新西蘭匍柄霉(Stemphylium eturmiunum)HSP42基因沉默突變株的構建及其生物學特性
2021-04-01 12:48:32萬能王群王石徐文夢趙麗麗孫文秀張修國李偉
山東農業大學學報(自然科學版) 2021年1期
萬能,王群,王石,徐文夢,趙麗麗,孫文秀,張修國*,李偉*
新西蘭匍柄霉(Stemphylium eturmiunum)HSP42基因沉默突變株的構建及其生物學特性
萬能1,王群2,王石2,徐文夢2,趙麗麗2,孫文秀1,張修國2*,李偉1*
1. 長江大學生命科學學院, 湖北 荊州 434025 2. 山東農業大學植物保護學院;山東省蔬菜病蟲生物學省級重點實驗室, 山東 泰安 271018
絲孢真菌是自然界中一類廣泛存在的真核微生物,匍柄霉屬()是絲孢真菌中的一個重要類群,而新西蘭匍柄霉()是其中的典型種,是研究絲孢真菌有性、無性進化的適用材料。熱休克蛋白(Heat shock protein,HSP),是一類進化保守的蛋白質家族,在生物體中發揮著重要的生理功能。本研究以為實驗材料,克隆鑒定了熱休克蛋白42()基因,通過根癌農桿菌介導的匍柄霉屬遺傳轉化系統,獲得了(03717)基因沉默菌株。以WT菌株為對照,對03717沉默菌株的生物學特性進行觀察發現:沉默03717后,其菌落生長減緩,菌絲細胞核分布不均勻;沉默03717后其產分生孢子時間推遲,產孢量減少,且經低溫誘導后無法產生子囊殼及子囊孢子,說明基因對的無性和有性發育具有重要影響。本研究為有性無性發育及小熱休克蛋白功能研究提供了更多依據。
新西蘭匍柄霉; 熱休克蛋白; 基因沉默
絲孢真菌是真菌中一個重要的類群,因其菌落外觀干燥,質地疏松,形態較大,繁殖能力強從而成為實驗室一類重點研究的微生物[1]。絲孢真菌主要以無性生殖方式繁殖,如暗色絲孢真菌(Dematiaceous hyphomycetes)450余屬的絕大多數種都以無性生殖方式生存繁殖,僅少數具有有性型[2]。匍柄霉屬為格孢腔菌()類,是重要的絲狀子囊真菌,而本研究所用的是其中的典型種,是能夠產生有性態的少數菌種之一,對于研究匍柄霉屬真菌的有性無性生殖進化具有重要的意義[3]。匍柄霉屬的大部分種是農業經濟作物的致病菌,能引起多種蔬菜病害,如大蒜葉枯病、洋蔥葉枯病、甜菜葉斑病、菠菜葉斑病等[4]。因此,對匍柄霉屬真菌的有性無性發育研究有重要的意義。
熱休克蛋白(Heat shock protein,HSP),也稱熱激蛋白,是生物體受到環境中物理、化學、生物等刺激時產生應激反應而合成的一類蛋白質[5]。HSP是一類龐大的蛋白質家族,在進化上較為保守,在各種生物體中都廣泛存在,并且發揮著重要的生理功能[5]。HSP種類繁多,最為常見的是以它們的分子量大小將其分為HSP60、HSP70、HSP90、HSP110及小分子熱休克蛋白sHSP幾個家族[6]。近年來,對HSP家族的研究取得了許多重要進展,尤其是對一些大分子熱休克蛋白的研究更加的深入和細致[5]。研究發現,在治療腫瘤的放射過程中,細胞內的HSP70和HSP90會高水平的表達,從而對機體的先天免疫系統產生強有力的刺激,增強免疫功能[7]。也有研究發現,HSP90在生物體的發育進化中也發揮獨特的作用[8]。對sHSP的研究也有了一定的成果,其對維持骨骼肌細胞骨架結構穩定和幫助其在損傷修復方面起著關鍵作用[9]。然而,在真菌中的sHSP研究仍需要我們進一步的探索。在本研究中,發現了基因對匍柄霉屬真菌的有性無性發育具有重要影響,也為sHSP的研究提供了更多的素材。
農桿菌介導轉化(ATMT)技術在真菌的遺傳轉化研究中得到了廣泛的應用,使得許多真菌基因功能得以闡明[10]。根癌農桿菌能夠將外源DNA質粒插入到宿主基因組中,外源基因會在宿主內得以表達,從而研究基因的功能[11]。基因沉默是研究真核生物基因功能的一種重要手段,而真菌細胞中的基因沉默是一種典型的RNAi途徑,是由雙鏈RNA引發的轉錄后基因沉默機制[12]。RNAi技術在近年來得到了不斷的研究和發展,已經越發成熟。在和粗糙脈胞菌()中使用了發夾結構RNA介導的基因沉默,其沉默效率是傳統RNAi途徑沉默效率的2.5倍左右[13]。在絲孢真菌中,利用ATMT技術將帶有發卡結構RNA的外源重組質粒導入宿主細胞中,從而引發目的基因在宿主基因組中的沉默,進一步研究基因的功能[14]。
1 材料與方法
1.1 材料來源
(EGS 29-099)菌株是與U.S. Army Natick Laboratories的Emory G. Simmons交換獲得(存放于山東農業大學植物保護學院植物病原真菌學和真菌資源研究室)。沉默載體pCIT和pCH由華中農業大學姜道宏老師實驗室惠贈,大腸桿菌DH5α購自TransGen Biotech corporation;農桿菌Agl-1和Agro菌株由吉林大學張世宏老師提供。
1.2 熱休克蛋白42基因沉默載體的構建
采用常規的CTAB法提取全基因組DNA,從其中找出基因(ORF-03717)所對應的完整核苷酸序列,并以基因組DNA為模板,利用帶有、識別位點的特異性引物PCR擴增得到03717基因的全長核苷酸序列,并將基因的啟動子區和基因的200 bp左右(共500 bp)分別正向和反向連接到沉默中間載體pCIT上,形成一個發卡結構。用Amp抗性的LB培養基進行篩選,經菌液PCR檢測后測序,便得到重組載體pCIT-03717。分析03717基因及沉默載體pCIT、pCH的多克隆酶切位點信息,選出和酶切位點,分別對pCIT-03717及pCH進行雙酶切,回收純化帶有基因的片段和線性化載體pCH,然后連接兩回收產物。通過大腸桿菌Trans5α 進行轉化實驗,并用Kana抗性的LB培養基進行篩選。通過菌液PCR檢測和測序公司測序后,獲得重組沉默載體質粒pCH-03717。特異引物如下:
03717s-1F CCATCGATGAACTGATGCCTGGAGGTGT
03717s-1R CCCAAGCTTCATGGGCTTCGTTATCAGAAC
03717s-2F CGGGATCCGAACTGATGCCTGGAGGTGT
03717s-2R TCCCCCGGGCATGGGCTTCGTTATCAGAAC
1.3 根癌農桿菌介導的S. eturmiunum遺傳轉化
參考熊晨琳[15]等建立的根癌農桿菌介導的遺傳轉化系統,首先篩選抑制WT生長的HygB最適濃度,然后制備WT菌絲懸浮液,最后進行根癌農桿菌介導的遺傳轉化。連續3次篩選后,仍可在HygB抗性的PDA平板上生長的菌落被認為是疑似沉默轉化子。
1.4 沉默轉化子的篩選和驗證
按照OMEGA公司的RNA提取試劑盒提取疑似沉默轉化子的RNA,并按照南京諾唯贊公司的反轉錄試劑盒說明書合成cDNA,以WT cDNA為對照,進行qRT-PCR分析,檢測03717基因在沉默轉化子中的沉默效率,沉默效率大于75%的即為沉默轉化子。
1.5 沉默轉化子的生物學性狀分析
沉默轉化子的生物學性狀分析主要包括:菌落形態觀察及生長速率測定,菌絲細胞核定位DAPI觀察,顯微鏡觀察分生孢子和有性態產生情況。具體方法參考熊晨琳[15]對敲除轉化子的生物學性狀分析。
2 結果與分析
2.1 HSP42基因克隆及沉默載體pCH-Gene的構建
(03717)基因序列全長為1035 bp,編碼344個氨基酸,預測分子量為37.252 kDa。克隆目的條帶在1000 bp處(圖1)。
綜合pCIT、pCH載體圖譜和03717基因序列,選擇合適的酶切位點,將03717基因先后連接到pCIT及pCH上,最終得到重組沉默載體pCH-03717。利用凍融法將重組沉默載體質轉入農桿菌感受態中,經PCR檢測后,保存陽性克隆菌液用于后續農桿菌介導的沉默轉化試驗。
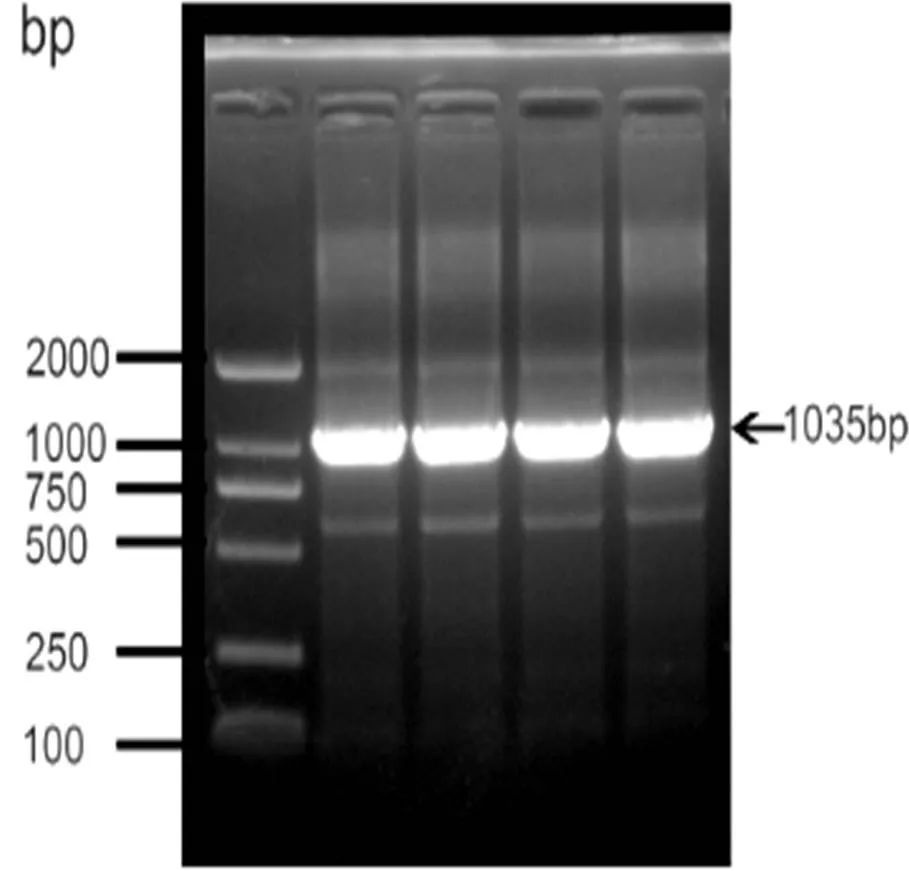
圖 1 03717基因PCR擴增結果
2.2 根癌農桿菌介導的S. eturmiunum遺傳轉化
將WT菌塊接種到含不同濃度HygB的PDA培養基上,觀察菌落的生長狀況。如圖2所示,在沒有HygB抗性的PDA平板上,菌株正常生長;隨著HygB濃度越來越高,菌絲的生長逐漸受到抑制;當HygB濃度增加到25 μg/mL時菌絲生長受到完全抑制。因此,最終確定篩選HygB的濃度為25 μg/mL。通過根癌農桿菌介導的遺傳轉化方法將重組沉默載體質粒轉移到WT菌株中后,通過PDA固體培養基(含25 μg/mL HygB抗性)篩選疑似的沉默轉化子。
2.3 沉默轉化子的獲得及驗證
連續三次篩選后仍然可以在HygB抗性的PDA平板上穩定生長的菌株被視為疑似沉默轉化子。提取疑似沉默轉化子和WT菌株的總RNA,反轉錄為cDNA,并以WT cDNA為對照,檢測幾個疑似沉默轉化子中03717基因的沉默效率,檢測結果如圖3。最終篩選出2個高效的03717沉默轉化子,沉默效率高達94%以上。
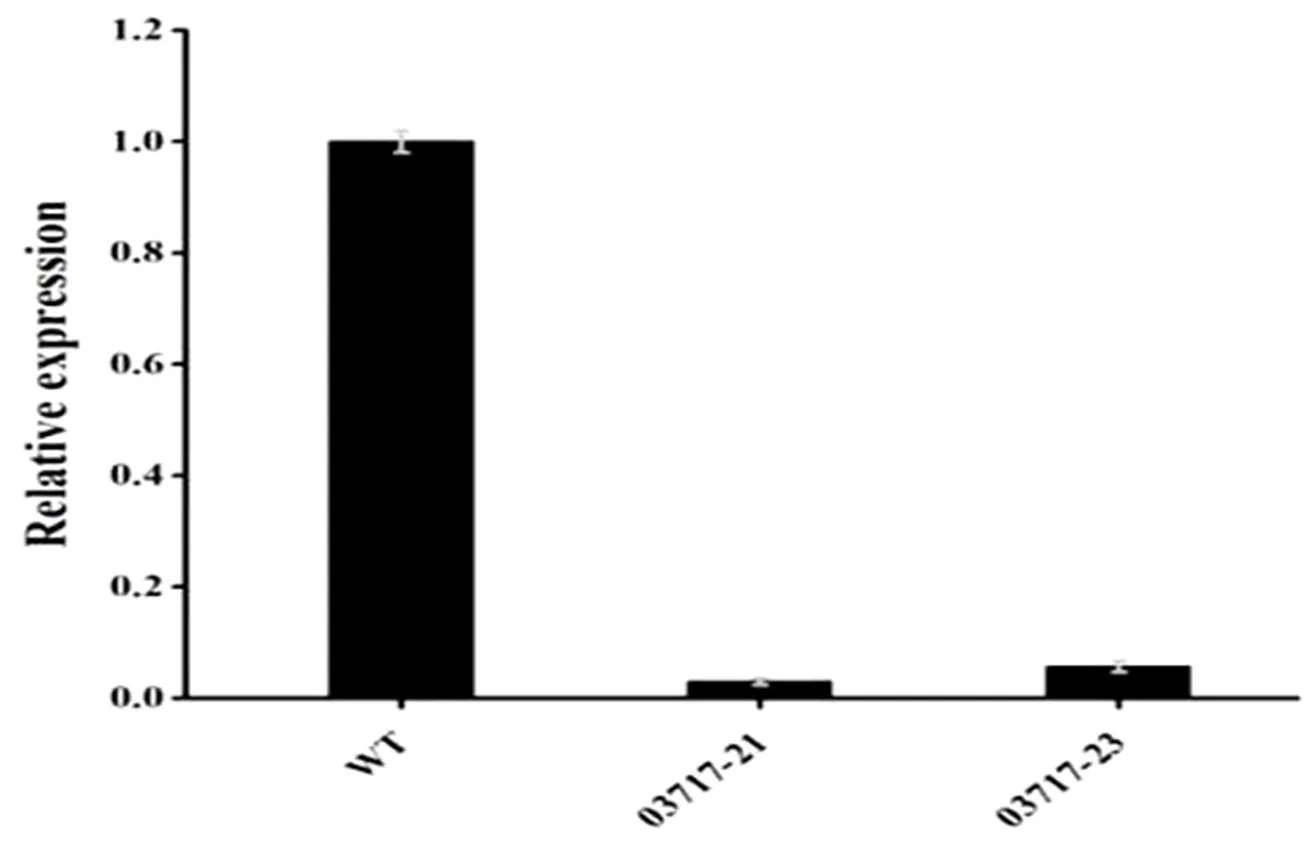
圖 3 03717在S. eturmiunum WT和沉默轉化子中的相對表達量
2.4 轉化子的生物學性狀分析
2.4.1菌落形態觀察及生長速率測定對WT菌株和03717沉默轉化子菌株同時進行培養,觀察菌落形態并測定生長速率。如圖4所示,沉默轉化子與野生型菌株相比呈現出菌落直徑略微減小、菌落顏色變淺及菌絲變緊密等特性;生長速率測定也顯示,03717沉默轉化子的生長速率略小于WT菌株的生長速率。說明03717基因對的菌落形態及菌絲生長發育具有一定調控作用。
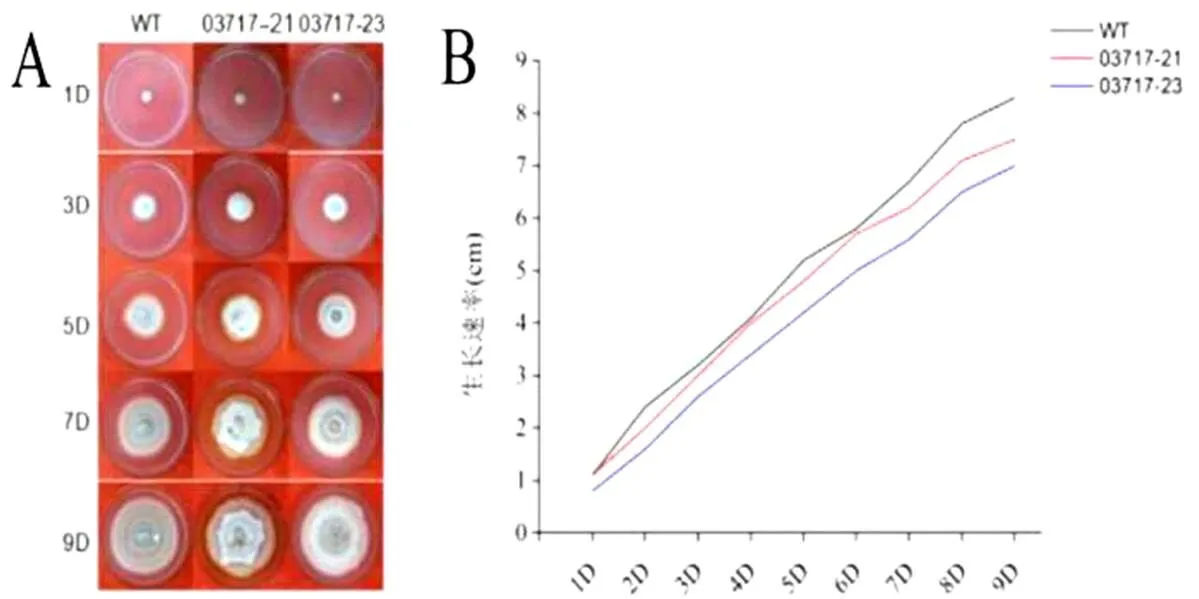
圖 4 S. eturmiunum WT及沉默轉化子菌落生長形態及生長速率
A:菌落生長形態;B: 生長速率測定
A:Colony growth morphology; B:Growth rate assay
2.4.2WT菌株和沉默轉化子的菌絲細胞核照相將插片培養4~6 d的WT菌株和沉默轉化子菌株爬片取出,通過DAPI染色制成臨時玻片,在40×熒光顯微鏡下觀察細胞核定位。如圖5所示,WT菌絲細胞核分布較為均勻,03717沉默轉化子細胞核分布無規律,且部分密集。結果證明,03717基因對的菌絲細胞核分布具有調控作用。
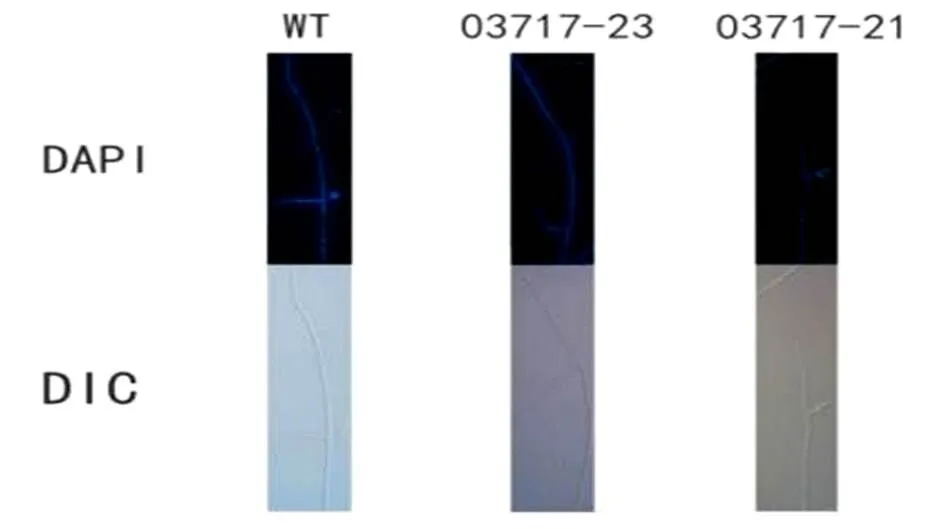
圖 5 S. eturmiunum野生型菌株和沉默轉化子核定位
2.4.3 分生孢子和有性態產生情況通過顯微觀察分生孢子和子囊殼產生情況,結果如圖6所示。沉默03717后仍然產生分生孢子,但產孢時間有所推遲,產孢數量也有所減少,結果說明03717基因對無性生殖有一定影響;WT菌株在20 d左右能夠正常產生子囊殼,4到5周就可產生子囊孢子,而03717沉默轉化子在6周后只能產生不成熟的子囊殼,經低溫誘導后無法產生子囊孢子,結果說明03717基因對有性生殖有較大影響。

圖 6 沉默轉化子分生孢子和子囊產生情況
A:分生孢子產生圖;B:子囊產生圖;C:子囊孢子產生圖
A:Conidia production ; B:Sporulation production ; C:Ascospore production
3 討論
本研究以匍柄霉屬典型種為材料,用根癌農桿菌介導的遺傳轉化方法及發夾結構RNA介導的基因沉默技術對基因進行了基因沉默,獲得了其沉默轉化子菌株,并觀察其生物學性狀。研究發現,沉默基因后,其菌落生長速率變慢,菌絲細胞核分布由較為均勻稀疏變得部分密集且無規律;沉默后其無性分生孢子產孢時間推遲且產孢量減少,沉默轉化子尚能產生不成熟的用于有性發育的子囊殼,但經低溫誘導后無法產生子囊孢子。從以上結果可以看出,基因對的無性及有性發育均有較大的影響。大分子熱休克蛋白在真菌中的研究已有了許多重大的進展,如HSP90通過介導植物免疫信號轉導途徑,參與先天免疫受體和蛋白激酶的活化與穩定,從而成為植物免疫中的重要組成成分[16]。然而,sHSP的功能在真菌中尚未得到太多的發現和驗證。本研究證明了對絲孢真菌的無性有性發育具有一定的影響,這為后人對sHSP的研究提供了更多的思路和素材,但影響無性有性發育的分子機制還未得到揭示,仍然需要我們進一步的探索。
[1] Wallroth FG. Flora Cryptogamica Germaniae, pars. Post [M]. Nuremberg: Schrag JL, 1833:923
[2] Alvaro CG, Thorner J. Heterotrimeric G protein-coupled receptor signaling in yeast mating pheromone response [J]. Journal of Biological Chemistry, 2016,291(15):7788-7795
[3] Inderbitzin P, Mehta YR, Berbee ML.species withanamorphs: a four locus phylogeny resolves new lineages yet does not distinguish among species in theclade [J]. Mycologia, 2009,101(3):329-339
[4] Falloon PG, Falloon LM, Grogan RG. Etiology and epidemiology ofleaf spot and purple spot of Asparagus in California [J]. Phytopathology, 1987,77(3):407-413
[5] 陳明帥,徐超,宋興超,等.熱休克蛋白的研究進展[J].經濟動物學報,2016,20(1):44-53
[6] Snoeckx LHEH, Comelussen RN, Van Nieuwenhoven FA,. Heat shock proteins and cardiovascular pathophysiology [J]. Physiological Reviews, 2001,81(4):1461-1497
[7] Lauber K, Brix N, Ernst A,. Targeting the heat shock response in combination with radiotherapy: Sensitizing cancer cells to irradiation-induced cell death and heating up their immunogenicity [J]. Cancer Letters, 2015,368(2):209-229
[8] Hatayama T, Takigawa T, Takeuchi S,. Characteristic expression of high molecular mass heat shock protein HSP105 during mouse embryo development [J]. Cell Structure & Function, 1997,22(5):517-525
[9] Yu JG, Fürst DO, Thornell LE. The mode of myofibril remodelling in human skeletal muscle affected by DOMS induced by eccentric contractions [J]. Histochemie, 2003,119(5):383-393
[10] 李俊香,古勤生.根癌農桿菌介導的真菌遺傳轉化研究進展[J].江蘇農學,2020,48(3):43-49
[11] Gamboa H, Judelson HS. Development of a bipartite ecdysone‐responsive gene switch for the oomycete Phytophthora infestans and its use to manipulate transcription during axenic culture and plant infection [J]. Molecular Plant Pathology, 2015,16(1):83-91
[12] Fire AZ, Xu AQ, Montgomery MK,. Potent and specific genetic interference by double-stranded RNA in[J]. Nature, 1998,391(6669):806-811
[13] Goldoni M, Azzalin G, Macino G,. Efficient gene silencing by expression of double stranded RNA in[J]. Fungal Genetics & Biology, 2004,41(11):1016-1024
[14] Meister G, Tuschl T. Mechanisms of gene silencing by double-stranded RNA [J]. Nature, 2004,431(7006):343-349
[15] 熊晨琳.asf1基因敲除突變株的構建及其生物學特性研究[D].泰安:山東農業大學,2017
[16] Song T, Ma Z, Shen D,. An oomycete CRN effector reprograms expression of plant HSP genes by targeting their promoters [J]. PLoS Pathog, 2015,11(12):e1005348
Construction and Biological Characteristics ofGene Silencing Mutant from
WAN Neng1, WANG Qun2, WANG Shi2, XU Wen-meng2, ZHAO Li-li2,SUN Wen-xiu1, ZHANG Xiu-guo2*, LI Wei1*
1.434025,2.271018,
Filamentous fungi is a kind of eukaryote which exists widely in nature, theis an important group of filamentous fungi, and theis a typical species, is a suitable material for studying sexual and asexual evolution of filamentous fungi. Heat shock protein (HSP), a class of evolutionarily conserved protein families, plays an important physiological function in organisms. The heat shock protein 42() gene was cloned and identified byas the experimental material, and the(03717) gene silencing strain was obtained bymediated genetic transformation system of. TakingWT strain as control, the biological characteristics of 03717 silent strain were observed: the colony growth slowed down and the mycelium nucleus distribution was uneven after silencing 03717; after silencing 03717, the conidia production time was delayed and the perithecia and ascospore could not be produced after low temperature induction. It shows that heat shock protein 42 gene has important effect on asexual and sexual development of. Forsexual development and small heat shock protein function, this study provides more evidence.
; heat shock protein; gene silencing
S763.15
A
1000-2324(2021)01-0001-06
10.3969/j.issn.1000-2324.2021.01.001
2020-03-18
2020-05-08
國家大宗蔬菜產業技術體系(CARS-25-03B)
萬能(1995 -),男,碩士研究生,研究方向:細胞生物學. E-mail:2514961357@qq.com
Author for correspondence. E-mail:zhangxg@sdau.edu.cn; wetli@yangtzeu.edu.cn
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小讀者(2021年2期)2021-03-29 05:03:48
科技傳播(2019年22期)2020-01-14 03:06:54
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
