種量)的一次項影響極顯著(P<0.01),X1、X2和X3的二次項影響極顯著(P<0.01),交互項X1X2(溫度和時間組合)和X2X3(接種量和時間組合)影響極顯著(P<001),X1X3(溫度和接種量組合)影響顯著(P<005)。由F值得出,試驗各因素對抑菌活性影響的主次順序表現為X1>X3>X2,即溫度>接種量>時間。
2.3.3響應面分析結果根據二次多項模型并利用Design-Expert軟件繪制出響應面分析結果(圖9、圖10、圖11),每個響應面分別表示2個獨立變量之間的相互作用,第3個變量保持在0水平,圖9是發酵溫度與時間對抑菌率的交互效應,圖10是發酵溫度和接種量對抑菌率的交互效應,圖11是發酵時間和接種量對抑菌率的交互效應。利用響應面優化器,得到優化后的發酵條件:發酵時間為53 h,發酵溫度為34.0 ℃,接種量為4.74%,預測最大抑菌率為56.5%。
2.4回歸模型的驗證試驗
為驗證預測值,用以上最優培養條件重復試驗5次,抑菌率分別為51.5%、58.6%、57.7%、562%、54.1%,平均值為55.6%,與預測值有較好的擬合性,證明利用響應曲面法優化抗菌蛋白發酵條件是可行的。在最優培養條件下,抗菌蛋白最高抑菌活性可達58.6%,比試驗設計初期的抗菌蛋白最低抑菌活性提高3.19倍。
2.5穩定性試驗
2.5.1溫度對抑菌蛋白活性的影響由圖12可知,抗菌蛋白在0~30 ℃抑菌活性與對照相比變化很小,其中20 ℃時抑菌活性最接近對照;抗菌蛋白在 40~50 ℃處理 30 min 后,其抑菌效果與對照比,變化不大;蛋白經80 ℃水浴處理 30 min后,與對照相比其抑菌活性下降21%,但仍有抑菌作用,且抑菌率接近30%;經 100 ℃ 水浴處理 30 min后抑菌活性明顯下降。結果表明,該抗菌蛋白在0~80 ℃時具有良好的熱穩定性,溫度適應范圍廣。
2.5.2pH值對抑菌蛋白活性的影響由圖13可知,處理后的抗菌蛋白與對照相比,pH值在6.5~
8.0范圍內抑菌活性有微小變化,pH值為2.0、10.0時仍分別保留原有活性的35.2%、47.1%。結果表明,該抗菌蛋白的pH值適應范圍較寬,同時也說明其具有良好的酸堿穩定性。由此說明,該蛋白在實際應用中,對土壤酸堿環境適應范圍較廣。
2.5.3紫外線對蛋白抑菌活性的影響由圖14可知,經過不同時間紫外線照射處理后,抗菌蛋白抑菌作用有明顯變化,未經紫外線照射時,抗菌蛋白對核盤菌的抑制效果最好,抑菌率達到最大,為551%。經過紫外線照射后,短時間內抑菌圈減小不明顯,抑菌活性稍有減弱,但隨著時間的增長,抑菌活性越來越低,經過120 min紫外照射后其抑菌活性基本消失。
2.6抗菌蛋白的分離純化
對分離純化的抗菌蛋白經SDS-PAGE檢測表明,在分子量約35 ku處有1條單一條帶(圖15)。
2.7抗菌蛋白對核盤菌的抑菌活性
采用平板對峙法培養3 d后,觀察發現,純化的抗菌蛋白對核盤菌有明顯的抑菌效果,采用優化后的培養條件比優化前抑菌圈的直徑明顯增加,抑菌率增加了3.19倍(圖16)。
3結論與討論
向日葵是我國東北和內蒙古中西部地區的主
要經濟作物,近年來連作現象普遍,使土壤中養分消耗過度,病蟲害越來越嚴重。化學農藥施用量不斷增加,土壤污染嚴重,地力難恢復,亟待采用合理的栽培方法,科學的防治措施,以便提高作物產量,推進向日葵產業的可持續發展。大量研究表明,枯草芽孢桿菌能夠促進植物生長發育,增加產量,能夠改善鹽堿地土壤理化性狀,使土壤微生物數量增多。此外,由于枯草芽孢桿菌無致病性,并且可以分泌多種酶和抗生素,可以起到防病促生產的作用[23-25]。
本研究所用枯草芽孢桿菌S-16菌株經過室內抑菌測驗、盆栽試驗、種子處理試驗已驗證其對向日葵菌核病有較好的抑制作用,并且發現該菌株可在向日葵根際土壤中大量定殖[22,26]。抗菌蛋白是芽孢桿菌的次生代謝產物,其產量通常受到培養基營養成分、發酵條件和高度復雜的代謝調節機制的影響。本研究旨在探究提高抗菌蛋白產量的最佳發酵條件,以純化出目的蛋白。許多研究表明,合理地使用優化方法可以顯著提高微生物代謝物的產量[27-31]。本試驗首先選擇7個對抗菌蛋白抑菌活性有影響的因素進行單因素試驗,結果篩選出3個因素對抑菌活性有顯著影響,分別是發酵溫度、發酵時間、接種量。然后,進一步針對上述3個因素做響應面試驗,最終得出該菌株產生抗菌蛋白的最佳發酵條件:可溶性淀粉為碳源,大豆蛋白胨為氮源,初始pH值為7.5,接種量為474%,培養溫度為34 ℃,培養時間為 53 h,轉速為 200 r/min,并且通過驗證,在此最佳發酵條件下,抗菌蛋白的抑菌活性比之前提高3.19倍。
據報道,枯草芽孢桿菌抗菌蛋白具有抗真菌和細菌等特性,其抗菌譜廣、穩定性高、對熱不敏感[32]。在本研究中,S-16菌株所產抗菌蛋白經熱穩定性測驗發現,在0~80 ℃熱穩定性良好,在 100 ℃ 條件下處理 30 min,抑菌活性明顯下降,且該抗菌蛋白具有較寬的pH值適應范圍,說明其酸堿穩定性好。另外,該蛋白經紫外線照射30 min以內,對抑菌活性影響小,處理1、2 h,抑菌活性明顯下降。根據上述試驗數據,說明該蛋白對溫度、pH值和短時間的紫外線不敏感。這些結果將為抗菌蛋白后續的抗菌機制研究和生防制劑的研制提供理論依據,也將為其應用于向日葵菌核病的生物防治提供參考。
參考文獻:
[1]Li Z Q,Wang Y C,Chen Y,et al. Genetic diversity and differentiation of Sclerotinia sclerotiorum populations in sunflower[J]. Phytoparasitica,2009,37(1):77-85.
[2]Gulya T,Harveson R,Mathew F,et al. Comprehensive disease survey of U.S. sunflower:disease trends,research priorities and unanticipated impacts[J]. Plant Disease,2019,103(4):601-618.
[3]Yue B,Radi S A,Vick B,et al. Identifying quantitative trait loci for resistance to Sclerotinia head rot in two USDA sunflower germplasms[J]. Phytopathology,2008,98(8):926-931.
[4]董金皋. 農業植物病理學[M]. 2版.北京:中國農業出版社,2007:190-191.
[5]孟慶林,馬立功,劉佳,等. 向日葵菌核病田間接種方法及品種抗病性研究[J]. 中國油料作物學報,2014,36(1):113-116.
[6]Talukder Z,Long Y,Seiler G J,et al. Introgression and monitoring of wild Helianthus praecox Alien segments associated with Sclerotinia basal stalk rot resistance in sunflower using genotyping-by-sequencing[J]. PLoS One,2019,14(3):e0213065.
[7]Ashwini N,Srividya S. Potentiality of Bacillus subtilis as biocontrol agent for management of anthracnose disease of chilli caused by Colletotrichum gloeosporioides OGC1[J]. Biotech,2014,4(2):127-136.
[8]Joseph B,Patra R R,Lawrence R. Characterization of plant growth promoting rhizobacteria associated with chickpea(Cicer arietinum L.)[J]. International Journal of Plant Production,2007,1(2):141-151.
[9]Zhao X Y,Zhao X M,Wei Y M,et al. Isolation and identification of a novel antifungal protein from a rhizobacterium Bacillus subtilis strain F3[J]. Journal of Phytopathology,2013,161(1):43-48.
[10]徐楊,王楠,李偉,等. 海洋枯草芽孢桿菌3512A抗真菌脂肽的分離純化及結構特性鑒定[J]. 中國生物防治,2009,25(4):328-333.
[11]Fiddaman P J,Rossall S. The production of antifungal volatiles by Bacillus subtilis[J]. The Journal of Applied Bacteriology,1993,74(2):119-126.
[12]劉雪,穆常青,蔣細良,等. 枯草芽孢桿菌代謝物質的研究進展及其在植病生防中的應用[J]. 中國生物防治,2006,22(增刊1):179-184.
[13]劉伊強,王雅平. 拮抗菌TG26的鑒定及其抗菌蛋白BI的純化和部分特性[J]. 植物學報,1994,36(3):197-203.
[14]劉永鋒,陳志誼,周明國,等. 枯草芽抱桿菌Bs-916的抑菌活性及其抑菌物質初探[J]. 農藥學學報,2007,9(1):92-95.
[15]彭兵,張樹斌,賈宇,等. 枯草芽孢桿菌菌株A抗菌蛋白的分離純化及抗真菌機理[J]. 中國農業科學,2011,44(1):67-74.
[16]Sha Y,Wang Q,Li Y. Suppression of Magnaporthe oryzae and interaction between Bacillus subtilis and rice plants in the control of rice blast[J]. Springer Plus,2016,5(1):1238.
[17]馬新,黃永,程娟,等. 枯草芽孢桿菌微囊劑的制備及其對番茄立枯病的防治效果[J]. 農藥學學報,2015,17(4):462-468.
[18]陳延熙,陳壁,唐文華. 增產菌(PGPR)增產效果的田間試驗[C]//全國微生物生態學術討論會. 1985.
[19]王全,王占利,高同國,等. 響應面法對解淀粉芽孢桿菌(Bacillus amyloliquefaciens)12-7產抗菌蛋白條件的優化[J]. 棉花學報,2016,28(3):283-290.
[20]楊潔,姬婧媛,康振生,等. 枯草芽孢桿菌E1R-j產抗菌脂肽發酵條件的優化[J]. 西北農業學報,2012,21(7):54-60.
[21]全鑫,楊艷艷,楊麗榮,等. 利用響應面法優化全蝕病生防菌YB-81的發酵條件[J]. 天津農業科學,2014,20(9):57-61.
[22]王祺,張一名,趙君,等. 枯草芽孢桿菌S-16抑菌活性物質的理化性質及培養條件的研究[J]. 中國生物防治學報,2015,31(3):439-444.
[23]Franco-Sierra N D,Posada L F,Santa-María G,et al. Bacillus subtilis EA-CB0575 genome reveals clues for plant growth promotion and potential for sustainable agriculture[J]. Functional & Integrative Genomics,2020,20(4):575-589.
[24]Hashem A,Tabassum B,Fathi Abd Allah E. Bacillus subtilis:a plant-growth promoting rhizobacterium that also impacts biotic stress[J]. Saudi Journal of Biological Sciences,2019,26(6):1291-1297.
[25]Sohaib M,Zahir Z A,Khan M Y,et al. Comparative evaluation of different carrier-based multi-strain bacterial formulations to mitigate the salt stress in wheat[J]. Saudi Journal of Biological Sciences,2020,27(3):777-787.
[26]張一名,甄熙,石林君,等. 生防菌S-16的定殖動態以及對向日葵菌核病的生防效果研究[J]. 河北農業科學,2012,16(1):36-38,75.
[27]Shu G W,Shi X Y,Chen H,et al. Optimization of goat milk with ACE inhibitory peptides fermented by Lactobacillus bulgaricus LB6 using response surface methodology[J]. Molecules,2017,22(11):2001.
[28]Wu W J,Ahn B Y. Statistical optimization of medium components by response surface methodology to enhance menaquinone-7 (vitamin K2) production by Bacillus subtilis[J]. Journal of Microbiology and Biotechnology,2018,28(6):902-908.
[29]Wang H B,Huang J G,Sun L,et al. An efficient process for co-production of γ-aminobutyric acid and probiotic Bacillus subtilis cells[J]. Food Science and Biotechnology,2019,28(1):155-163.
[30]Singh P,Shera S,Banik J,et al. Optimization of cultural conditions using response surface methodology versus artificial neural network and modeling of L-glutaminase production by Bacillus cereus MTCC1305[J]. Bioresource Technology,2013,137:261-269.
[31]劉珍珠,樊振,劉歡歡,等. 枯草芽孢桿菌的篩選及其與光合細菌復配對養殖水體的凈化[J]. 江蘇農業科學,2020,48(6):164-167.
[32]于杰,張榮意,譚志瓊,等. 枯草芽孢桿菌B25抗真菌作用及抗菌蛋白的分離純化[J]. 基因組學與應用生物學,2016,35(3):629-634.郭亞軍,趙明,陳小軍,等. 三氟苯嘧啶在稻田中的降解動態和殘留分析[J]. 江蘇農業科學,2021,49(2):71-75,80.
