放牧和氮添加對半干旱沙質草地優勢種糙隱子草及群落功能性狀的影響
2021-04-01 11:58:34孫一梅郭愛霞張森溪左小安
草地學報 2021年3期
孫一梅, 田 青*, 郭愛霞, 張森溪, 呂 朋, 左小安,4
(1. 甘肅農業大學林學院, 甘肅 蘭州 730070; 2. 中國科學院西北生態環境資源研究院, 烏拉特荒漠草原研究站, 甘肅 蘭州 730000;3. 中國科學院大學, 北京 100049; 4. 中國科學院西北生態環境資源研究院奈曼沙漠化研究站, 甘肅 蘭州 730000)
草地作為陸地生態系統的重要組成部分,占我國國土面積的41.7%[1]。過去幾十年,由于不合理的人類活動和劇烈的氣候變化導致科爾沁沙地沙漠化進程加快,生態環境不斷惡化,使其成為我國北方農牧交錯帶沙漠化最為嚴重的地區之一[2]。放牧作為草地利用方式之一,能直接或間接地影響植物群落組成、結構、功能性狀及生產力[3-4],同時氮沉降作為全球變化的主要內容之一,對半干旱沙質草地植物光合能力和養分利用效率影響較大[5]。有學者發現放牧和氮添加交互作用可能對草地生態系統存在影響,如Hodgkinson等[6]發現食草動物通常會放大資源對植物的影響;Gao等[7]發現在低耐牧性和高養分環境下,放牧與養分間表現為拮抗作用;劉思洋等[8]則發現圍封和低氮交互作用會促進植被恢復和提高植物多樣性,但高氮會降低植物多樣性和系統穩定性。因此,探究放牧和氮添加對半干旱沙質草地植物功能性狀的影響能更好地為未來草地的合理利用提供理論依據。
放牧和氮添加條件下植物通過改變各功能性狀來響應和適應環境的變化,如高度、葉面積、比葉面積、葉干物質含量及葉片氮含量等,這些性狀容易測定且與植物光合能力及資源獲取能力密切相關,因此可以很好地預測生態系統功能[9]。從不同研究水平看,種群水平的功能性狀更多地受基因和表型可塑性的影響,而群落水平上的功能性狀則更多地受環境影響[10]。另外,群落中優勢種的功能性狀對表征生境變化和群落演替方向具有一定的指示作用[11],反映出物種水平功能性狀與群落水平基本行為和功能間的密切關系[12-13]。糙隱子草(Cleistogenessquarrosa)為多年生叢生小禾草,屬C4植物,作為科爾沁沙質草地的優勢種,其水分利用效率、耐牧性、抗旱能力及光合能力等優于與其共存的C3植物[14]。研究發現,放牧可以增加糙隱子草總葉面積和葉片數,降低其單葉面積和株高[15],氮添加可以影響其高度、比葉面積及根系深度等。以往相關研究多以植物個體或群落功能性狀對放牧或氣候變化單個響應機制[16-18]為主,很少有研究基于種群和群落水平來探究功能性狀對放牧和氮添加交互作用的響應。因此,本研究以科爾沁沙地優勢種糙隱子草及其所在群落為研究對象,通過野外控制試驗,探究半干旱沙質草地優勢種糙隱子草和群落功能性狀對放牧和短期氮添加的響應,進一步揭示優勢種糙隱子草功能性狀與其所在群落性狀間的關系,從而增進氮沉降背景下草地生態系統如何響應放牧干擾的理解。
1 材料與方法
1.1 研究區概況
研究區(42°55′~42°57′ N,120°40′~120°43′ E)位于內蒙古通遼市奈曼旗境內,地處科爾沁沙地中南部,屬溫帶大陸性半干旱季風氣候,海拔約360 m,年均氣溫約6.4℃。全年≥10℃的有效積溫在3 000℃以上,無霜期150 d,年均降水量約360 mm,其中70%~80%的降水集中在6—8月,年均蒸發量約1 972.8 mm。年均風速3.2 ~ 4.1 m·s-1,春冬兩季盛行西北風,夏秋兩季為西南風[19]。該研究區流動沙丘、半固定沙丘、固定沙丘、丘間低地等各種沙丘類型交錯分布,土壤類型以風沙土、沼澤土、沙質栗鈣土為主。試驗區主要植物有:糙隱子草(Cleistogenessquarrosa)、豬毛蒿(Artemisiascoparia)、砂藍刺頭(Echinopsgmelini)、尖頭葉藜(Chenopodiumacuminatum)、白草(Pennisetumcentrasiaticum)、狗尾草(Setariaviridis)等。
1.2 試驗設計
在中國科學院西北生態環境資源研究院奈曼沙漠化研究站開展試驗。于2019年4月底選取長期封育(F,1996年封育)和放牧(G,2014年開始放牧處理)草地,放牧草地為封育草地附近無圍欄保護的公共自由放牧區。放牧強度為1頭牛·hm-2,屬于中度放牧[20]。放牧時間選擇在非生長季(9月—來年4月),生長季(5—8月)禁牧。在圍封和放牧區分別設置2個氮素水平:不施氮(N0)和添加氮(N+),小區面積為6 m×6 m,每個處理6次重復,共24個小區,試驗小區在樣地內隨機布設,小區之間設置2 m的緩沖帶。氮添加量參考國際上常用的氮素添加水平20 g·m-2·a-1[21],分兩次施加,每次10 g·m-2·a-1。施肥時間選擇5月和7月中旬,氮肥施用量根據小區面積和尿素含氮量(46%)計算。為減小氮素側向干擾,在每個試驗小區的四周挖1 m深,用包有塑料紙的金屬隔板隔開。研究選取的樣地及試驗處理位于連續的同一地段,地勢平坦,環境條件相對均質,有效地避免了空間異質性的干擾。
1.3 性狀測定
于2019年8月生物量高峰期進行調查取樣,在研究區樣地的每個小區內隨機設置3個1 m×1 m的樣方,調查樣方中植物群落物種組成、蓋度和高度。采用齊地面刈割法分物種收獲每種植物,裝入信封袋帶回實驗室,60℃烘干至恒重后稱重。采樣時為了避免邊緣效應,限制采樣點離邊界至少1 m。選擇糙隱子草(種群水平)及其群落內其它共優種(群落水平)作為研究對象,每種隨機選取5 ~ 10株植物,選擇10 ~ 20片完全伸展且無病蟲害的成熟葉片,用剪刀剪下,置于濕潤的濾紙之間,迅速放入自封袋儲藏于便攜式車載冰箱內(內部溫度< 5℃)。回實驗室后將葉片浸入水中,處于黑暗環境下冷藏(5℃)12 h。取出葉片用紙迅速吸去葉表水分,在百萬分之一的電子天平上稱得采樣葉片的總葉飽和鮮重。然后將葉片平鋪在掃描儀上掃描,再用軟件Image J計算葉面積(cm2)。掃描測定完的葉片裝入信封袋,放入60℃烘箱內烘48 h至恒重,用電子天平稱取葉片干重。將烘干的葉片磨成粉狀,過0.25 mm篩,保存備用。分析前在65℃的干燥箱中烘干1 h,放入干燥器中保存待測。利用元素分析儀(Costech ECS 4010,意大利)測定葉片氮含量。
性狀指標包括:植株高度、葉面積、比葉面積、葉干物質含量和葉氮含量。其中比葉面積和葉干物質含量的計算公式分別為:比葉面積(m2·kg-1)=葉面積/葉干重;葉干物質含量(g·g-1)=葉干重/葉飽和鮮重[22]。
重要值=(相對高度+相對蓋度+相對生物量)/3,式中:相對高度為某一物種的高度占樣方內全部物種高度之和的百分比;相對蓋度為某一物種的蓋度占樣方內全部物種蓋度之和的百分比;相對生物量為某一物種的生物量占樣方內全部物種生物量之和的百分比。
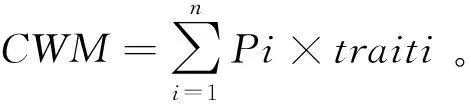
式中Pi是物種i在群落內的相對生物量,即物種i的生物量與群落中全部物種生物量和的比值;traiti是物種i的性狀值。
1.4 數據分析
雙因素方差分析比較放牧與氮添加及二者交互作用對糙隱子草和群落功能性狀的影響,單因素方差分析比較糙隱子草和群落水平功能性狀在不同處理間的差異,多重比較采用最小顯著差異法(LSD)。采用回歸分析法探究糙隱子草功能性狀與群落功能性狀之間的關系。數據統計分析與制圖分別利用SPSS 20.0與Sigmaplot 10.0軟件進行。文中數據均為平均值±標準誤。
2 結果與分析
2.1 放牧與短期氮添加處理下植物重要值變化
調查發現,試驗小區以一年生草本為主,多年生植物所占比例較少。另外,放牧與氮添加處理對植物重要值有明顯影響。圍封草地在不施氮的情況下優勢種為糙隱子草、砂藍刺頭和豬毛蒿,其重要值分別為22.44%,21.26%和20.06%;氮添加處理下優勢種為糙隱子草、白草和豬毛蒿,其重要值分別為25.37%,23.02%和19.45%。放牧草地在不施氮的情況下優勢種為砂藍刺頭、糙隱子草和白草,其重要值分別為23.19%,22.48%和14.36%;氮添加處理下優勢種為糙隱子草、豬毛蒿和白草,其重要值分別為26.25%,17.20%和15.99%(表1)。可以看出,糙隱子草在不同處理下都具有較高的重要值,其中,同一氮水平下放牧增加了糙隱子草重要值,氮添加增加了圍封和放牧草地糙隱子草的重要值。同時,不論是圍封區還是放牧區,氮添加使共優種一年生草本砂藍刺頭的重要值降低,使多年生植物白草重要值有所增加。
2.2 放牧與短期氮添加對糙隱子草功能性狀的影響
方差分析表明,放牧對糙隱子草葉面積有極顯著影響(P<0.01),氮添加對其葉干物質含量和葉氮含量有顯著影響(P<0.05),兩者交互作用對其性狀均無顯著影響(表2)。具體來看,氮添加使糙隱子草高度有不同程度的增加,放牧使其高度降低,但未達到顯著水平;FN+處理下葉面積顯著高于GN0和GN+處理,而FN+處理下的葉干物質含量顯著低于FN0處理(P<0.05);就葉氮含量而言,不論是圍封區還是放牧區,氮添加均使其顯著增加(P<0.05)(圖1)。說明糙隱子草主要通過改變葉面積來響應放牧干擾,通過改變葉干物質含量和葉氮含量來響應氮沉降。
2.3 放牧與短期氮添加對群落功能性狀的影響
方差分析表明,放牧對半干旱沙質草地植物群落高度有顯著影響(P<0.05),氮添加對葉面積、比葉面積和葉氮含量有極顯著影響(P<0.01),放牧和氮添加兩者交互處理對葉干物質含量有顯著影響(P<0.05)(表3)。具體來看,放牧(GN0和GN+)顯著降低了植物群落高度(P<0.05),而氮添加對其影響不顯著;FN+處理下葉面積顯著高于只進行圍封或放牧處理(P<0.05);比葉面積在氮添加(FN+和GN+)處理下顯著高于只進行放牧處理(P<0.05),而氮添加使得放牧區植物群落葉干物質含量顯著降低(P<0.05);就葉氮含量而言,放牧會使其降低,但未達到顯著水平,而氮添加(FN+和GN+)處理顯著增加了圍封和放牧區植物群葉氮含量(P<0.05)(圖2)。說明群落水平上植物通過改變高度來響應放牧干擾,通過葉面積、比葉面積及葉氮含量變化來響應氮沉降。
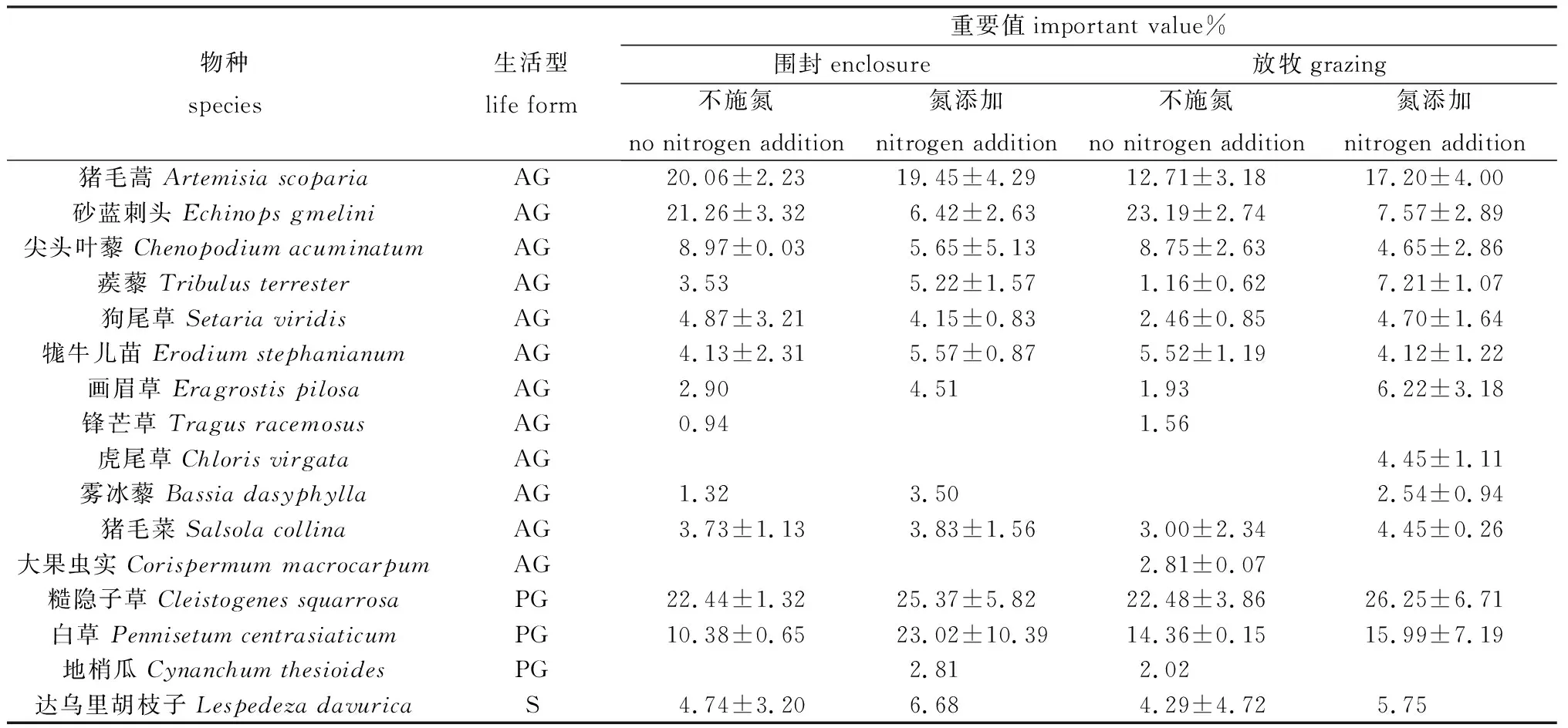
表1 不同處理下半干旱沙質草地植被重要值的變化Table 1 Changes of important values of vegetation in semi-arid sandy grassland under different treatment
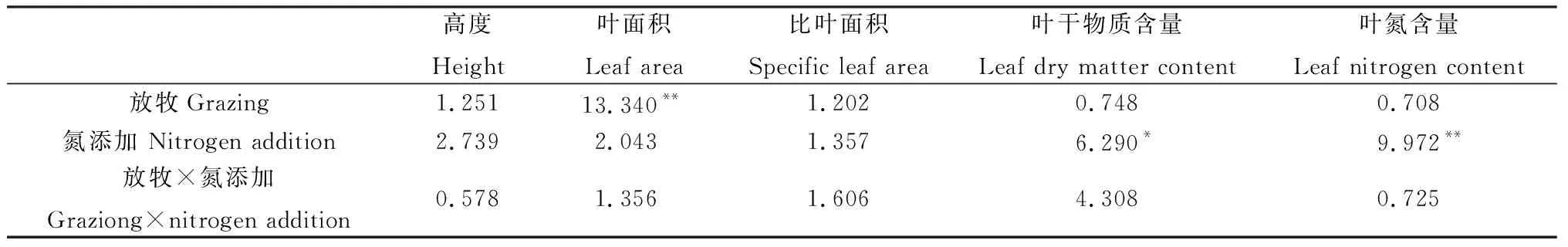
表2 放牧、氮添加及其交互作用對糙隱子草功能性狀的影響(F值)Table 2 Effects of grazing,nitrogen addition and its interaction on functional traits of Cleistogenes squarrosa (F value)
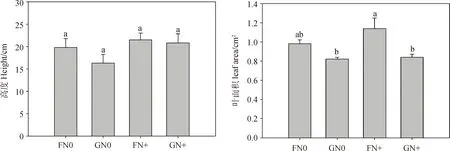
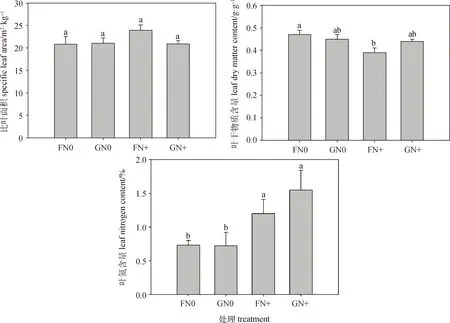
圖1 不同處理間糙隱子草功能性狀的變化Fig.1 Changes of functional traits of Cleistogenes squarrosa under different treatments注:FN0:圍封不施氮;GN0:放牧不施氮;FN+:圍封并施氮;GN+:放牧并施氮。不同小寫字母表示差異顯著(P<0.05),下同Note:F:fenced grassland with no nitrogen;G:grazing grassland with no nitrogen;FN+:fenced and nitrogen addition;GN+:grazing and nitrogen addition. Different lowercase letters indicate significant differences at the 0.05 level,the same as below
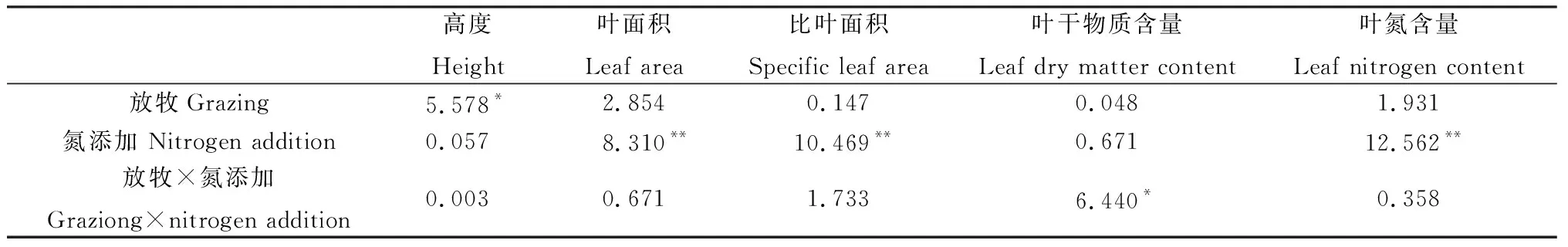
表3 放牧、氮添加及其交互作用對群落功能性狀的影響(F值)Table 3 Effects of grazing,nitrogen addition and its interaction on community functional traits (F value)
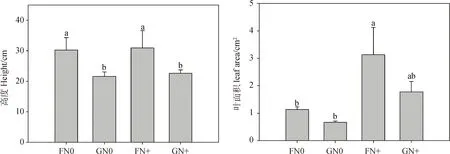
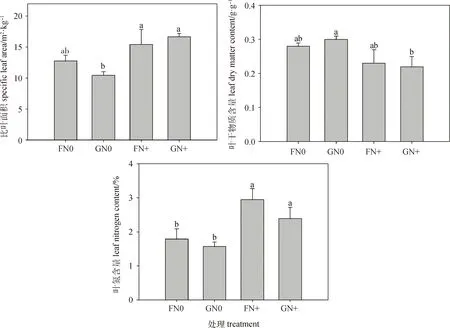
圖2 不同處理間群落功能性狀的變化Fig.2 Changes of community functional traits under different treatments
2.4 糙隱子草功能性狀與植物群落功能性狀間的關系
回歸分析表明,糙隱子草高度、葉面積、比葉面積、葉干物質含量及葉氮含量與群落相對應的功能性狀間存在不同程度的正相關關系(圖3)。其中,葉面積和葉氮含量在兩者間存在顯著正相關關系(分別為:R2=0.39,P<0.01;R2=0.24,P<0.05),而高度、比葉面積和葉干物質含量在兩者間相關性不顯著。說明糙隱子草的葉面積和葉氮含量在一定程度上能解釋群落水平葉面積和葉氮含量的變化。
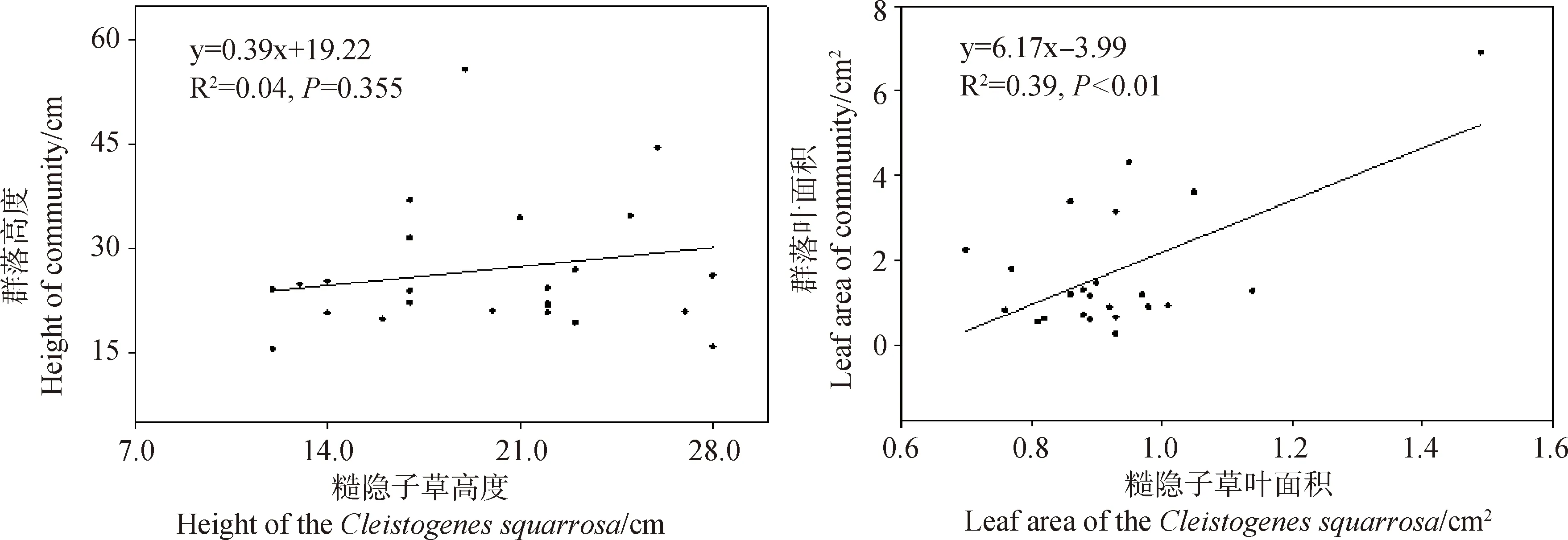
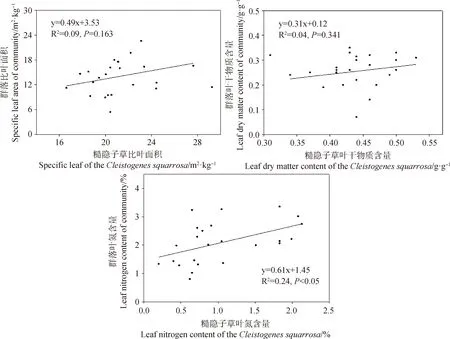
圖3 糙隱子草功能性狀與群落功能性狀間的關系Fig.3 Relationship between functional traits of Cleistogenes squarrosa and community functional traits
3 討論
3.1 放牧和短期氮添加對植物優勢度的影響
重要值反映了物種在群落中的優勢地位,重要值越大說明物種在群落中的地位越高,反之則越低[24]。本研究中放牧增加了糙隱子草的重要值,氮添加增加了圍封和放牧草地糙隱子草的重要值,說明放牧和氮添加在一定程度上可以促進糙隱子草的生長,進一步驗證了糙隱子草的超補償性生長[15]。另外,圍封樣地在不進行施氮處理下以糙隱子草、砂藍刺頭和豬毛蒿為主,添加氮素后砂藍刺頭的重要值降低,白草重要值明顯增加,說明在排除放牧干擾后白草獲取氮素的能力可能強于砂藍刺頭。因此,長期氮添加可能會影響植物群落優勢種的地位。
3.2 優勢種糙隱子草和植物群落功能性狀對放牧和短期氮添加的響應
放牧對植物生長會產生一定的影響,而植物本身也會通過調節其外部形態及生理特征產生避牧或耐牧機制,從而適應環境變化[25]。有學者認為,植株高度是植物對放牧作出響應最直觀的性狀[16]。本研究發現,放牧顯著降低了植物群落高度,符合前人提出的矮小化型變理論[26],而放牧對糙隱子草高度無明顯影響,這可能是由于糙隱子草自身高度較矮,且木質素和纖維素成分相對較高,適口性較差,不利于反芻動物的消化代謝,進而減少了家畜對其進行采食。Zheng等[27]發現,位于群落下層的糙隱子草比上層的羊草(Leymuschinensis)具有更強的避牧性和耐牧性,本研究結果與其一致。氮添加可以促進養分受限環境中植株高度的增加,以增強對光資源的競爭能力[28],但本研究中短期氮添加對群落高度無顯著影響,這可能與養分在植物不同構件中的分配差異有關[29]。增加植物高度有利于光資源的獲取,但同時伴隨著較高的成本,如投資和維持莖等支撐結構而產生成本,以及水分輸送到較高部位而產生的成本[30],因此植物可能會優先將養分分配到葉片中,同樣可以達到提高光合競爭能力的效果。
葉面積是可塑性最強、受載畜率影響縮減最為嚴重的性狀,比葉面積和葉干物質含量與植物生長與生存對策有緊密聯系,是植物比較生態學研究中的首選指標[31]。本研究結果表明,相同氮添加處理下放牧降低了糙隱子草葉面積,說明糙隱子草主要通過減小葉片大小來響應放牧干擾,而群落葉面積無顯著變化,可能是由于群落植物間存在補償效應[32]。此外,本研究發現只進行放牧處理對糙隱子草和植物群落比葉面積和葉干物質含量均無顯著影響,張晶等[17]通過研究科爾沁沙地4種草地類型中的18種植物發現,比葉面積和葉干物質含量不受放牧干擾,本研究結果與此基本一致。氮添加顯著增加了放牧區植物群落比葉面積,降低了放牧區群落葉干物質含量,說明適度放牧和短期氮添加交互作用可以通過改變與資源獲取相關的性狀來提高半干旱沙質草地植物快速獲取資源的能力,促使植物可以在不利環境中快速完成資源的累積[9,33]。
氮素是限制陸地生態系統植物生長、維持和繁殖不可或缺的養分,且與植物光合作用密切相關[34-35]。在放牧草地,氮添加可以補償因采食造成的光合面積下降,植物也可以通過調節資源分配策略將新產生的光合產物優先分配給光合組織,引起葉性狀的相應變化,進而提高植物葉性狀對家畜干擾后的恢復能力[36]。本研究中,氮添加使圍封和放牧區糙隱子草和群落葉氮含量都顯著增加,說明不論圍封草地還是放牧草地,氮添加均能緩解土壤氮素對植物生長的限制,提高植物資源競爭能力及光合利用效率,同時較高的比葉面積也可以提高植物的氮素利用效率[37]。大部分研究發現,比葉面積和葉氮含量較高以及葉干物質含量較低的植物具有較高的養分周轉率,通常表現出快速投資-收益型策略,本研究中植物性狀在群落水平表現出相似的變化,可能與本研究區以短命的一年生草本為主有關。
3.3 優勢種糙隱子草功能性狀與群落功能性狀間的相關性
群落中的優勢種可以很好地代表植物群落,也可以將觀察到的個體屬性很好地放大到群落水平甚至整個生態系統水平[36]。從糙隱子草與群落功能性狀間的相關性來看,糙隱子草葉面積和葉氮含量對群落葉面積和葉氮含量有顯著影響,高度、比葉面積和葉干物質含量對群落性狀影響較小。岳喜元等[38]發現典型草原優勢種羊草對群落水平植物的葉面積、葉干物質含量和葉氮含量有明顯影響,本研究結果與此結果具有一定的差異,可能是由于研究對象自身的形態與光合途徑差異造成的[39]。羊草為一年生C3植物,處于所在群落的上層,對光資源有更高的競爭能力,而糙隱子草為多年生C4禾草,處于群落下層,在一定程度上會受到上層植被的遮光,且糙隱子草通常采取保守型資源利用策略[27],因此與資源競爭能力密切相關的性狀,如高度、比葉面積和葉干物質含量,對群落性狀影響不顯著。此外,半干旱沙質草地植物群落以一年生植物為主,而糙隱子草為多年生植物,因此猜測半干旱沙質草地植物群落中一年生植物的功能性狀,尤其是與資源獲取、利用和植物光合競爭能力相關的性狀,對群落水平性狀變化的解釋度更高,但這需要今后進一步驗證。
4 結論
放牧和短期氮添加導致群落內植物優勢度發生變化,放牧增加了糙隱子草的重要值,氮添加使糙隱子草和白草重要值有所增加。種群水平上,放牧顯著降低了糙隱子草葉面積,氮添加增加了葉氮含量,而降低了圍封草地糙隱子草葉干物質含量。群落水平上,氮添加顯著增加了圍封區植物葉面積和葉片氮含量,而放牧樣地中,植物群落主要通過增加比葉面積和葉片氮含量、降低葉干物質含量來響應氮沉降。糙隱子草葉面積和葉氮含量能較好地解釋群落水平植物葉面積和葉氮含量的變化。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
中國科技論壇(2017年7期)2017-07-25 08:49:53
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
媽媽寶寶(2017年2期)2017-02-21 01:21:24
爆笑show(2016年7期)2017-02-09 09:36:13
國際漢語學報(2016年1期)2017-01-20 08:21:20
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
