二氯乙腈對大腸桿菌基因表達的影響
2021-03-30 06:18:50任杰輝西安理工大學西北旱區生態水利國家重點實驗室陜西西安710048
中國環境科學 2021年3期
李 冬,程 文,秦 璐,任杰輝,鄭 興(西安理工大學,西北旱區生態水利國家重點實驗室,陜西 西安 710048)
飲用水消毒是飲用水安全的重要保障,避免了霍亂、痢疾和傷寒等常見疾病的擴散傳播[1].然而,隨著消毒工藝的進步與發展,消毒后產生了許多種類的消毒副產物(DBPs),研究發現多種癌癥(如結直腸癌、膀胱癌等)的發生與通過飲用水接觸DBPs直接相關[2].因此,深入研究并控制飲用水消毒后產生的DBPs,對居民飲水健康具有重要價值.
消毒副產物主要包括三鹵甲烷、鹵乙酸、鹵乙腈、亞硝胺等類物質,這些物質常常存在于消毒后的飲用水中.相關研究[3-4]指出飲用水中檢出 700余種DBPs,對人體健康存在潛在危害,但僅有一少部分DBPs受到監管或規范,如三鹵甲烷及鹵乙酸.因而,掌握各類DBPs的危害及毒性作用,對DBPs的管控具有重要價值.近年來由于含氮廢水排放及氯胺消毒使用的增加,含氮消毒副產物(N-DBPs)逐漸在飲用水中被檢出.已有研究[5-6]表明 N-DBPs比含碳消毒副產物(C-DBPs)具有更強的三致效應,因而促使N-DBPs備受關注[7-9].鹵乙腈類消毒副產物(HANs)是最常檢測到的 N-DBPs[10],也是供水中已知的 10種DBPs中最有害的,占總毒性的45-83%[11].二氯乙腈(DCAN)作為一種典型的 HANs,與大多數N-DBPs相比,因其具有在水處理廠中檢出率高[12]、含量高[13]、致癌致畸性強等特點,逐漸受到廣泛關注.目前的研究中發現 DCAN會誘導小鼠皮膚產生腫瘤,甚至導致小鼠 DNA 損傷加劇細胞凋亡[14-15];使中國倉鼠卵巢細胞密度降低及 DNA損傷[16].然而,關于 DCAN暴露的毒理學特性研究較少,導致對DCAN管控的研究受到限制.
目前的毒性檢測一般是以誘導損傷檢測為主,包括Ames試驗、染色體畸變試驗、體外彗星試驗、內源基因突變,以及嚙齒動物和水生生物體內致癌性生物實驗[17-22].而大多數體外檢測只檢測單一類型的損傷,提供細菌或其他生物體經歷的整體性生理變化信息非常有限;體內檢測方法操作繁瑣、檢測周期長、有違 3R(減量化、再利用、再循環)原則,所以利用高通量和耗時短的分析方法來評估大量環境污染物的毒性更加迫切.近年來以高通量技術為研究手段的毒理基因組學不斷發展為環境污染物的毒性評估提供了新的研究工具[23],使研究人員能夠在特定條件下,對環境污染物在基因水平上的毒性作用進行較為全面的分析.因其可對環境應激源暴露作出應答,具有速度快、高通量等優勢被廣泛應用[24].
本研究采用以毒理基因組學為基礎的實時基因表達譜方法,將 DCAN作用于特定 E.coli,對其所誘導的應激反應進行分析,探究其毒理作用的基因機制,為研究其對人類健康可能產生的直接或間接毒性作用提供新思路.
1 材料與方法
1.1 主要試劑及儀器
DCAN(純度 98%+,上海阿拉丁生化科技股份有限公司),M9低鹽培養基(濃度:5×,上海瑞楚生物科技有限公司),二甲基亞砜(DMSO,分析純,廣東省化學試劑工程技術研究開發中心),全自動移液工作站(epMotion 5075t,德國Eppendorf AG)、微孔板細胞成像系統(Cytation5,USA Bio-Tek).DCAN 不溶于水,用0.01% DMSO 作助溶劑.因在最大耐受濃度研究中,以DMSO為空白對照,考察了對菌株基因表達的影響,故將DMSO濃度設定為0.01%,確保對實驗結果無影響.
1.2 實驗受試菌株
研究中采用的菌株為綠色熒光蛋白(GFP)轉染的大腸桿菌基因文庫(菌株),E.coli K12,MG1655(含低拷貝質粒pUA66或pUA139、卡那霉素抗性基因及快速折疊的gfpmut2基因)包含不同的應激反應基因啟動子[25].
1.3 實驗方法
以 DCAN在水中的最大溶解度為上限濃度進行預實驗,測定其生長抑制情況(OD600),確定細胞存活率≥95%時的濃度,即最大耐受濃度(EC5)為1.429×10-3mg/L,以EC5為實驗濃度上限依次向下稀釋10倍進行實驗研究.
菌株置于96孔板中培養,37℃條件下靜止過夜培養;以1×M9培養基將菌液按體積比1:5稀釋,轉接至 384孔板;將 384孔板中的菌液在 37℃下培養5~6h,使光密度 OD600[26-29]達到早期指數生長階段(OD600讀數約0.2);培養后的384孔板中添加不同濃度的DCAN;為了測定DCAN誘導的基因轉錄水平效應,將 384孔板置于微孔板細胞成像系統中,同時測量光密度(OD600,細胞生長)和熒光讀數(GFP水平,Ex:45nm,Em:528nm),以 2h為暴露時間,每 5min測定讀數,實驗設置3組平行樣.
1.4 實驗數據處理
以5次移動平均法處理GFP和OD數據,然后采用對照組進行校正.將GFP信號歸一化,以P=GFP/OD計算,并分別在暴露于和未暴露于DCAN的情況下對背景進行校正(不含GFP的E.coli菌株).基因表達的改變,也被稱為誘導因子 I[21],以基因在化學樣品暴露的實驗條件下及在沒有任何化學樣品暴露的對照條件下的比率表示.采用每個時間點I的自然對數ln I進行進一步數據分析,ln I值:中性=0、上調>0、下調<0.同時,通過整合誘導因子 I隨時間變化來計算轉錄效應水平指數(TELI),用于量化DCAN存在下基因表達水平的改變,指示給定基因在一定暴露期內的累積轉錄效應[30-31],通過識別和分析與特定應激反應途徑相關的基因變化,探究DCAN的毒性作用模式.
1.5 自組織映射(SOM)分析
SOM 常用于基因表達數據的探索性分析,目前已被有效地用于共表達基因組的發現或化學物質的分類研究中[32].為從濃度-時間-基因三維數據集中發現具有相似表達水平的基因,識別 DCAN的毒性作用模式,采用微陣列軟件套件 MeV 4.9[33]進行SOM分析及可視化,參數設置如表1所示.
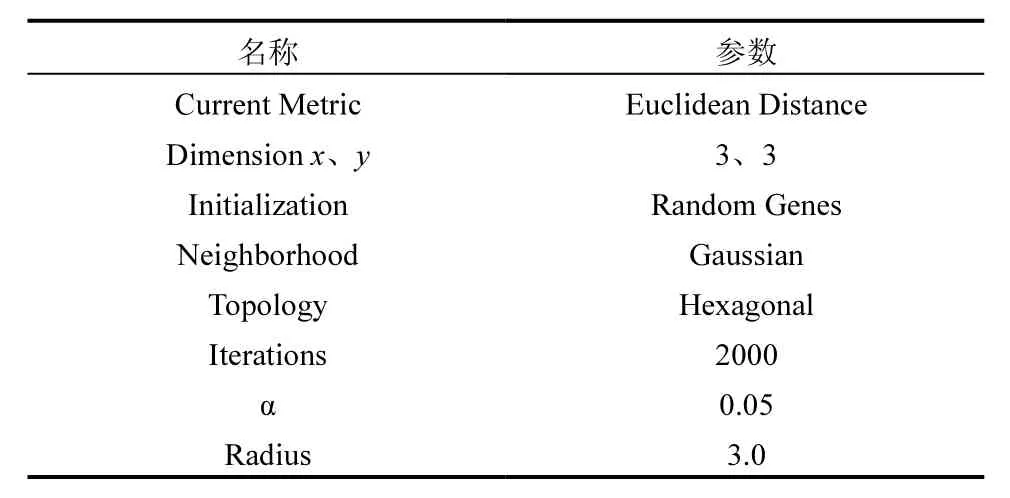
表1 SOM分析參數設置Table 1 Parameter setting of SOM analysis
1.6 DCAN毒性終點確定
轉錄效應水平指數(TELI)是最近開發的毒理基因組學數據解釋指數,且與表型毒性研究中所采用的終點指標存在一定的相關性[29].TELI值分析方法考慮到化合物暴露導致的表達改變基因的數量、特性、程度及時間序列,可以確定單個基因或代表特定途徑或整個應激反應的變化,故采用TELI值為基礎的數據擬合劑量-反應曲線,進行分子毒性終點的推導.在毒理基因組學中,以TELI值法表示的基因表達水平達 1.5時的濃度用 TELI1.5表示,反映了亞急性毒性中毒性作用的最低濃度,故采用graphpad Prism 5.0軟件以四參數非線性回歸模型擬合TELI劑量-效應曲線,由此得出各應激途徑的毒性終點TELI1.5.
2 結果與討論
2.1 DCAN對E.coli實時基因表達的影響
通過在6個濃度梯度的DCAN中暴露E.coli菌株120min的實驗方法,獲得時間及濃度兩個維度下的基因表達譜,結果如圖 1所示,圖中提供了有關各種應激反應途徑和功能基因的詳細表達信息,根據應激基因的主要功能和參與的不同應激機制程度分為5個功能組,包括DNA應激(含SOS反應)、氧化還原應激(含解毒和氧化還原平衡)、蛋白質應激、膜(脂質應激和膜轉運體)和普通應激[15].從圖譜中可以看出,DCAN的基因表達譜較為復雜具有動態性,隨時間及濃度不同,不同基因產生了不同的表達結果,如otsB在1.429×10-4mg/L時表達上調趨勢明顯,為正向調控;低濃度暴露促進了 mutS的表達;uvrY在不同濃度中表達水平不斷變化;motA在所有濃度中都呈現出上調趨勢;而 umuD僅在最高實驗濃度中表達上調趨勢明顯;其他一些基因表達呈現出短時效應及延遲效應.圖譜中還顯示大多數普通應激基因表達呈上調趨勢,說明 DCAN暴露過程中對細胞內的穩態環境可能產生嚴重干擾;與其他類型應激相比,大部分與DNA應激相關的基因表達水平變化比較明顯,這些基因表達水平變化可能與生物體內多個基因的激活和信號通路變化相關.

圖1 DCAN暴露下E.coli菌株的實時基因表達譜Fig.1 Real-time gene expression profile of E.coli exposed to DCAN
2.2 DCAN對E.coli毒性作用分析
采用SOM聚類方法,根據基因的時間表達模式識別出9類基因簇(圖2a).第I類包含基因數目達35個,約占總基因數的32%,且多與DNA損傷及氧化還原應激相關,基因表達呈下調趨勢;而第Ⅳ類基因則在 40min后從中性狀態轉變為下調狀態;DCAN暴露下,第Ⅴ與Ⅵ類中的基因并未產生表達改變.與其他基因簇相比,第Ⅸ類有 28個與普通應激、膜及DNA損傷應激相關的基因均處于上調狀態.與對照組相比,參與DNA修復的SOS系統中uvrA、ybfE、nfo和sbmC等基因表達水平較高,屬于上調基因,其中 uvrA是檢測損傷并啟動下游內質網級聯反應的關鍵蛋白,優先與受損DNA結合[34],修復多種DNA病變過程,與核苷酸切除修復(NER)途徑相關,它的過度表達表明它在修復各種各樣的損傷中起著積極的作用;核酸內切酶nfo在雙脫氧核糖核酸中催化單鏈斷裂,參與堿基切除修復(BER),它的上調表明DCAN暴露使DNA鏈發生斷裂,同時nfo基因對過氧化氫和其他活性氧(ROS)等氧化劑有反應,說明DCAN通過氧化應激進而誘導 DNA損傷的發生.RecE是E.coli經典RecET重組系統的一部分,與游離的雙鏈DNA(dsDNA)末端結合,采用與I型外切酶類似的機制,過程性消化 dsDNA,從而使受損DNA鏈的核苷酸重新定位在活性位點內的50′末端鏈上[35],對DNA進行修復,RecE表達上調同樣說明DCAN會促進E.coli DNA損傷.SOM分析中存在多個 DNA修復相關基因的啟動子活性改變表明DCAN引起DNA損傷,該結果與之前的研究結果一致[15,36-37].Lipscomb等[38]和 Ahmed 等[39]的研究也指出DCAN會誘導DNA鏈斷裂,進而造成了DNA損傷.在DCAN暴露下SOS應急系統中的基因也存在表達抑制的情況,但并不是所有的 SOS基因都參與生理性 DNA 修復途徑[40].因此,其對不同基因毒性的參與和活性有待進一步研究.
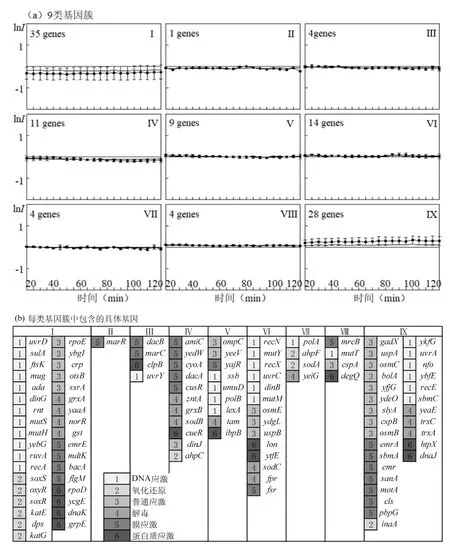
圖2 DCAN的SOM圖Fig.2 The SOM diagram of DCAN
除SOS應急基因外,22個與其他應激相關的基因包括耐藥/敏感基因、普通應激基因和解毒機制基因在 DCAN暴露條件下呈上調趨勢,其中耐藥基因cmr(又稱 mdfA),是轉運蛋白超家族(MFS)的成員,其產物是一種多藥外排蛋白,其過度表達可導致廣泛化合物的耐藥性;motA與鞭毛合成和運動功能有關,其表達改變會影響 E.coli代謝功能;emrA、sbmA、sanA、pbpG與膜轉運蛋白及排泄有關,cls涉及細胞膜及磷脂的合成.上調基因中還包括氧化還原應激中的inaA、yeaE、trxC、trxA基因,yeaE、trxC、trxA與解毒機制相關,DCAN暴露觸發了解毒機制,表明氧化還原應激是重要的毒性作用模式,Esmat等[15]的研究發現 DCAN存在條件下通過氧化應激及神經退行性病變進而對胎鼠腦組織產生不良影響.與細胞死亡有關的 slyA表達也表現出上調趨勢,基因cyoA在40min后表達呈下調趨勢,說明DCAN的存在干擾了電子傳遞可能影響細胞內 ATP水平,對能量代謝產生影響.許多與普通應激相關的基因在DCAN暴露中存在過表達的情況,表明細胞生物化學和物理穩態可能受到干擾,但在現有研究中未發現相關報道,表明普通應激可能是 DCAN潛在的毒性作用模式.其他功能基因暴露在 DCAN中表達水平發生了改變,說明 DCAN對其他類型的應激反應也存在一定影響.
2.3 DCAN毒性終點確定
劑量是決定外源化學物對機體造成損害作用的主要因素之一[41],劑量的多少決定了毒性的強弱.因此,為了獲取DCAN定量分子毒性終點,采用TELI值法對不同功能基因的表達水平進行計算,獲得了不同濃度下不同途徑應激反應的毒性變化,結果如表2所示,發現DNA應激及普通應激的TELI值隨濃度增大呈增加趨勢,表明毒性隨濃度增加而增加,但在其他應激反應中沒有呈現較好相關變化趨勢.為進一步探究濃度與毒性作用的關系,采用四參數非線性回歸模型擬合了DCAN 6個濃度梯度的劑量-效應關系曲線,擬合結果如圖3所示.不同應激反應表現出特征性的“S形”曲線或“S形”曲線的一部分.該研究結果與以劑量的對數對藥效作圖,多呈 S型曲線的理論[42]相一致.從劑量效應曲線中可以觀察到 DNA損傷是 DCAN誘導的主要毒性作用模式,該結果與SOM分析結果一致.在DCAN最低濃度暴露下,DNA損傷非常明顯,其他各類型應激反應變化水平呈現氧化還原應激>普通應激>膜應激>蛋白質應激的趨勢,說明普通應激和氧化應激損傷也是DCAN誘導的重要毒性作用模式.這些結果均表明DCAN對重組基因的大腸桿菌存在潛在毒性作用.
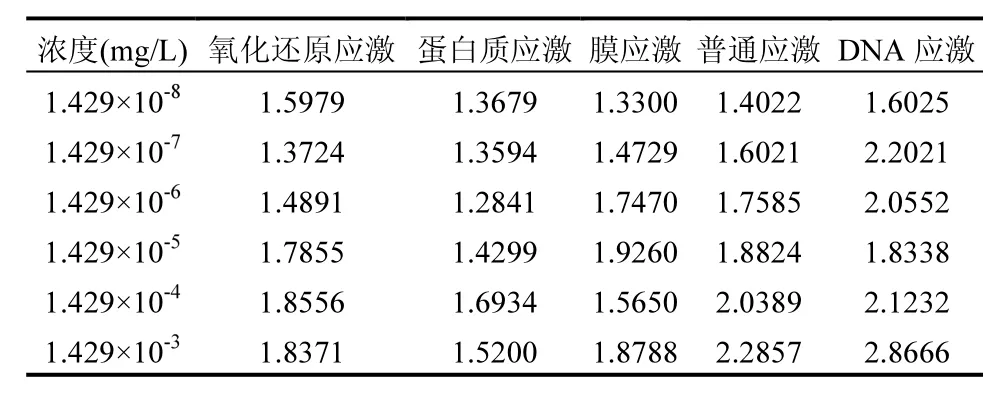
表2 DCAN暴露下不同應激反應毒性變化Table 2 Toxicity changes of different stress responses to DCAN

圖3 基于TELI值的DCAN劑量-效應曲線Fig.3 Dose-response curves of DCAN based on TELI values
通過實驗分析及模型計算得到DCAN的不同應激反應類型毒性終點,結果如圖4所示(因普通應激擬合后數據范圍較為寬泛,無法得出TELI1.5具體數據值).從圖中可以看出,DNA 應激的 TELI1.5為6.54×10-9mg/L,與其他應激反應的 TELI1.5相比數量級最小,濃度小而產生的基因表達水平與其他應激基因表達一致,說明 DNA損傷是主要的毒性作用模式.

圖4 DCAN的毒性終點Fig.4 The toxicity endpoint of DCAN
3 結論
3.1 E.coli暴露于DCAN中,獲得的實時基因表達譜復雜又具有動態性,不同基因隨時間及濃度改變產生了不同的表達結果.
3.2 在高實驗濃度 DCAN 暴露條件下,多個參與SOS調節的基因啟動子活性發生改變,從而引起DNA損傷,并通過正向調控inaA、yeaE、trxC、trxA基因加劇氧化還原應激,同時與普通應激相關的基因的過表達也說明E.coli細胞生物化學和物理穩態可能受到干擾.
3.3 DCAN劑量-效應關系曲線表現出特征性的“S形”曲線或“S形”曲線的一部分,且與其他應激反應的毒性終點相比,DNA應激的 TELI1.5數量級最小,因而DNA損傷是主要的毒性作用模式.
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
發明與創新(2016年38期)2016-08-22 03:02:52
