巖溶區(qū)固氮植物適應(yīng)土壤氮素變化的潛在策略
2021-03-24 06:58:26潘復(fù)靜章潤(rùn)陽(yáng)秦國(guó)鑫陳英倩梁月明劉坤平
桂林理工大學(xué)學(xué)報(bào) 2021年4期
關(guān)鍵詞:植物
潘復(fù)靜, 章潤(rùn)陽(yáng), 秦國(guó)鑫, 陳英倩, 梁月明, 劉坤平, 張 偉
(1.桂林理工大學(xué) a.環(huán)境科學(xué)與工程學(xué)院; b.廣西環(huán)境污染控制理論與技術(shù)重點(diǎn)實(shí)驗(yàn)室科教結(jié)合科技創(chuàng)新基地,廣西 桂林 541006; 2.中國(guó)地質(zhì)科學(xué)院巖溶地質(zhì)研究所, 自然資源部、 廣西壯族自治區(qū)巖溶動(dòng)力學(xué)重點(diǎn)實(shí)驗(yàn)室,廣西 桂林 541004; 3.中國(guó)科學(xué)院環(huán)江喀斯特生態(tài)系統(tǒng)觀測(cè)研究站, 廣西 環(huán)江 547100)
0 引 言
我國(guó)西南巖溶地區(qū)面積達(dá)540 000 km2[1]。該地區(qū)的土壤主要由碳酸鹽巖風(fēng)化而成, 成土速率慢, 形成1 cm厚度土壤所需的時(shí)間高達(dá)幾千年至幾萬(wàn)年[2-3], 而且土-石二元結(jié)構(gòu)的光滑接觸面使得水土容易流失[4], 導(dǎo)致該地區(qū)土壤總量偏少。此外, 由于土壤是碳酸鹽巖風(fēng)化而成, 其鈣含量較高, 而鈣易與有機(jī)質(zhì)和磷結(jié)合形成穩(wěn)定化合物, 氮磷等養(yǎng)分釋放的難度大, 氮磷有效性變低[5]。土壤N和P元素是影響植物生長(zhǎng)的重要營(yíng)養(yǎng)元素, 其主要來(lái)源是土壤有機(jī)質(zhì)和母巖[6-7], 植物獲取這兩種養(yǎng)分也主要是從土壤中來(lái)。盡管巖溶地區(qū)土壤氮磷含量較同緯度地帶性土壤高, 然而土壤淺薄、 高鈣鎂和pH等制約其養(yǎng)分供給能力, 導(dǎo)致植被恢復(fù)早期的草叢階段受到氮限制、 灌木階段受到氮磷共同限制和后期的喬木階段受到磷限制的狀況[8-9]。
當(dāng)植物受到N限制或當(dāng)土壤中的可利用氮含量較低時(shí), 一些植物可以利用空氣中的氮進(jìn)行生物固氮, 這一類(lèi)植物即固氮植物。固氮植物能夠與固氮微生物形成根瘤菌, 將空氣中的氮?dú)膺€原為氨[10]。植物固氮過(guò)程受土壤環(huán)境中氮素濃度的影響, 不同的固氮植物采用不同的氮素獲取策略[11-13]。因此, 基于土壤環(huán)境可利用氮含量的響應(yīng)靈敏性程度, 固氮植物適應(yīng)土壤氮素變化的固氮策略分為兼性固氮策略、 專(zhuān)性固氮策略、 調(diào)節(jié)過(guò)度固氮策略(介于兼性和專(zhuān)性的固氮策略)共3種。兼性固氮策略是固氮植物的固氮速率隨著土壤環(huán)境中可利用氮含量的提高而降低, 固氮量以滿(mǎn)足自身需求為主; 專(zhuān)性固氮策略是固氮植物的固氮速率不隨土壤環(huán)境中的可利用氮含量的變化而變化, 即使在氮含量高的土壤中仍然具有較高的固氮速率; 調(diào)節(jié)過(guò)度固氮策略是固氮植物的固氮量少于自身需求量, 其固氮速率不會(huì)很高[12]。在土壤中添加無(wú)機(jī)氮的情況下, 大豆在不同生育期的根瘤數(shù)量下降13%~20%, 固氮酶活性下降可達(dá)19%~32%[14]。這種情況可能是改變了固氮菌的群落結(jié)構(gòu), 降低了其多樣性, 導(dǎo)致固氮速率下降[15]; 另外, 生物固氮是一種耗能很高的生物化學(xué)過(guò)程, 植物和固氮微生物在較高氮素濃度情況下直接吸收土壤中的有效氮, 以降低能耗[16]。在自然生態(tài)系統(tǒng)中, 兼性固氮策略可能比較常見(jiàn)。另外, 土壤環(huán)境的氮素濃度也會(huì)影響固氮植物對(duì)土壤磷素養(yǎng)分的獲取策略。根據(jù)前期研究中布設(shè)的21個(gè)灌叢樣方(10 m×10 m), 發(fā)現(xiàn)灌叢廣泛分布有固氮植物[17]; 巖溶區(qū)灌叢生態(tài)系統(tǒng)中固氮植物根際土的有效磷含量顯著高于非固氮植物[18]。而叢枝菌根真菌與磷酸酶是植物獲取土壤磷素的兩種重要策略[19-20]。可見(jiàn), 固氮植物的引種不僅可以消減土壤氮素的限制, 還可以活化土壤磷素[21], 因而被廣泛應(yīng)用于巖溶等生態(tài)系統(tǒng)的植被恢復(fù)。
在巖溶生態(tài)系統(tǒng), 植物從植被恢復(fù)早期的N限制逐步過(guò)渡到恢復(fù)后期的P限制[9], 也伴隨著土壤N含量的逐漸升高[22]。土壤氮含量水平的升高對(duì)固氮植物的固氮速率的影響是判斷固氮植物的固氮策略的有效指標(biāo), 可通過(guò)人為添加N素到土壤中, 研究固氮植物的固氮速率。另外, 根據(jù)文獻(xiàn)[18], 灌叢階段中固氮植物根際土的有效磷也存在顯著差異, 這可能與巖溶區(qū)不同固氮植物采用的氮磷養(yǎng)分獲取策略有關(guān)。而當(dāng)土壤N含量逐漸升高, 不同固氮植物的氮磷養(yǎng)分獲取策略可能發(fā)生變化。因此, 本文根據(jù)已有研究數(shù)據(jù), 確定香合歡(Albiziaodoratissima)與木豆(Cajanuscajan)兩種固氮植物作為研究對(duì)象, 通過(guò)添加氮肥的盆栽試驗(yàn), 探討不同固氮植物的固氮策略以及土壤氮素濃度變化對(duì)不同固氮植物的氮磷養(yǎng)分獲取策略的影響, 為巖溶區(qū)植被恢復(fù)選擇合適的固氮植物提供科學(xué)依據(jù)。
1 材料與方法
1.1 盆栽試驗(yàn)設(shè)置
盆栽試驗(yàn)設(shè)在中國(guó)科學(xué)院環(huán)江喀斯特生態(tài)系統(tǒng)觀測(cè)研究站(24°44′N(xiāo), 107°51′E)。該地區(qū)屬于亞熱帶季風(fēng)氣候, 年平均降雨量為1 389 mm, 年平均溫度為18.5 ℃。雨季主要集中在每年的4—8月, 占年降雨量的70%[23]。
2017年10—12月, 在研究站區(qū)域內(nèi)采集香合歡與木豆種子。 兩種固氮植物是根據(jù)早期在巖溶區(qū)布設(shè)的21個(gè)灌叢樣地的植被調(diào)查數(shù)據(jù)確定的, 二者是耐貧瘠、 耐干旱、 能夠形成根瘤并分布廣的固氮植物, 而區(qū)別則是香合歡為喬木樹(shù)種、 木豆為直立灌木樹(shù)種[17]。2018年2月, 從觀測(cè)站附近選擇巖溶典型灌叢階段樣地, 土壤類(lèi)型為棕色石灰土。采集0~20 cm厚度的土樣(約400 kg), 過(guò)2 mm篩后4 ℃保存。從過(guò)篩后的土壤隨機(jī)選取了5份200 g的樣品進(jìn)行養(yǎng)分含量測(cè)定, 其全氮和有效磷含量分別為4.48和1.92 mg/kg。
2018年3月, 將過(guò)篩土壤用布袋裝好, 放入滅菌鍋用蒸汽滅菌法在135 ℃下滅菌10 min, 取出后自然降溫, 立即把滅菌土填滿(mǎn)育苗袋(容積約 0.3 L), 育苗袋預(yù)先用75%酒精進(jìn)行了消毒。盆栽試驗(yàn)設(shè)計(jì)了2個(gè)氮肥添加處理: 添加氮肥(10 gN·(m2·a)-1, 化肥種類(lèi)為氯化銨(NH4Cl))與不添加氮肥(0 gN·(m2·a)-1)。每個(gè)氮肥處理設(shè)置16盆, 共32盆, 分別把兩種固氮植物的種子播到兩種處理的育苗袋各8個(gè)重復(fù)(2個(gè)處理×2種植物×8重復(fù))。種子出苗(大約2周)后, 每盆選擇2株長(zhǎng)勢(shì)相似的幼苗移栽到與上述育苗袋的處理相同的塑料盆中(容積約 13 L, 每盆裝土 10 kg, 共32盆), 待幼苗生長(zhǎng)穩(wěn)定(約1周)后, 每盆僅保留1株長(zhǎng)勢(shì)較好的幼苗, 每隔2~3天澆一次水以使土壤持水率維持在40%左右。
1.2 樣品采集
連續(xù)培養(yǎng)15周后(2018年6月), 每個(gè)處理中選擇長(zhǎng)勢(shì)相似的5盆植物(共20盆, 2個(gè)處理×2種植物×5重復(fù)), 分別采集固氮植物根瘤、 根、 莖、 葉及根際土樣品。根際土采集步驟為: 首先把土壤連同植株從培養(yǎng)盆取出, 捏松土壤, 再輕輕抖動(dòng)植株和土壤使松散的土掉落, 最后將仍粘附根系表面的土壤輕輕取下則為根際土樣品[24]。將收獲到的一部分根, 全部的莖、 葉樣品在65 ℃下烘干48 h至恒重, 稱(chēng)量, 用于測(cè)算生物量和根冠比。烘干后的根、 莖、 葉片粉碎過(guò)100目篩(0.15 mm), 用于測(cè)定植物各器官的碳、 氮和磷含量。未烘干根樣品的一半浸泡于95%乙醇保存, 用于叢枝菌根侵染率的測(cè)定; 另一半根樣品在-20 ℃保存, 用于酸性磷酸酶活性的測(cè)定。一部分根際土壤樣品在風(fēng)干后用于土壤養(yǎng)分含量的測(cè)定。
1.3 樣品測(cè)定
1.3.1 酸性磷酸酶活性測(cè)定 植物根系酸性磷酸酶活性采用MUB熒光光度法[25]測(cè)定。稱(chēng)取0.5 g植物細(xì)根(每個(gè)樣品3份),剪碎后放入5 mL離心管中。第一份加入2 mL的50 mmol·L-1醋酸鈉緩沖液(用冰醋酸調(diào)節(jié)pH 5.0);第二份加入 800 μL 醋酸鈉緩沖液和 200 μL 10 μmol·L-1的標(biāo)準(zhǔn)物(4-甲基傘形酮,4-Methylumbelliferone)溶液;第三份加入 800 μL醋酸鈉緩沖液和 200 μL 200 μmol·L-1的底物(4-甲基傘形酮磷酸酯,4-Methylumbelliferyl phosphate)溶液。37 ℃孵育1 h,加入10 μL NaOH(1 mol·L-1)終止反應(yīng),用酶標(biāo)儀(Tecan Infinite M200 PRO,Tecan, 瑞士)于 365 nm波長(zhǎng)處激發(fā)、405 nm處檢測(cè)。
1.3.2 菌根侵染率測(cè)定 叢枝菌根真菌侵染率采用根段頻率常規(guī)法[26]測(cè)定。將待測(cè)根系清洗干凈, 并用已消毒的手術(shù)剪把根系剪成1 cm長(zhǎng)的根段, 再將根段放入50 mL離心管中, 加入足夠的10 % KOH溶液浸沒(méi)根段, 放入90 ℃水浴鍋中煮至透明, 用清水洗凈根段, 再加2%鹽酸酸化, 倒掉鹽酸后加入0.01%酸性品紅染色, 染色后挑選50個(gè)細(xì)小根段壓片, 顯微鏡下觀察根段侵染狀況。
根段侵染率(%)=∑(0×根段數(shù)+10%×根段數(shù)+20%×根段數(shù)+…+100%×根段數(shù))/觀察總根段數(shù), 其中: 0, 10%, 20%, …,100%表示根段被侵染的程度。
1.3.3 固氮速率測(cè)定 根瘤的固氮速率采用乙炔還原法[20]測(cè)定。具體步驟為: 從每個(gè)處理中(每株植物)中選取10~15顆顆粒完整、 帶有部分須根的粉色、 淺黃色或者褐色的根瘤(為確保根瘤活性, 用于培養(yǎng)的根瘤不提前采摘), 放入125 mL透明的玻璃培養(yǎng)瓶中, 塞緊瓶蓋, 用高純乙炔氣體置換瓶中10 %的頂空氣體, 在相對(duì)一致的環(huán)境條件下培養(yǎng)0.5 h后, 抽取瓶中30 mL氣體裝入已抽真空的12 mL規(guī)格儲(chǔ)氣瓶中, 用GC-FID儀(Agilent GC 7890A, Agilent, USA)測(cè)定乙烯含量; 每個(gè)樣品的根瘤固氮速率做2個(gè)技術(shù)上的重復(fù)、 1個(gè)空白對(duì)照。測(cè)定乙烯后, 通過(guò)轉(zhuǎn)化可得到單位質(zhì)量根瘤的固氮速率(μmol·L-1)。
1.3.4 土壤與植物養(yǎng)分的測(cè)定 植物碳含量采用重鉻酸鉀容量法-外加熱法測(cè)定。土壤與植物氮含量采用凱氏定氮法-流動(dòng)注射儀(FIAstar 5000, FOSS, Hiller?d, Denmark)測(cè)定。土壤氨態(tài)氮和硝態(tài)氮利用2 mol·L-1KCl浸提, 用流動(dòng)注射儀(AA3, Seal, 德國(guó))測(cè)定。土壤全磷(TP)采用氫氧化鈉(片狀)熔融, 土壤有效磷(AP)用0.5 mol·L-1NaHCO3浸提, 植物磷含量則采用H2SO4+H2O2消煮, 三者都用鉬藍(lán)比色法測(cè)定[27]。
1.4 統(tǒng)計(jì)與分析
在Microsoft Excel里整理數(shù)據(jù), 用SPSS 20.0對(duì)數(shù)據(jù)進(jìn)行統(tǒng)計(jì)分析。利用獨(dú)立樣本t檢驗(yàn)及多重比較分析(LSD)方法, 比較施肥與不施肥對(duì)兩種固氮植物固氮速率、 根系磷酸酶活性、 叢枝菌根侵染率、 植物各器官養(yǎng)分、 土壤養(yǎng)分的差異。采用Pearson相關(guān)關(guān)系方法分析固氮植物固氮速率、 磷酸酶活性、 菌根侵染率與土壤養(yǎng)分含量之間的相關(guān)性。
2 結(jié) 果
2.1 氮肥添加對(duì)巖溶區(qū)固氮植物根際土壤氮磷有效性的影響
氮肥添加下, 香合歡與木豆根際土的全磷、 有效磷、 銨態(tài)氮含量顯著提高(圖1a、 b、 c); 木豆的硝態(tài)氮含量提高, 但香合歡的降低(圖1d), 香合歡的根際土有效磷含量顯著低于木豆(圖1b); 不添加氮肥條件下, 香合歡的根際土硝態(tài)氮含量顯著高于木豆(圖1d)。 總體上, 添加氮肥情況下根際土的全磷、 有效磷、 銨態(tài)氮含量顯著高于不添加氮肥的情況, 而硝態(tài)氮的差異不明顯。
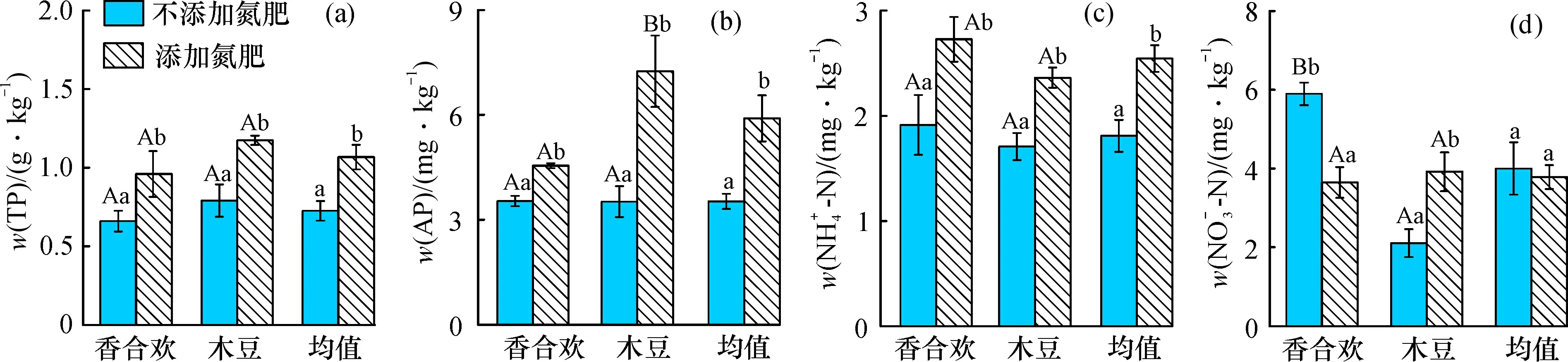
圖1 氮肥添加對(duì)兩種固氮植物根際土的全磷、 有效磷、 銨態(tài)氮和硝態(tài)氮含量的影響
2.2 氮肥添加對(duì)巖溶區(qū)固氮植物生物量、 根冠比和養(yǎng)分含量的影響
添加氮肥情況下, 香合歡的總生物量顯著提高, 木豆的提高不顯著, 總體上生物量有一定的提高(圖2a); 香合歡的根冠比顯著降低, 木豆的降低不明顯, 但總體上顯著降低(圖2b)。添加和不添加氮肥兩種情況下, 木豆的總生物量顯著高于香合歡, 但根冠比顯著低于香合歡(圖2)。
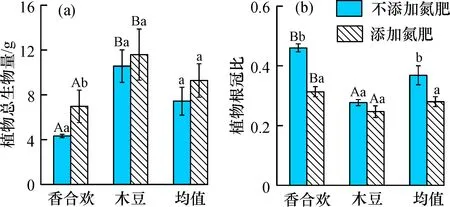
圖2 氮肥添加對(duì)兩種固氮植物總生物量與根冠比的影響
氮肥添加下, 木豆的根系氮和磷含量顯著提高(圖3b、 c); 木豆和香合歡的莖的氮含量顯著降低(圖3e)。總體上, 植物莖和葉的碳含量顯著高于不添加的情況, 而莖和葉的氮含量顯著低于不添加的情況(圖3d、 e、 g、 h)。另外, 氮肥添加下香合歡的根、 莖和葉的碳、 氮和磷的含量與木豆的不存在顯著差異; 而不添加氮肥情況下, 香合歡根、 莖和葉的氮含量和根系的磷含量顯著高于木豆(圖3b、 c、 e、h)。

圖3 氮肥添加對(duì)兩種固氮植物根、 莖和葉的碳、 氮和磷含量的影響
2.3 氮肥添加對(duì)巖溶區(qū)固氮植物固氮速率、 菌根侵染率和根系磷酸酶活性的影響
添加氮肥條件下, 香合歡和木豆的固氮速率顯著降低(圖4a); 香合歡的菌根侵染率顯著降低, 但是木豆的菌根侵染率變化不明顯, 總體上顯著降低(圖4b); 木豆的根系磷酸酶活性顯著升高, 但香合歡的根系磷酸酶活性變化不明顯, 總體上顯著升高(圖4c)。
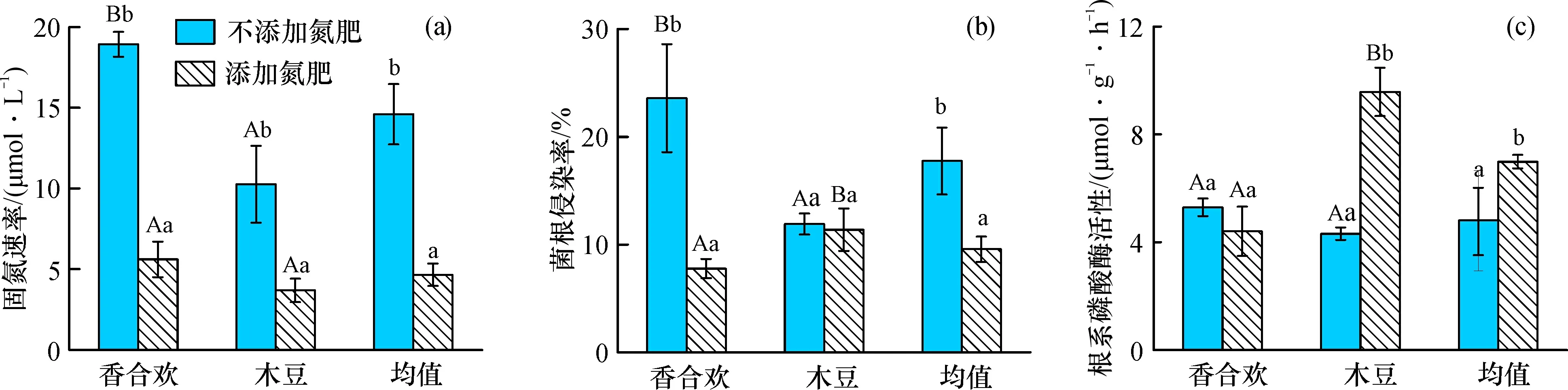
圖4 氮肥添加對(duì)兩種固氮植物固氮速率、 菌根侵染率與根系磷酸酶活性的影響
添加氮肥條件下, 香合歡的菌根侵染率和根系磷酸酶活性低于木豆, 但兩種植物的固氮速率無(wú)顯著差異。不添加氮肥條件下, 香合歡的固氮速率、 菌根侵染率、 根系磷酸酶活性均高于木豆。固氮植物的固氮速率與菌根侵染率和硝態(tài)氮存在顯著正相關(guān)關(guān)系, 但與有效磷存在顯著負(fù)相關(guān)關(guān)系; 菌根侵染率與硝態(tài)氮存在顯著正相關(guān)關(guān)系; 根系磷酸酶活性與有效磷存在顯著正相關(guān)關(guān)系(表1)。
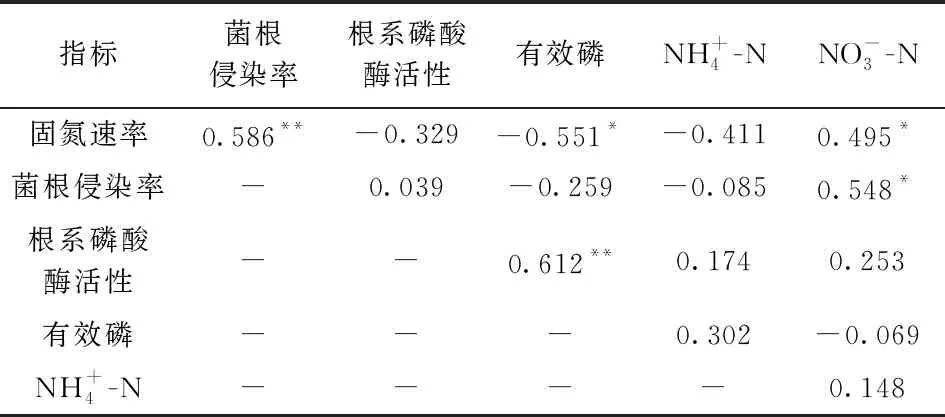
表1 植物的固氮速率、 菌根侵染率、 磷酸酶活性與土壤氮磷有效性的相關(guān)性
3 討 論
3.1 氮添加對(duì)巖溶區(qū)固氮植物生長(zhǎng)的影響
氮肥添加情況下, 總體上巖溶區(qū)兩種固氮植物總生物量顯著提高, 而根冠比顯著降低。 具體表現(xiàn)為香合歡的總生物量顯著提高, 但是根冠比顯著降低, 木豆的總生物量和根冠比變化不明顯(圖2)。可以看出, 添加氮肥影響了巖溶區(qū)固氮植物的生物量和根冠比的變化, 而對(duì)香合歡這兩方面的影響更大。氮素是植物生長(zhǎng)必須的生源要素, 而在巖溶地區(qū), 土壤低氮狀況是該地區(qū)植被恢復(fù)早期植物生長(zhǎng)受到限制的一大因素。在較低土壤含氮量的情況下, 植物為了提高吸收養(yǎng)分的效率和數(shù)量, 往往通過(guò)把生物量分配到地下, 更多的根系生物量使得根系的數(shù)量、 根尖的數(shù)量、 根系的長(zhǎng)度等得到提高[28]。當(dāng)?shù)侍砑拥酵寥喇?dāng)中, 會(huì)使得土壤中的供氮水平提高。添加氮肥的土壤有效磷與銨態(tài)氮含量顯著高于不施氮肥的情況(圖1)。在較高土壤含氮量情況下, 充足的供氮量使得植物吸收更加容易或高效, 有利于植物提高光合作用效率而加快生長(zhǎng), 生物量增長(zhǎng)迅速(圖2a)。氮肥添加情況下植物莖和葉的碳含量顯著提高(圖3), 且總生物量顯著提高(圖2a)。植物的養(yǎng)分狀況與土壤氮磷含量密切相關(guān)[29], 氮肥的添加能夠增加植物的磷含量[30]。本研究中, 氮肥添加情況下植物根系和葉片的磷含量得到一定程度的提高(圖3)。在營(yíng)養(yǎng)較為充足的情況下, 植物則會(huì)分配更多的生物量在地上部分, 而降低對(duì)地下部分的分配。因此, 一般情況下, 氮肥添加到土壤, 植物的根冠比會(huì)降低[31]。綜上可以得出, 土壤氮素的變化會(huì)影響到巖溶區(qū)固氮植物的生長(zhǎng)以及在養(yǎng)分吸收的生物量分配。
3.2 氮添加對(duì)巖溶區(qū)固氮植物固氮策略的影響
植物的充足營(yíng)養(yǎng)、 生物量增長(zhǎng)和分配狀況會(huì)影響到植物的適應(yīng)性策略。研究發(fā)現(xiàn), 添加氮肥情況下, 巖溶區(qū)兩種固氮植物的固氮速率顯著低于不添加氮肥的情況(圖3)。在生態(tài)系統(tǒng)中, 固氮植物能夠形成根瘤菌, 將空氣中的氮?dú)夤潭ü┲参锷L(zhǎng)利用或輸入到土壤環(huán)境中。固氮植物的固氮速率與土壤環(huán)境中可利用氮水平是可以相互反饋的, 當(dāng)土壤中可利用氮水平較低時(shí), 植物會(huì)提高固氮速率, 反之則降低固氮速率[32-33]。例如, 榿木(Alnusincana)幼苗的固氮速率和根瘤生物量[34]和熱帶固氮植物(Ingapunctate)的共生固氮速率和根瘤密度[20]隨著氮添加濃度的增加而降低。原因可能是: 生物固氮是一個(gè)極其耗能的過(guò)程[32], 植物在缺氮的環(huán)境中才會(huì)重點(diǎn)利用這個(gè)策略, 即在氮素貧瘠環(huán)境中固氮植物加強(qiáng)與根瘤菌的共生作用, 提高固氮速率以緩解土壤氮素供應(yīng)的不足[35]。但是, 隨著土壤環(huán)境中可利用氮水平的提高, 固氮植物通過(guò)根系就可以吸收足夠的氮素滿(mǎn)足自身的生長(zhǎng), 減弱與根瘤菌的共生作用, 表現(xiàn)為較低的固氮速率[36]。這種固氮策略為兼性固氮策略。綜上可以得出, 本研究的兩種巖溶區(qū)固氮植物的固氮策略為兼性固氮策略。
土壤氮素變化導(dǎo)致的固氮速率的變化也與一系列的根系生物化學(xué)特征變化有關(guān)。一方面, 土壤氮素變化導(dǎo)致固氮速率與菌根侵染率發(fā)生變化。固氮植物的固氮速率與菌根的侵染率具有顯著的正相關(guān)關(guān)系(表1)。叢枝菌根真菌能夠促進(jìn)植物吸收氮磷養(yǎng)分, 已有研究表明, 陸地上80%的植物能夠與叢枝菌根形成共生關(guān)系[37], 尤其是養(yǎng)分貧瘠的條件下, 植物會(huì)增加與叢枝菌根的共生關(guān)系, 促進(jìn)植物對(duì)土壤氮磷養(yǎng)分的吸收[38], 從而緩解植物生長(zhǎng)受養(yǎng)分的限制。但是, 在養(yǎng)分比較充足的條件下, 植物會(huì)減弱與叢枝菌根的共生關(guān)系, 降低根系的菌根侵染率, 減少能量消耗。本研究發(fā)現(xiàn), 盡管添加氮肥情況下香合歡根系的叢枝菌根侵染率顯著低于不添加氮肥的情況, 而木豆根系叢枝菌根侵染率的變化不大, 但是, 總體上兩種巖溶區(qū)固氮植物的叢枝菌根侵染率降低明顯(圖4b)。發(fā)生這種現(xiàn)象的原因可能是不同植物種對(duì)氮肥添加的濃度范圍靈敏性相異, 但是叢枝菌根侵染率與氮肥濃度之間的變化趨勢(shì)還不夠清楚, 未來(lái)的研究應(yīng)布設(shè)一系列氮肥添加濃度梯度對(duì)固氮植物菌根侵染率的影響研究。
另一方面, 土壤氮素變化導(dǎo)致磷酸酶活性變化。磷酸酶是植物獲取土壤磷素的一種重要策略, 能夠把土壤中的有機(jī)磷轉(zhuǎn)化為無(wú)機(jī)磷, 有利于植物磷素的吸收[39-41]。本研究中, 氮肥添加顯著提高了木豆根系的酸性磷酸酶活性而稍微降低了香合歡根系的酸性磷酸酶活性, 總體上則是氮肥添加提高了根系的酸性磷酸酶活性(圖3), 出現(xiàn)此現(xiàn)象可能與植物的生物量和養(yǎng)分含量的提高有關(guān): 土壤中氮素增加導(dǎo)致植物的生物量提高, 養(yǎng)分的吸收量提高, 充足的氮素供應(yīng)使得植物對(duì)磷素的需求持續(xù)增大, 可能會(huì)導(dǎo)致有效磷的供應(yīng)缺乏。一般情況下, 土壤磷素有效性較低的環(huán)境將有利于刺激植物分泌酸性磷酸酶[42-44]。以上研究表明, 植物根系磷酸酶活性除了受環(huán)境中土壤磷素含量影響外, 還與植物種適應(yīng)環(huán)境養(yǎng)分含量變化的適應(yīng)策略有關(guān)。在巖溶地區(qū), 植被演替早期的土壤氮含量較低, 植物受到氮限制, 而隨著植被恢復(fù)的正向演替, 土壤的氮含量逐漸提高, 磷含量提升不明顯, 演替后期的植物更容易受到磷限制[9, 45]。氮添加控制試驗(yàn)?zāi)M了巖溶地區(qū)植被恢復(fù)土壤氮含量的變化, 氮素的充足使得植物更容易缺磷, 從而促使磷酸酶活性的提高。
3.3 巖溶區(qū)生態(tài)恢復(fù)中固氮植物的潛在選擇
不添加氮肥情況下, 香合歡的根冠比、 根系的氮磷含量與莖、 葉的氮含量、 固氮速率、 菌根侵染率、 根系磷酸酶活性高于木豆。這種現(xiàn)象表明, 在巖溶地區(qū)植被演替早期土壤氮含量較低的情況下, 香合歡比木豆更容易適應(yīng)土壤的氮限制。此外, 添加氮素使得木豆的菌根侵染率和根系磷酸酶活性高于香合歡, 表明在巖溶地區(qū)隨著植被恢復(fù)后期土壤氮素的升高, 木豆比香合歡更能適應(yīng)植被恢復(fù)后期的土壤環(huán)境。固氮植物可以使得生態(tài)系統(tǒng)中土壤氮含量升高, 為其他生物物種提供氮素資源。巖溶生態(tài)系統(tǒng)是一個(gè)生態(tài)脆弱、 養(yǎng)分缺乏、 植被可持續(xù)發(fā)展差的生態(tài)系統(tǒng), 其生態(tài)恢復(fù)應(yīng)需進(jìn)行人工干預(yù)。基于以上結(jié)果和討論, 在巖溶地區(qū)生態(tài)恢復(fù)早期進(jìn)行人工干預(yù)時(shí)選擇香合歡作為植物群落配置的物種更為合適, 而在生態(tài)恢復(fù)后期選擇木豆作為植物群落配置的物種更為合適。
4 結(jié) 論
本文通過(guò)控制性試驗(yàn)研究發(fā)現(xiàn), 氮肥添加有利于提高巖溶區(qū)固氮植物的總生物量與土壤銨態(tài)氮和有效磷含量, 但是降低了植物的固氮速率。植物共生固氮速率受環(huán)境氮素濃度的影響, 高氮會(huì)抑制固氮植物的固氮速率, 巖溶區(qū)固氮植物的固氮策略為兼性固氮策略。
不添加氮肥情況下, 香合歡的根冠比、 根系的氮磷含量與莖、 葉的氮含量、 固氮速率、 菌根侵染率、 根系磷酸酶活性高于木豆; 添加氮素情況下, 木豆的菌根侵染率和根系磷酸酶活性高于香合歡。可見(jiàn), 在巖溶地區(qū)生態(tài)恢復(fù)早期進(jìn)行人工干預(yù)時(shí)選擇香合歡作為植物群落配置的物種更為合適, 而在生態(tài)恢復(fù)后期選擇木豆作為植物群落配置的物種更為合適。但是, 巖溶地區(qū)不同植被恢復(fù)階段生長(zhǎng)有大量的固氮植物, 要揭示不同固氮植物的固氮策略以及為植被恢復(fù)提供適宜不同恢復(fù)階段、 不同地區(qū)、 不同坡位的物種配置方案, 還需要廣泛、 深入地開(kāi)展野外定位監(jiān)測(cè)和多因子控制試驗(yàn)的綜合性研究, 這是今后工作的一個(gè)重要方向。
猜你喜歡
少兒科學(xué)周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學(xué)閱讀指南·低年級(jí)版(2021年3期)2021-03-19 06:12:40
小天使·二年級(jí)語(yǔ)數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學(xué)苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫(huà)刊)(2017年11期)2018-01-09 19:15:14
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學(xué)周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
