胚胎絨毛膜對微塑料顆粒與鎘聯合作用的影響
2021-03-30 06:18:52段鑫越關文玲程昊東戴媛媛端正花天津理工大學環(huán)境科學與安全工程學院天津0084天津市水產研究所天津00南開大學環(huán)境科學與工程學院天津0050
中國環(huán)境科學 2021年3期
段鑫越,關文玲,程昊東,戴媛媛,汪 磊,端正花,*(.天津理工大學環(huán)境科學與安全工程學院,天津 0084;.天津市水產研究所,天津 00;.南開大學環(huán)境科學與工程學院,天津 0050)
塑料是合成或半合成的有機聚合物.因其具有優(yōu)良的物化性能,被廣泛應用于工業(yè)生產和日常生活中.2014年我國就成為全球塑料消費量最大的國家,占全球塑料消費量的 26%(Plastics—the Facts 2015).因此我國的塑料污染問題尤為嚴重.環(huán)境中的塑料垃圾可能會持久存在并被物理破碎為塑料微粒,即微塑料[1-2].近年來我國環(huán)境中微塑料的研究報道日益增加[3],尤其微塑料作為塑料污染物在水環(huán)境中廣泛存在[4].周倩等[5]觀測到渤海地區(qū)每年通過大氣沉降獲得的微塑料量約為 2.33×1013個,相當于約0.9~1.4t.Zhao等[6]調查發(fā)現長江口和其附近海域地區(qū)水體表面漂浮的微塑料豐度分別為(4137±2461),(0.167±0.138)個/m2.此外,食用海鹽中也檢測到了微塑料顆粒的存在.海鹽和淡水鹽中微塑料的含量分別為 550~681,43~364個/kg[7].微塑料的量級與許多動物的食物粒徑相似,使得微塑料易于被動物誤食[8].目前有限的調查和模擬結果均證實,這些廣泛存在的微塑料可以進入部分生物體內,但是微塑料及其附著的污染物在生物體內的分布與毒性機制還缺乏系統(tǒng)地探討.
水體中重金屬污染是全球水環(huán)境面臨的嚴峻問題[9],并且重金屬對機體產生的氧化性壓力是其毒性作用機制的重要方面[10-11].海洋水體環(huán)境認為是微塑料的一個重要的匯.漂浮在海洋水體中的微塑料與重金屬離子之間的作用關系必須引起重視.不同材質微塑料對重金屬的較強吸附能力已被證實[12-13].微塑料與重金屬復合污染的協(xié)同效應也被報道[14-15].如 Luís等[16]發(fā)現 5μm 的聚苯乙烯(Polystyrene,PS)顆粒與 Cr復合污染能夠增強暇虎魚仔魚機體脂質過氧化水平和AChE活性的抑制率.但是這些研究注重的都是能進入機體的微塑料與重金屬的聯合效果.
絨毛膜是一層特殊的生物結構,在幼魚孵化前一直包圍著胚胎.絨毛膜上具有大量直徑達 500nm的孔[17],這些孔允許必要的氧、鹽離子和一定尺寸的納米顆粒通過.不同粒徑的外源化合物在胚胎絨毛膜上的穿透性是不一樣的[18].絨毛膜的存在降低了胚胎對外源化合物的敏感性[19-20],但絨毛膜本身與外源化合物交互作用后的毒性效應值得引起關注[21].水體中重金屬離子與微塑料聯合暴露時,微塑料載帶的重金屬離子通過胚胎絨毛膜的含量和生物可利用性可能會發(fā)生變化,從而最終改變水體中重金屬離子對胚胎發(fā)育的毒性.Lee等[22]認為50nm PS納米顆粒能夠穿透胚胎絨毛膜并與金離子發(fā)生協(xié)同毒性效應,而500nm PS納米顆粒不能穿透胚胎絨毛膜,但能增強金離子在胚胎絨毛膜表面的吸附,降低了金離子在胚胎體內的含量.那么這些不能穿透絨毛膜的微塑料顆粒與重金屬聯合作用后,重金屬在胚胎體內的濃度可能會降低,最終體現出來的一定是拮抗效應嗎?這個問題在目前研究中未見報道.
本課題組前期研究表明,100nm和70~250μm的PS顆粒都不能穿透胚胎絨毛膜,但是會吸附在絨毛膜表面,從而導致絨毛膜空間內形成低氧微環(huán)境,使得胚胎表現出缺氧的毒性效應[21].據此,本研究采用水體直接暴露的方式,以100nm和70~250μm兩種粒徑的PS顆粒為研究對象,探討這些不能穿透胚胎絨毛膜的微塑料顆粒對重金屬毒性的改變,對比微塑料顆粒粒徑大小造成的它們與重金屬聯合作用效果的差異,并進一步基于代謝組學技術探討了這種差異產生的機制.
1 材料與方法
1.1 材料
氯化鎘(CdCl2)購于Sigma-Aldrich(純度>99.9%,美國).納米PS顆粒懸浮液(100nm,n-PS)和微米PS微塑料顆粒(70~250μm,μ-PS)分別購于紐邦生物科技有限公司(深圳,中國)和億能塑料有限公司(東莞,中國),具體形態(tài)如圖1所示.

圖1 納米(100nm)(A)和微米(70~250 μm)(B)PS顆粒的形態(tài)Fig.1 Morphology of PS particles with nano(100nm)(A)and micro(70~250μm)(B)sizes
成年斑馬魚購自天津市花鳥魚蟲市場.飼養(yǎng)用水經生物過濾器過濾并充分曝氣.每日喂食新鮮紅蟲 2次,于(26±1)℃光/暗(14/10)h周期培養(yǎng) 1月后,開始收集魚卵.用立體顯微鏡(尼康,日本)挑選出發(fā)育正常的魚卵備用.
1.2 實驗設置
稱取一定量的氯化鎘(CdCl2)和微塑料固體溶于胚胎營養(yǎng)液[23]中,稀釋到需要的濃度.根據預實驗的結果,本試驗中 Cd2+的暴露濃度為 1mg/L,微塑料濃度為 20mg/L.本試驗設置空白對照組 Ctr、Cd,以及μ-PS/n-PS和Cd的聯合暴露組共計4組,分別標記為Ctr、Cd、μ-PS+Cd和n-PS+Cd.溶液在使用之前新鮮配置,并采用超聲波水浴超聲處理10min.
1.3 檢測和表征
1.3.1 斑馬魚胚胎絨毛膜和體內 Cd含量的測定實驗包含Ctr、Cd、μ-PS+Cd和n-PS+Cd共計4組,每組設6個平行.在100mL燒杯中放入100枚發(fā)育良好的受精卵(4hpf,hours post fertilization),分別加入上述 4組暴露溶液,放置培養(yǎng)箱中培養(yǎng).參考 Lee等[22]的方法,用手術剪和鑷子在顯微鏡下人工剝離即將孵化(大約60hpf)的胚胎和絨毛膜各50個,分別作為胚胎和絨毛膜的一個樣本,在蒸餾水中反復沖洗至少 3次,小心放入 15mL離心管內,-80℃保存.而后加入10mL硝酸消解后定容至12mL,過微孔濾膜后,利用 ICP-MS(Agilent 7500a,美國)測定胚胎絨毛膜和體內Cd的含量.
1.3.2 斑馬魚胚胎代謝組學分析 實驗包含 Ctr、Cd、μ-PS+Cd和n-PS+Cd共計4組.挑選50枚發(fā)育良好的受精卵放入 100mL的燒杯中,加入空白和暴露溶液50mL,放入人工氣候箱進行培養(yǎng) 60hpf.而后將胚胎取出,用雙蒸水仔細清洗后,轉移到 1.5mL離心管中,盡量吸干水分后凍干,每組設3個平行.
在每管中添加 1350μL預冷萃取混合物(甲醇/氯仿=3:1,體積比)和 30μL 內標物(L-2-氯苯丙胺,1mg/mL),混勻30s后勻漿;冰浴超聲萃取5min后,11000g、4℃離心15min;將300μL上清液轉移到新鮮試管中.從每個樣品中取出 100μL制備質量控制(QC)樣品.QC樣本操作同實驗.樣本在真空濃縮器中蒸發(fā)后,添加20μL甲氧基胺化鹽酸鹽(20mg/mL),然后在 80℃下培養(yǎng) 30min,并在 70℃下用 30μL BSTFA試劑(含1% TMCS,V/V)衍生1.5h.將樣品逐漸冷卻至室溫.吸取 2μL衍生化后的反應液用自動進樣器進行GC-TOFMS(Gas chromatography tandem time of flight mass spectrometry,Agilent 7890,美國)分析.
數據采集以 scan模式獲得總離子流色譜圖,以峰面積定量,利用內標進行歸一化.使用ChromaTOF軟件(V4.3x,LECO,美國)對質譜原始數據進行峰提取、基線矯正、解卷積、峰對齊和峰積分等分析,對單個峰進行過濾以去除噪音.基于四分位數距對偏離值進行過濾,只保留單組空值不多于 50%或所有組中空值不多于50%的峰面積數據.使用SIMCA軟件(V14.1,Sartorius Stedim Data Analytics AB,瑞典)進行主成分分析(principal component analysis,PCA).所有代謝物在LECO—Fiehn Rtx5數據庫(LECO,美國)通過質譜匹配及保留時間指數匹配進行物質信息搜索整理.通過KEGG(Kyoto Encyclopedia of Genes and Genomes)注釋分析找到所有差異代謝物參與的通路,并對差異代謝物所在通路進行綜合分析(包括富集分析和拓撲分析),進一步篩選與代謝物差異相關性最高的關鍵通路,并用氣泡圖的形式表示.
1.3.3 微塑料與Cd聯合對斑馬魚胚胎的毒性表征將Ctr、Cd、μ-PS+Cd和n-PS+Cd上述4組暴露溶液加入到直徑10cm玻璃表面皿中,每個表面皿加入50mL試液和 20枚挑過的發(fā)育良好的受精卵,放入(26±1)℃光照培養(yǎng)箱中持續(xù)暴露 72h.在倒置顯微鏡(ZEISS Axiovert 200mol/L,德國)下記錄不同暴露組胚胎24和48hpf的血流速度和心跳次數,以及胚胎72hpf的孵化抑制率.實驗重復3次,結果用均值±SD表示.
效應指標計算方法如下:
血流速度=顯微鏡軟件下測量斑馬魚胚胎血管長度,用秒表記錄血液流經血管時間,血流速度為血管長度與血液流經血管時間的比值,μm/s;
心跳次數=顯微鏡下秒表記錄斑馬魚胚胎心臟1min跳動次數,times/s;
孵化抑制率=顯微鏡下以未破胚胎絨毛膜作為孵化抑制的標準,孵化抑制率為未孵化胚胎數占暴露總胚胎數的百分率(%).
1.4 統(tǒng)計分析
采用 Origin9.0軟件進行統(tǒng)計分析,結果用均值±SD表示;組間用student-t檢驗,P<0.05表示組間有顯著差異.
2 結果與討論
2.1 Cd在斑馬魚胚胎和胚胎絨毛膜中的蓄積
Cd單獨暴露組中,斑馬魚胚胎絨毛膜上 Cd的蓄積量為 3.82mg/g,此時胚胎體內 Cd的蓄積量為0.24mg/g(圖2).與μ-PS聯合暴露后,胚胎絨毛膜上和胚胎體內 Cd的含量分別為 11.35和 0.20mg/g,即μ-PS的吸附使得Cd在胚胎絨膜上的蓄積量顯著升高(P=0.01),而在胚胎體內的含量降低(P=0.001).
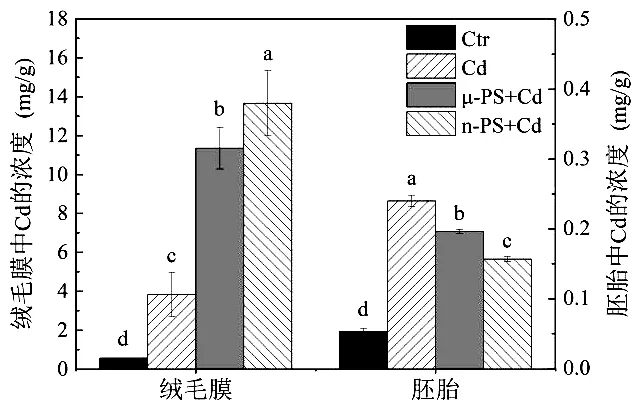
圖2 斑馬魚胚胎和絨毛膜中Cd的蓄積Fig.2 The contents of Cd in zebrafish embryo and chorion圖
n-PS+Cd暴露下,斑馬魚胚胎絨毛膜上吸附了更多的Cd,含量為13.66mg/g,約為Cd單獨暴露時的3倍(P=0.01).此時胚胎體內Cd的蓄積量也進一步降低,為 0.16mg/g.證明相對μ-PS,n-PS能夠更為顯著地降低溶液中Cd的生物可利用性(P<0.01).
2.2 代謝通路分析
2.2.1 主成分分析 本研究進一步檢測出各暴露組斑馬魚胚胎共計 230個有效差異代謝物,它們主要由糖類、氨基酸、脂肪酸和醇類等組成.主成分分析證明了空白對照組與各暴露組在代謝方面存在差異.如圖 3所示,Ctr、Cd、μ-PS+Cd與 n-PS+Cd暴露組組成不同的簇狀分布,其中μ-PS+Cd組在Ctr和Cd暴露組之間,n-PS+Cd組與各暴露組距離最遠.

圖3 不同處理組代謝成分的主成分(PCA)分析(n=3)Fig.3 The principal component analysis(PCA)of metabolic components in different treatment groups(n=3)
2.2.2 代謝通路分析 基于 KEGG 代謝通路的研究發(fā)現,μ-PS與Cd聯合暴露后,Cd的代謝通路無顯著變化(圖4A).但是,n-PS能顯著改變Cd誘導的胚胎代謝通路的表達(圖4B),主要表現在,與Cd單獨暴露相比,n-PS+Cd處理組中胚胎的甘油酯代謝[-ln(P)=2.823]、牛磺酸/次牛磺酸代謝[-ln(P)=1.917]、亞油酸代謝[-ln(P)=1.917]、甘氨酸/絲氨酸/蘇氨酸代謝[-ln(P)=1.888]和花生四烯酸代謝[-ln(P)=0.676]都顯著增強.
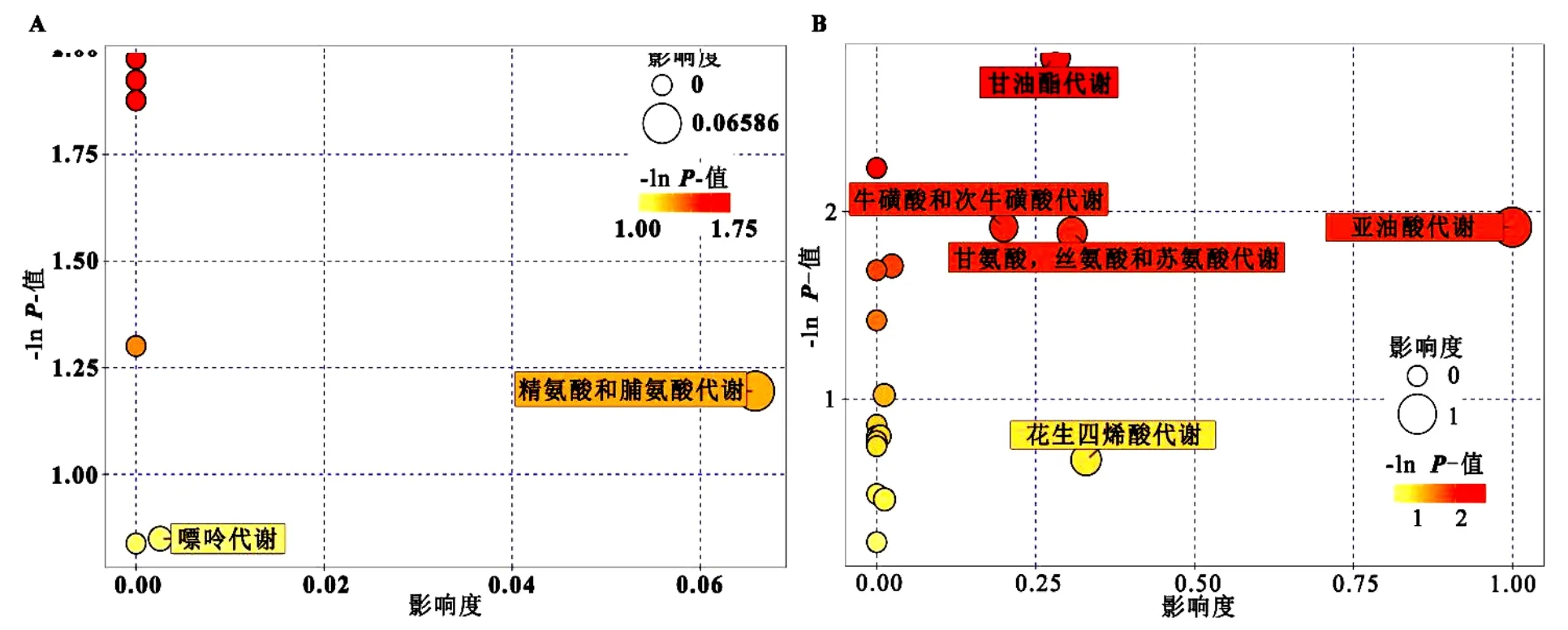
圖4 不同處理組代謝通路變化對比分析Fig.4 The comparative analysis of metabolic pathways in different treatment groups
2.2.3 氧化壓力相關代謝物表達分析 為探討不同粒徑PS顆粒對Cd造成的氧化壓力的影響,本研究進一步分析了牛磺酸和四種不飽和脂肪酸(油酸、亞油酸、花生四烯酸和 2-酮乙酸)的表達變化.牛磺酸的表達表現為 n-PS+Cd~Cd>Ctr>μ-PS+Cd;油酸、亞油酸和花生四烯酸的表達都表現為 n-PS+Cd>Cd>μ-PS+Cd>Ctr;2-酮乙酸的表達表現為 n-PS+Cd~Cd>μ-PS+Cd~Ctr(圖 5).因此,μ-PS 能夠一定程度上降低Cd的氧化壓力,而n-PS卻不能.

圖5 氧化壓力相關代謝物表達變化(n=3)Fig.5 Changes in the expressions of metabolites related to the oxidative stress(n=3)
2.3 斑馬魚胚胎發(fā)育毒性
2.3.1 單獨暴露對斑馬魚胚胎的發(fā)育毒性 本課題組前期研究表明,100nm和70~250μm的PS顆粒都不能穿透胚胎絨毛膜,但是會吸附在絨毛膜表面,從而導致絨毛膜空間形成內低氧微環(huán)境,主要的毒性表征為心跳加快、血流加速和孵化率抑制,并且n-PS的作用更為顯著[21].
本研究中1mg/L Cd單獨暴露下,斑馬魚胚胎血流速度自 24hpf就顯著降低,約為 301.70μm/s,與空白對照組相比降低了21.38%(P=0.02);72hpf孵化抑制率為33.33%(P<0.01).此外,1mg/L Cd暴露對斑馬魚胚胎發(fā)育表現出更為顯著的亞致死效應,具體毒性指標包括48hpf無黑色素沉積(8.33%,圖6B)、心包囊水腫(4.17%,圖 6C)和發(fā)育延緩(4.17%,圖 6D).這些毒性癥狀在μ-PS和n-PS單獨及與Cd的聯合暴露組中均未出現.

圖6 Cd與不同粒徑PS顆粒對斑馬魚胚胎發(fā)育的聯合毒性效應Fig.6 The combined toxic effects of Cd and PS particles with different sizes on the development of zebrafish embryos
2.3.2 微米PS顆粒對Cd毒性的影響 與Cd單獨暴露相比,μ-PS+Cd暴露組中胚胎 48hpf血流速度(P<0.01)和心跳次數(P=0.02)均顯著增大,并恢復到Ctr組的水平(P>0.05);胚胎72hpf孵化抑制率也降低到 18.75%(P<0.01).因此,μ-PS與 Cd聯合暴露表現為拮抗效應.
究其原因,一方面,20mg/L的μ-PS的毒性顯著低于1mg/L Cd的,對胚胎發(fā)育的毒性僅表現為缺氧引起的血流加速[21];另一方面,μ-PS對Cd有很強的吸附作用,降低了溶液中Cd的生物可利用性.前期研究表明μ-PS暴露 48h后,部分胚胎絨毛膜空隙被μ-PS顆粒所堵塞[21],阻止了 Cd向絨毛膜內斑馬魚胚胎的傳遞,使得絨毛膜上Cd的含量升高、胚胎體內 Cd的蓄積量降低(圖 2).氧化性壓力是外源化合物產生毒性的重要機制之一[24-26].盡管代謝通路表達變化不顯著(圖 4A),但是從氧化性壓力顯著相關的牛磺酸和四個不飽和脂肪酸的表達來看(圖5),μ-PS能夠在一定程度上通過降低Cd的生物可利用性,從而進一步降低Cd對胚胎的氧化壓力和毒性效應.
2.3.2 納米PS顆粒對Cd毒性的影響 與Cd單獨暴露相比,n-PS+Cd暴露組中斑馬魚胚胎24hpf時的血流速度和心跳次數反而升高,分別為 574.64μm/s和 55.55times/sec,分別高于空白對照組 49.88%(P<0.01)和 20.65%(P=0.02)(圖 6).這區(qū)別于 Cd 單獨暴露下胚胎血流速度的降低作用,對應于n-PS單獨暴露下24hpf時胚胎的心跳加速作用[21].
究其原因,一方面,n-PS與 Cd聯合暴露能夠進一步增強Cd對斑馬魚胚胎的氧化壓力.n-PS與Cd聯合暴露中,氧化壓力相關的代謝產物表達都顯著增強(圖5),其對應的牛磺酸/次牛磺酸代謝、亞油酸代謝都也顯著增強(圖4B),這與n-PS單獨暴露中的結果是一致的[21].谷胱甘肽是機體氧化應激次級防御系統(tǒng)中重要組成部分.甘氨酸/絲氨酸/蘇氨酸代謝在谷胱甘肽的合成中至關重要[27-28],它們在 n-PS+Cd暴露組中的表達也顯著增強,這也對應于n-PS單獨暴露中谷胱甘肽代謝通路增強[21].另一方面,相對Cd單獨暴露組,n-PS+Cd暴露組中脂肪酸代謝(甘油酯、亞油酸和花生四烯酸代謝)和氨基酸代謝活性進一步的增強,證明機體需求更多的能量以應對機體細胞的生命活動[29-30].它們在n-PS單獨暴露下也是增強的[21],而在μ-PS+Cd暴露組中卻無顯著變化.這些氧化壓力和能量代謝相關的差異代謝物可以作為n-PS胚胎發(fā)育毒性的標志物.
n-PS+Cd暴露組中,48hpf血流速度(P<0.01)和心跳次數(P=0.02)均顯著增大,并恢復到 Ctr暴露組的水平(P>0.05);胚胎 72hpf孵化率抑制率降低到26.39%(P=0.017).從這些指標來看,n-PS也表現為抑制了Cd的胚胎發(fā)育毒性.與μ-PS相比,n-PS與Cd聯合作用下,斑馬魚胚胎絨毛膜上 Cd的吸附量增多、胚胎體內Cd的濃度降低的更多(圖2).如果只考慮Cd的體內蓄積濃度對胚胎發(fā)育的影響,結果應該是n-PS+Cd組的毒性降低更多.但是從72hpf孵化率這個指標來看,n-PS對 Cd毒性降低的程度低于μ-PS(P=0.006).
一般地,孵化酶受到抑制、絨毛膜的機械性能變強、環(huán)境缺氧及胚胎自發(fā)運動變弱等因素都可能導致胚胎孵化延遲或失敗[31].前期研究表明n-PS單獨暴露下,24h后即可形成在胚胎絨毛膜表面形成密實的PS包裹層,導致胚胎缺氧并且絨毛膜機械性能增強,導致胚胎孵化率降低[21].并且如上所述,n-PS與Cd聯合暴露能夠進一步增強Cd作用下斑馬魚胚胎的氧化壓力及細胞的能量需求.因此,n-PS與 Cd的聯合暴露不僅僅是影響Cd的生物有效性,還體現出n-PS的自身效應.
微塑料由于其特殊的物理結構,對重金屬具有較強吸附能力[32],從而改變了重金屬的生物可利用性[33-34].由于污染物物理化學性能和受試生物結構和耐受性的差別,使得不同種類微塑料和重金屬的聯合作用效果存在爭議[35],并且目前的研究都側重于那些能夠被機體攝入的微塑料顆粒.例如,PS微塑料顆粒增加了金納米粒子對海洋微藻 T.chuii的毒性[36],但它們卻沒有改變金納米粒子對海魚仔魚的毒性[37].聚氯乙烯(Polyvinyl chloride,PVC)微塑料顆粒對Cd在貽貝中的生物可利用性無顯著影響[38].本研究中納米和微米級的 PS微塑料顆粒由于胚胎絨毛膜的阻隔作用,一定程度上降低了Cd對斑馬魚胚胎的生物可利用性和毒性效應,但是不同粒徑PS微塑料顆粒及其復合污染物,經由胚胎絨毛膜,會對水生生物早期發(fā)育造成不同的聯合毒性效果.因此,本研究能夠為微塑料在環(huán)境中的生物可利用性和毒性效應研究提供一些新的思考方向.
3 結論
3.1 70~250μm的PS顆粒被斑馬魚胚胎絨毛膜阻擋后,降低了 Cd在斑馬魚胚胎體內的蓄積,抑制了Cd的胚胎發(fā)育毒性.
3.2 100nm的PS雖然更顯著降低了Cd的生物可利用性,但是能夠增強Cd暴露下斑馬魚胚胎的氧化性壓力和細胞的能量需求,使得100nm PS顆粒降低Cd的毒性不如70~250μm PS顆粒顯著.
