新疆伊犁野生阿魏菇菌株的分離鑒定與培養特性
2021-03-31 09:52:22木古麗木哈西吾爾恩阿合別爾迪
微生物學雜志 2021年6期
關鍵詞:生長
木古麗·木哈西, 吾爾恩·阿合別爾迪, 劉 方
(1.南開大學 生命科學學院微生物學系 分子微生物學與技術教育部重點實驗室,天津 300071;2.伊犁師范大學 微生物資源保護與開發利用重點實驗室, 新疆 伊寧 835000)
阿魏菇(Pleurotusferulae)又名阿魏側耳、阿魏蘑,屬真菌門(Eumycota)擔子菌亞門(Basidiomycolina)擔子菌綱(Basidiomycetes)傘菌目(Agarieales)側耳科(Pleurotaceae)側耳屬(Pleurotus),是一種珍稀食、藥用菌。我國野生阿魏菇主要分布在新疆塔城和阿勒泰地區[1]。據文獻報道,阿魏菇含有豐富的多糖、蛋白質、脂肪酸、維生素和食用纖維,且富含多種礦質元素和微量元素[2]。因其生于阿魏植物根際,故其與阿魏草的醫療效果相似[3]。研究還發現,人工栽培阿魏菇的不同生物活性提取物具有抗氧化、抗炎、抗腫瘤、提高機體免疫力等功效[4-6]。1983年起,我國科學家相繼對分布于新疆托里縣、木壘縣、青河縣及石河子地區的野生阿魏菇進行了人工馴化栽培,同時進行遺傳分類研究[7-8]。截止目前,還沒有關于新疆伊犁野生阿魏菇菌株的分離鑒定與培養特性等方面的相關研究。在前期的伊犁野生阿魏菇資源野外調查中,筆者在分布區采集到兩種形態特征各異的野生阿魏菇子實體,如果能成功將其馴化并推廣,將具有較高的應用價值。因此,本研究以該兩種野生阿魏菇為研究對象,進行菌種分離純化與鑒定,并對優良菌株的培養條件進行初步篩選,研究結果將對伊犁野生阿魏菇菌種資源的保護利用及我國食用菌資源庫的豐富有重要意義。
1 材料與方法
1.1 材料
1.1.1 供試菌株 兩種伊犁野生阿魏菇子實體采自新疆伊寧縣喀什鄉新疆阿魏原生境保護區,地理位置分別為E82°07′32.7″、N43°44′48.7″和E82°06′40.3″、N43°44′28.4″,海拔分別為1 192 m和1 092 m。通過子實體組織分離及菌種純化,獲得伊犁野生阿魏菇菌株YL-A1和YL-A2,并保存于新疆伊犁師范大學微生物資源保護與開發利用重點實驗室。
1.1.2 培養基 ①PDA培養基:馬鈴薯200 g,葡萄糖20 g,瓊脂粉20 g,蒸餾水1 000 mL,用于菌種分離、形態特征觀察、菌種培養及保藏。②基礎培養基:馬鈴薯200 g,葡萄糖20 g,蛋白胨4 g,KH2PO41 g,瓊脂20 g,蒸餾水1 000 mL,用于確定菌絲體最適培養條件。
1.1.3 儀器與設備 立式壓力蒸汽滅菌器(YXQ-LS-75SII,上海博訊實業有限公司);隔水電熱恒溫培養箱(GHP-9050,上海艾牧生物科技有限公司);超凈工作臺(JJ-CJ-2FD,蘇州凈化);電子天平(TP-213,北京賽多利斯儀器系統有限公司);高速臺式離心機(HC-3018R,安徽中科中佳科學儀器有限公司);瓊脂糖水平電泳儀(G500320,上海生物工程股份有限公司);基因擴增儀(TC-48/H(a),杭州博日科技有限公司)。
1.2 方法
1.2.1 菌種分離培養 菌種分離純化采用組織分離法[9]。將野外采集到的阿魏菇子實體,用75%酒精和1%升汞依次進行表面消毒,在超凈臺下沿中線切開子實體,切取菌蓋與菌柄連接處內部組織數塊于PDA培養基平板上,25 ℃恒溫避光培養3~5 d,并多次進行菌絲轉接,最終獲得純培養物,并進行試管保種。
1.2.2 菌株鑒定 ①子實體形態特征及菌絲拮抗試驗:觀察菌株YL-A1和YL-A2野生子實體形態,記錄其表觀特征,參照相關文獻[10-11]進行形態學初步鑒定;觀察菌絲在平板上的生長情況,記錄菌落形狀、大小及色澤等特征。將活化后的菌株YL-A1和YL-A2,同時接種于一個PDA平板上相距2 cm左右的位置,3次重復,25 ℃避光培養,觀察菌株的生長特性及菌絲間的拮抗情況[12-13]。②基于ITS序列的分子鑒定與系統發育分析:稱取約0.5 g野生子實體及菌絲體,液氮冷凍研磨,再使用真菌基因組DNA小量抽提試劑盒(上海生工生物工程有限公司)提取野生子實體和菌絲體的基因組DNA,經凝膠電泳定性檢測后,作為模板,用真菌通用引物ITS1F(CTTGGTCATTTAGAGGAAGTAA)和ITS2(GCTGCGTTCTTCATCGATGC)進行PCR擴增[14-15]。擴增體系為10×PCR Buffer 2.5 μL,dNTP(2.5 mmol/L)25 L,引物(10 μmol/L)0.5 μL,TaqDNA聚合酶(5 U/μL)0.25 μL,模板DNA 1 μL,ddH2O補足至25 μL。PCR擴增程序為94 ℃預變性5 min;94 ℃變性30 s,55 ℃退火45 s,72 ℃延伸45 s,共30個循環;最后72 ℃延伸10 min,終止溫度為4 ℃。PCR產物經電泳檢測后送至測序機構(上海生工生物工程有限公司)。將序列測定結果提交至NCBI(National Center for Biotechnology Information)數據庫,獲取GenBank登錄號,再用BLAST程序進行在線同源性比對,下載相似度較高的已發表序列,以香菇Lentinulaedodes(GenBank登錄號:U33092)的ITS序列為外類群,應用MEGA 3.1軟件[16],采用鄰接法構建系統發育樹。
1.2.3 供試菌株YL-A1菌絲生長條件優化的單因素試驗 ①溫度對菌絲生長的影響:根據前期的預試驗,初步確定供試菌株生長的最適溫度范圍在18~30 ℃之間。采用PDA培養基,在直徑為7 cm的平板上接種直徑5 mm菌絲塊,分別置于18、20、22.5、25、30 ℃下避光培養5 d,每個處理均設置3次重復。按照文獻[17]中的方法測量菌落直徑后計算菌絲生長速度,同時記錄其長勢。②pH值對菌絲生長的影響:經前期預實驗后,將菌絲塊分別接種于pH為6.5、7.0、7.5、8.0、8.5的PDA培養基上,置于25 ℃培養,接種、培養及測量方法同1.2.3①,每個處理3次重復。③碳源對菌絲生長的影響:在基礎培養基中分別添加20 g不同的碳源(葡萄糖、蔗糖、麥芽糖、可溶性淀粉和乳糖),接種、培養及測量方法同1.2.3①,每個處理均設置3個重復。④氮源對菌絲生長的影響:在基礎培養基中分別添加4 g不同氮源(蛋白胨、酵母浸粉、硝酸鉀、硝酸銨、硫酸銨),接種、培養及測量方法同1.2.3①,每個處理3次重復。⑤無機鹽對菌絲生長的影響:將基礎培養基中1 g KH2PO4,分別用MgSO4、NaCl、CaCl2、KCl替換,接種、培養及測量方法同1.2.3①,每個處理3次重復。
1.2.4 不同培養因子對供試菌株YL-A1菌絲生長影響的正交試驗 不同單因子之間的交互作用對菌絲生長會有一定的影響,故在不同溫度、碳源、氮源和無機鹽等單因素試驗結果的基礎上,進行L9(34)正交試驗,四種單因素及其水平見表1,主要指標為菌絲生長速度,最終篩選各因子之間的最優組合,每組試驗設3次重復。

表1 菌株YL-A1菌絲生長正交試驗因素與水平
1.2.5 數據統計分析 采用MEGA3.1軟件分析ITS序列片段,繪制系統發育樹;用Excel和SPSS13.0軟件進行數據分析。
2 結果與分析
2.1 伊犁野生阿魏菇菌株鑒定
2.1.1 不同菌株的形態特征及菌絲間的拮抗反應 菌株YL-A1的野生子實體生于新疆阿魏的根莖部土壤(圖1A),單生,白色、松軟,花瓣形,表面有龜裂花斑,蓋緣微向內卷曲,菌蓋腹面淡黃色,有蔥蒜樣臭味(圖1B);菌株YL-A2的野生子實體生于經砍伐后剩余的新疆阿魏腐爛根上,群生,淡黃色,近圓形,表面光滑,無龜裂花斑,內實,菌肉奶油色,有蔥蒜樣臭味(圖1C)。二者經分離純化后的菌落形態無顯著性差異,均為白色,菌落平展,邊緣較整齊(圖1D)。上述描述的子實體和菌落形態特征,與曹玉清等[18]、牟川靜等[19]和陳忠純[8]報道的阿魏菇基本一致。菌絲生長特性比較及拮抗試驗結果表明(圖1D),在相同培養條件下,菌株YL-A1不僅生長速度快,而且菌絲致密,二者之間還具有明顯的拮抗反應。因此,以YL-A1為優良菌株進行后續的培養特性優化。
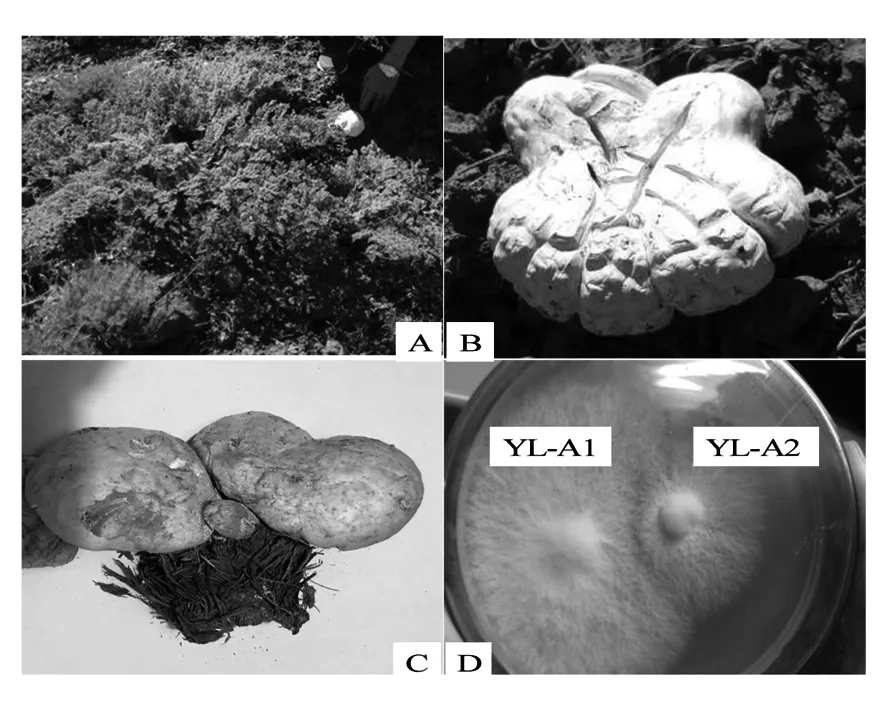
圖1 伊犁野生阿魏菇菌株YL-A1和YL-A2形態特征
2.1.2 分子鑒定與系統發育分析 rDNA ITS區段序列測定結果顯示,分別從子實體和菌絲體擴增的ITS片段是一致的,這說明經組織分離培養后得到的菌絲純培養物來源于該子實體。分離菌株YL-A1(ITS序列 GenBank登錄號:MN460317)和YL-A2(ITS序列 GenBank登錄號:MN809132)的目標擴增產物長度為分別為322 bp和326 bp。通過在線BLAST比對,發現二者序列相似度達99%,菌株YL-A1與YL-A2相比,有1個缺失位點和1個轉換位點。且二者與阿魏菇(Pleurotusferulaginis)(ITS序列 GenBank登錄號:NR_158885)[20]的遺傳關系最近,相似性均達到96%,結合生境特征及形態特征,將菌株YL-A1和YL-A2鑒定為阿魏菇(Pleurotuseryngiivar.ferulae)。系統發育分析(圖2)發現,菌株YL-A1和YL-A2單獨聚為A組,且二者有一定的遺傳距離,C[21-22]和D[22]組均來自歐洲,而E組[21,23]均來自中國。這說明即使同為阿魏菇,因長期處于不同生態環境,導致在遺傳背景上出現差異。
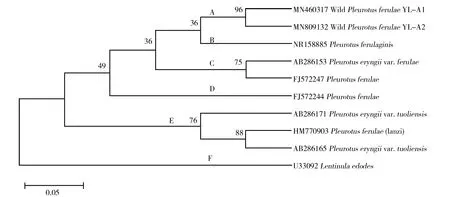
圖2 基于ITS序列構建的系統發育樹
2.2 不同培養條件對阿魏菇菌株YL-A1菌絲生長影響的單因素試驗
2.2.1 溫度 由表2可以看出,在18.0~30.0 ℃時菌絲均能生長,這與預試驗結果一致。在22.5 ℃條件下,菌絲生長最快,生長速度為(3.62±0.05)mm/d;25.0 ℃條件下,菌絲生長稍慢,但菌絲生長勢較好,表現為菌絲致密均勻。因此,綜合菌絲生長速度和長勢可知,菌株YL-A1菌絲體的最適培養溫度為22.5~25.0 ℃,屬中低溫型菌株。

表2 不同溫度對菌絲生長的影響
2.2.2 pH值 由表3可知,在pH 6.5~8.5的培養基上,菌絲均能生長,但就生長勢而言存在差異。pH 7.0~8.0時,菌絲生長致密、潔白,而在pH 6.5和8.5時,菌絲出現老化且稀疏,分析原因可能是不合適的pH改變了酶活性基團的構象,從而改變了底物的特性,對菌絲造成脅迫所致。故菌株YL-A1培養的最適pH為7.5,生長速度可達4.26 mm/d;其次為pH 8.0,生長速度為4.13 mm/d。因此,阿魏菇的菌絲體適宜在弱堿性的培養基中培養。

表3 不同pH值對菌絲生長的影響
2.2.3 碳源 由表4可知,伊犁野生阿魏菇菌絲生長過程中對碳源的利用具有選擇性,在以葡萄糖為碳源的培養基上生長最快(4.86 mm/d),菌落完整、菌絲潔白,且致密健壯;而以蔗糖和麥芽糖為碳源時,菌絲體的生長速度和長勢一般,當以乳糖為碳源時,其生長速度最慢,菌落稀疏且不完整。而以淀粉為碳源,其菌絲雖濃密而潔白,但生長速度緩慢。故最適于阿魏菇菌株YL-A1生長的碳源為葡萄糖。

表4 不同碳源對菌絲生長的影響
2.2.4 氮源 由表5可知,在添加不同氮源的培養基中菌絲體均能生長,其中在含蛋白胨和硝酸鉀的培養基中長速最快,其次是酵母浸粉,硫酸銨中最慢。從菌絲長勢上看,蛋白胨和酵母浸粉為氮源時最佳,菌絲粗壯濃密、潔白,菌落厚重、致密,表面平坦,邊緣規則,故菌株YL-A1的最佳氮源是蛋白胨,其次為酵母浸粉。
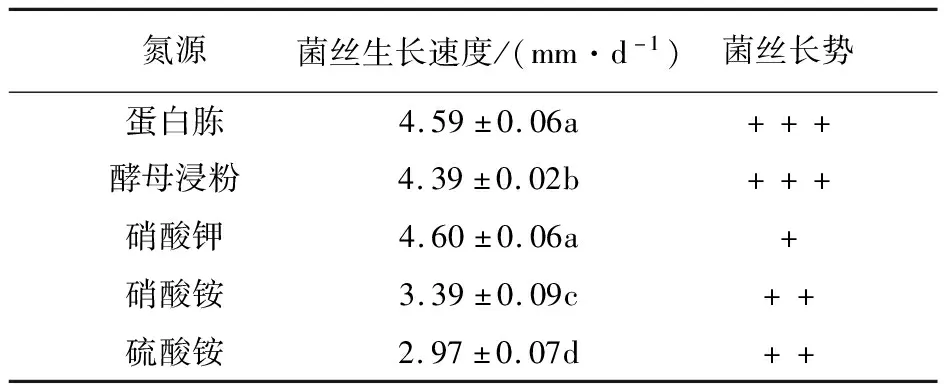
表5 不同氮源對菌絲生長的影響
2.2.5 無機鹽 培養基中不同無機鹽的添加對菌絲體生長的影響不同(表6),KH2PO4最佳,菌絲體日均長速達3.26 mm。其次為MgSO4,生長速度為3.05 mm/d,而且菌落邊緣整齊,菌絲粗壯,潔白。

表6 不同無機鹽對菌絲生長的影響
2.3 不同培養條件對阿魏菇菌株YL-A1菌絲生長影響的正交試驗
按表1設計的各個單因素及其不同水平,進行影響菌絲生長的正交試驗,結果見表7。根據R值可知,各因素對菌絲生長的影響順序依次為無機鹽(D)>溫度(A)>碳源(B)>氮源(C)。綜合各因素的K值和直觀比較,得出菌株YL-A1的最佳培養條件為A3B1C2D1。由方差分析結果可知,四個單因素影響的統計學意義并不顯著,可知各因素間組合對菌絲生長具有影響,因此考慮交互作用。最終確定菌株YL-A1的最佳培養條件:在培養基中添加20 g/L葡萄糖、4 g/L蛋白胨、1g/L KH2PO4,25 ℃恒溫避光培養,此條件下菌絲生長速度快且長勢最佳(表7)。
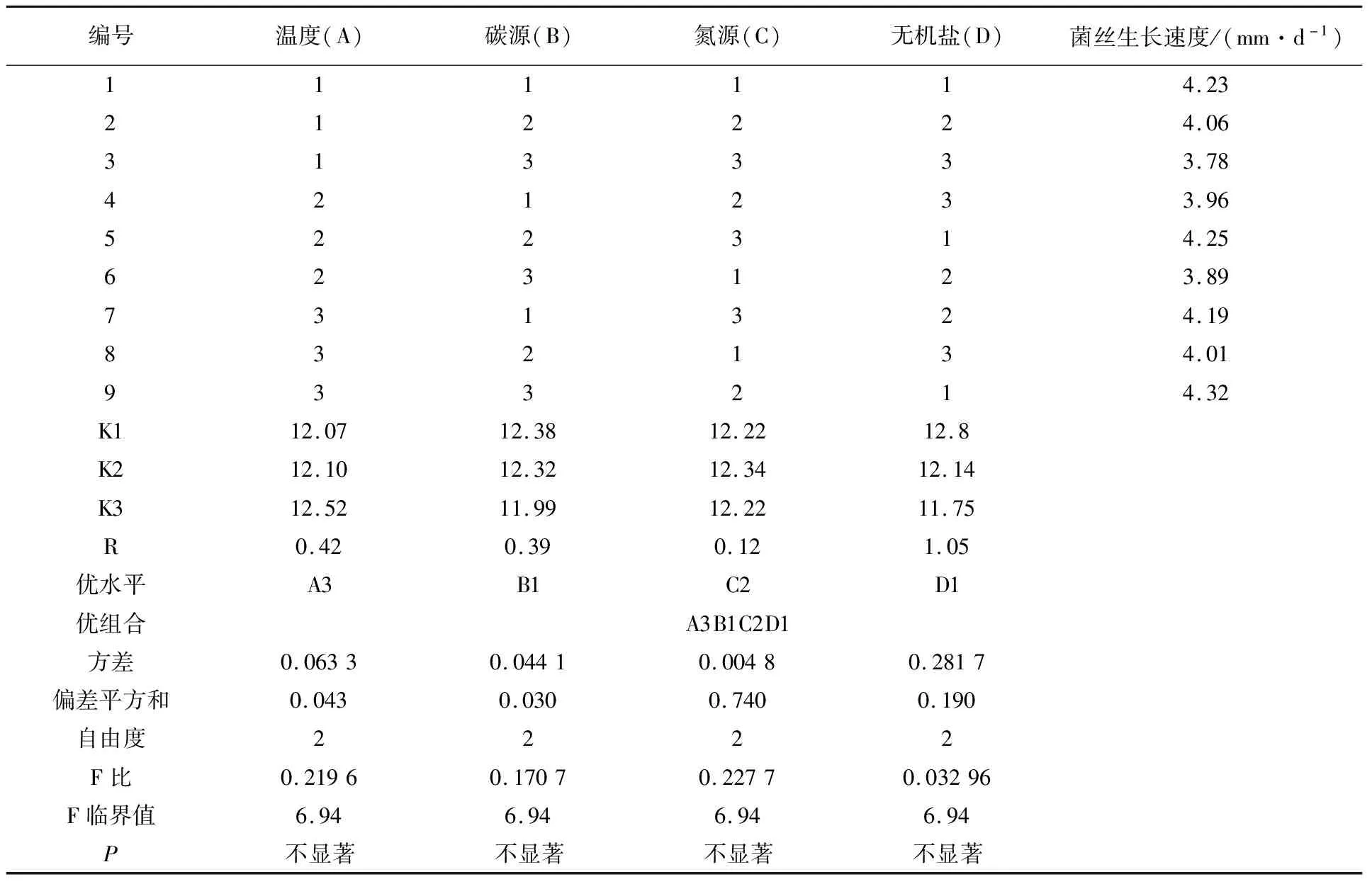
表7 正交試驗結果
3 討 論
伊犁河谷牧區具有獨特的生態環境,蘊藏了大量未知、有待開發和利用的豐富、獨特的食、藥用真菌資源。其中野生阿魏菇的人為破壞嚴重,導致自然資源匱乏[24],且還未實現菌種資源的異地保護。本研究成功實現了兩株伊犁野生阿魏菇菌種的分離純化和保藏。菌株YL-A1在生長速度和長勢上優于YL-A2,且相互拮抗。分子鑒定及系統發育分析發現,菌株YL-A1和YL-A2雖都鑒定為阿魏菇(Pleurotuseryngiivar.ferulae),但有一定的遺傳差異,說明即使來源于相同的地理位置,也可能因為生長基質、植被、海拔等環境因素的不同導致遺傳變異,ITS序列表現出差異。該試驗也表明僅通過ITS序列,在NCBI中進行BIAST比對后得出的結果可能會受具體物種ITS區的可變程度及基因庫的完善程度等因素的影響,并不可信,故不能盲目下結論[25-26],這與燕勇等[27]得出的rDNA-ITS分子鑒定方法并不能鑒定出所有真菌的結論一致,認為在野生大型真菌的分類鑒定過程中,須在傳統的形態學表型鑒定方法的基礎上,結合現代分子鑒定方法,綜合加以確定分類地位。而本研究中的兩株野生阿魏菇菌株在遺傳上的差異也成為其在生物學培養特性上的必然差異。因此,選擇生物學特性相對優良的菌株YL-A1進行后續培養特性的優化。
對于大型真菌而言,優良性能菌株的主要篩選指標是其菌絲生長速度和長勢[28]。本研究中,通過不同單因素和正交試驗,確定菌株YL-A1最適生長溫度為22.5~25.0 ℃,該結果與陳忠純等[29]的研究相似,而與劉志宏等[30]最適27 ℃的研究結果稍有偏差,分析原因可能與菌種遺傳特性及菌株生長的地理環境的不同有關。最適pH為7.5,這與劉志宏的研究結果一樣,而與曹玉清等[18]的研究結果存在一定的差異,其原因可能是在長期的進化過程中,該大型真菌為適應當地生態地理環境而進化成對酸堿度范圍具有一定耐受性的新菌種。最適碳源為葡萄糖,其次是蔗糖和麥芽糖,說明該菌能較好的吸收利用單糖和二糖,這與該大型真菌本身的酶系及其酶調節活動相關,如Kakde等[31]研究表明,葡萄糖能促進菌絲中脂肪酶活性的提高,促進菌絲生長、菌落密度的加厚和分枝的增多。菌絲生長最適氮源為蛋白胨,這一結果與宮志遠等[32]的研究結果一致。這是因為蛋白胨既提供氮素,又能提供碳素營養和生長因子;酵母浸粉屬于菌類蛋白質,更易吸收利用;此外,銨態氮的吸收利用效果優于硝態氮,分析可能是銨態氮中的氮化合價與細胞有機合成中的氮原子處于相同的氧化水平,而硝態氮中的氮需要進行同化還原才能被吸收利用;由于硫酸根離子濃度過高會對菌絲產生毒害作用[33],故該菌不能很好地利用硫酸銨。此外,在培養基中添加KH2PO4和MgSO4對菌絲生長狀態具有一定的促進作用,這與文獻中報道的磷和鎂元素是糖代謝中許多酶類發揮作用所必需的結論相一致[31,34]。本研究對伊犁野生阿魏菇進行了分離鑒定,確定了菌絲最適培養條件,這將有助于選育高產、抗逆的阿魏菇新菌株,為進一步人工馴化和開發利用提供參考。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14