鎘和銅對海洋浮游植物的毒性效應及其機制研究進展
2021-04-11 05:48:10段美娜
廣東海洋大學學報 2021年2期
關鍵詞:效應
段美娜,覃 亮,龍 婧,林 娜,趙 輝
(1.廣東海洋大學 化學與環境學院//2.南方海洋科學與工程 廣東省實驗室(湛江)南海資源大數據中心,廣東 湛江 524088)
重金屬污染物一般是指Hg、Cd、Pb、Cr、Cu、Zn和類金屬As等生物毒性顯著的重金屬[1]。重金屬污染物難降解且難溶于水,但對生物體的脂肪組織具有較高親和力,可以在生物體內富集并通過食物鏈逐級傳遞,進而危害人體健康[2]。近年來,隨著我國海洋經濟以及近岸城市化、工業化的發展,各種含重金屬的工業廢水、生活污水排入海洋,致使海洋環境中有毒重金屬元素含量越來越高[3]。浮游植物是海洋生態系統的初級生產者,對海洋生態系統的能量流動、物質循環和信息傳遞等方面起著十分重要的作用[4]。同時,浮游植物是海洋重金屬污染的第一層受體,也是最先受到重金屬影響的海洋生物種群之一。因此全面了解重金屬污染對海洋浮游植物的毒性效應及機制十分必要。
重金屬的生物毒性因重金屬種類[5]、元素存在形態[1]和浮游植物種類[6]而有所不同。重金屬如Cd、Hg和As等,對浮游植物而言屬于非必需元素,低濃度的此類重金屬即具有生長發育毒性效應[7-11]。Cd是具有“三致性”危害(致癌、致畸、致突變)的有毒污染物之一,已被聯合國組織確定為優先檢測的環境污染物。人類活動諸如垃圾焚燒、金屬和合金制造、電鍍、采礦、冶煉和焊接工藝等將Cd釋放到水環境中。Cd可以沿著海洋食物鏈傳遞、富集濃縮,最終進入人體,對人體內臟器官、血液循環系統和生殖系統等產生損傷。重金屬如Cu、Zn和Mn等,是浮游植物維持正常生命活動所必需的微量元素[7],它們對浮游植物具有低劑量刺激、高劑量抑制的毒物興奮效應,即當此類重金屬的含量高出某一閾值時才會對浮游植物產生毒性影響[8-9]。Cu是一種常見的重金屬,銅鋅礦的開采和冶煉、金屬加工、機械制造、鋼鐵生產等都是海洋環境中Cu污染的來源。據數據統計表明,我國近海河口環境Cu污染水平在過去幾十年中呈逐漸上升趨勢[2-3]。近海海域Cd和Cu污染問題已成為海洋生態學、環境科學和生態毒理學研究領域的熱點。
海洋環境中滲透壓、鹽類和碳源與淡水環境相比具有巨大差異,因而海洋浮游植物應對環境脅迫的機制極有可能與淡水浮游植物不同。全面了解重金屬對海洋浮游植物的毒性及其作用機制,對于預測生物群落變動趨勢和改善海洋生態環境有十分重要的意義。
本研究通過分析非必需元素Cd和必需元素Cu對海洋浮游植物的毒性效應及機制,以期為今后開展重金屬對海洋浮游植物毒害機理的深入研究提供依據。
1 Cd的毒性效應及機制
1.1 Cd的毒性效應
Cd的性質穩定且不易降解,易在生物體內蓄積,已有多篇文獻報道Cd對海洋浮游植物的生理生化過程毒性效應[12],例如生長抑制效應[11,13-29]、光合器官的損傷效應[5,11,15,24-25,27,30]、氧化應激效 應[5,13-15,17-18,22,30]、色素降解效應[5,11,13-17,21]等 (表1)。
1.2 Cd的毒性作用機制
鎘在海水中有多種存在形態,一般認為Cd2+對浮游植物具有較高的生物可利用性和毒性,因此后文主要以Cd2+為代表闡述其毒性作用機制。Cd2+對海洋浮游植物的毒性效應存在很大的種間和種內差異,但高濃度的Cd2+對絕大部分海洋浮游植物的生長發育均具抑制效應[11,13-29,34]。
1.2.1Cd對鈣信使系統的影響 Ca2+作為細胞中最重要的第二信使,參與了細胞許多生理過程的信號轉導。胞內Ca2+濃度的改變是Ca2+信使系統啟動的中心環節。Cd2+離子半徑與Ca2+相近,結構相似,Cd2+與Ca2+競爭性地利用Ca2+通道進入細胞 內[35-36],因此外界環境Cd2+濃度增加可能導致海洋浮游植物細胞對Ca2+的吸收量減少[11]。除此之外,Cd2+還可以通過自然抵抗力相關巨噬細胞蛋白、ATP結合盒轉運蛋白等通道進入浮游植物細 胞[35-36]。
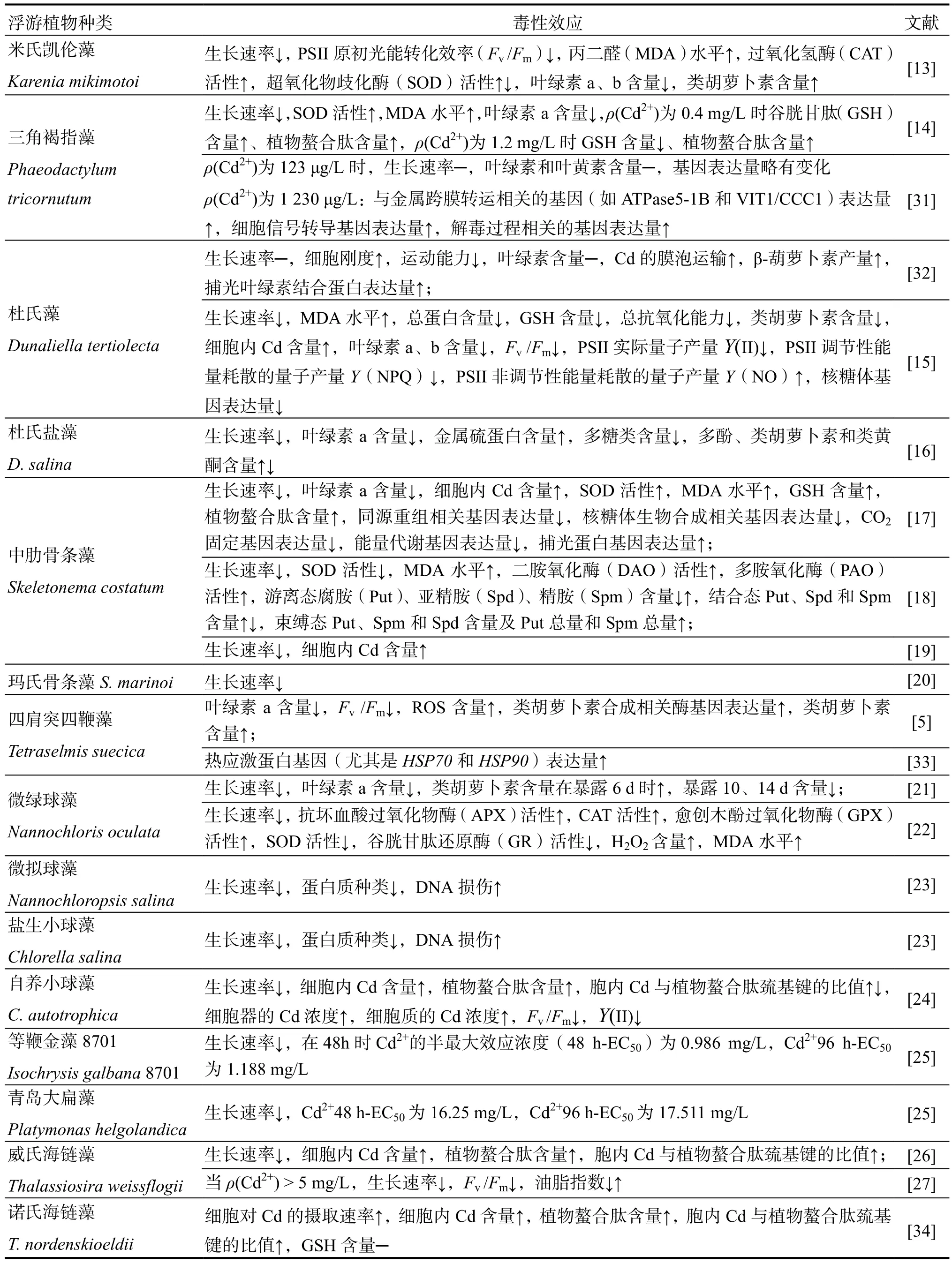
表1 重金屬Cd對海洋浮游植物的毒性效應Table 1 The toxicological effects of heavy metals (cadmium) on marine phytoplankton

續表1(Continued)
細胞內Cd2+與Ca2+之間亦存在陽離子競爭效應:1)可以替代Ca2+與下游的Ca2+信號靶蛋白結合,進而影響細胞內一系列與Ca2+相關的信號轉導過程[31];2)直接與Ca2+競爭性結合Ca2+-ATPase,導致Ca2+-ATPase活性降低,影響Ca2+的跨膜運輸;3)在線粒體、質膜、微管、肌動蛋白等的Ca2+結合部位,直接競爭或取代Ca2+,使胞質中游離的Ca2+濃度增加。另外,Cd2+極易與線粒體膜上的巰基結合,導致線粒體內膜結構改變,膜通透性增加。大約80%的胞內Ca2+貯存在線粒體、內質網等鈣庫中,一旦線粒體等膜結構功能被Cd2+破壞,將導致胞質中Ca2+濃度的增加。胞內Ca2+濃度的大幅增加,將激活某些酶如內源性核酸內切酶,導致DNA的片段化[23],從而誘導細胞凋亡。
1.2.2Cd對光合作用損傷 海洋浮游植物的光合作用對Cd2+脅迫十分敏感。Cd2+對海洋浮游植物光合作用的毒性作用機理較為復雜,可能的途徑包括5種:1)進入細胞的Cd2+可直接與葉綠素中的Mg2+發生替換,使葉綠素失去捕光功能,導致光合速率降低[5,11,13,15,24,27]。2)Cd2+具有高度的巰基親和性,可以與含巰基的生物大分子特異性結合[24,26,34],抑制海洋浮游植物的葉綠素合成酶(如原葉綠素酸酯還原酶、δ-氨基-γ-酮戊酸脫水酶)活性,同時增強葉綠素酶的活性。葉綠素酶能夠抑制原葉綠素酸酯還原酶復合物的合成。因而,Cd2+通過對葉綠素合成酶和葉綠素酶活性的影響,導致葉綠素降 解[5,11,13-17,21]。3)Cd2+可以作用于海洋浮游植物光系統電子傳遞鏈的許多位點,尤其是PSII電子供體側的放氧復合體位點,抑制光反應過程中的電子傳遞[11,30],降低光合作用效率[5,11,13,15,24,27,30]。4)Cd2+能抑制碳同化過程中的關鍵酶活性[17]。碳酸酐酶是海洋浮游植物光合作用中的關鍵酶,可以加速HCO3-和CO2之間的轉換,為光合作用中CO2固定的酶促反應提供反應底物。Cd2+可以替換碳酸酐酶活性中心的Zn2+,抑制碳酸酐酶的活性,進而干擾CO2固定過程。5)Cd2+可以破壞葉綠體結構,影響葉綠體的正常功能。對南極嗜冷單細胞藻(K.Antarctica)研究表明,Cd2+脅迫損傷了葉綠體超微結構,導致基粒片層消失、類囊體空泡、基粒堆疊混亂[11]。
1.2.3Cd誘導氧化脅迫 Cd2+進入海洋浮游植物細胞后,不僅影響光合作用,還會間接誘導ROS的生成,造成細胞的氧化損傷。ROS是生物體的正常代謝產物。海洋浮游植物體內存在抗氧化系統,包括抗氧化酶和抗氧化劑。抗氧化酶主要包括超氧化物歧化酶(SOD)、谷胱甘肽過氧化物酶(GPX)、抗壞血酸過氧化物酶(APX)、谷胱甘肽還原酶(GR)、過氧化氫酶(CAT)等,抗氧化劑主要包括還原型谷胱甘肽(GSH)、類胡蘿卜素(Car)等。未受到環境脅迫時,在抗氧化系統作用下海洋浮游植物體內ROS維持在較低水平。在受到Cd2+脅迫時,海洋浮游植物細胞內通常會積累大量的 ROS[5,22],導致氧化脅迫。雖然Cd2+是非氧化還原性金屬,無法通過Fenton和Haber-Weiss反應直接產生ROS,但Cd2+可以通過以下方式間接導致海洋浮游植物體內ROS含量的增加:1)置換蛋白質等生物大分子中的Fe或Cu等過渡性金屬,引起細胞內游離的過渡性金屬含量上升,過渡性金屬可以通過Fenton和Haber-Weiss反應加速ROS的生成,從而加劇氧化脅迫。2)Cd2+可以抑制線粒體和葉綠體的電子傳遞[30],并導致線粒體和葉綠體等細胞器的結構性損傷[11],使光呼吸功能紊亂,導致ROS的產生。3)Cd2+引起抗氧化系統功能的失調,如抑制抗氧化酶的活性[13-14,17,22]或對抗氧化劑的消 耗[14,15,30]等,導致氧化脅迫。氧化脅迫可以引起膜組分中脂質的過氧化[13-15,17,22],其直接的結果是膜不飽和脂肪酸減少、膜脂的流動性降低、膜功能異常,引起細胞透明性病變、纖維化,導致細胞裂 解[11,28]。
高濃度的ROS會導致氧化損傷,而低濃度的ROS可作為信號分子誘導浮游植物細胞的程序性死亡。H2O2呈電中性并且能夠通過細胞膜,存在時間較長,常被看作ROS中的最佳信號分子。對綠藻(Chlorella saccharophila和Micrasterias)[37-38]和甲藻(Peridinium gatunense)[39]的研究表明,在環境脅迫下H2O2作為主要的ROS信號分子,誘導了caspase-3-like酶活性上升,從而介導了細胞的類程序性死亡的發生。
2 Cu的毒性效應及機制
2.1 Cu的毒性效應
Cu是海洋浮游植物生長必需的微量元素,是呼吸作用和光合作用中多種酶的輔助因子。但過量的Cu對海洋浮游植物的毒性很大,會影響浮游植物的光合作用[5,27,40-46]、色素含量[5,40,42,45,47-48]、脂肪酸代謝[27]、碳水化合物合成[42,47,49]、ATP生成[42,49]、蛋白質合成[23,33,42,44,47-50]、基因表達[5,33,42,44,46]等多種生理生化過程。Cu對海洋浮游植物的毒性效應因受試物種不同而有所差異(表2)。
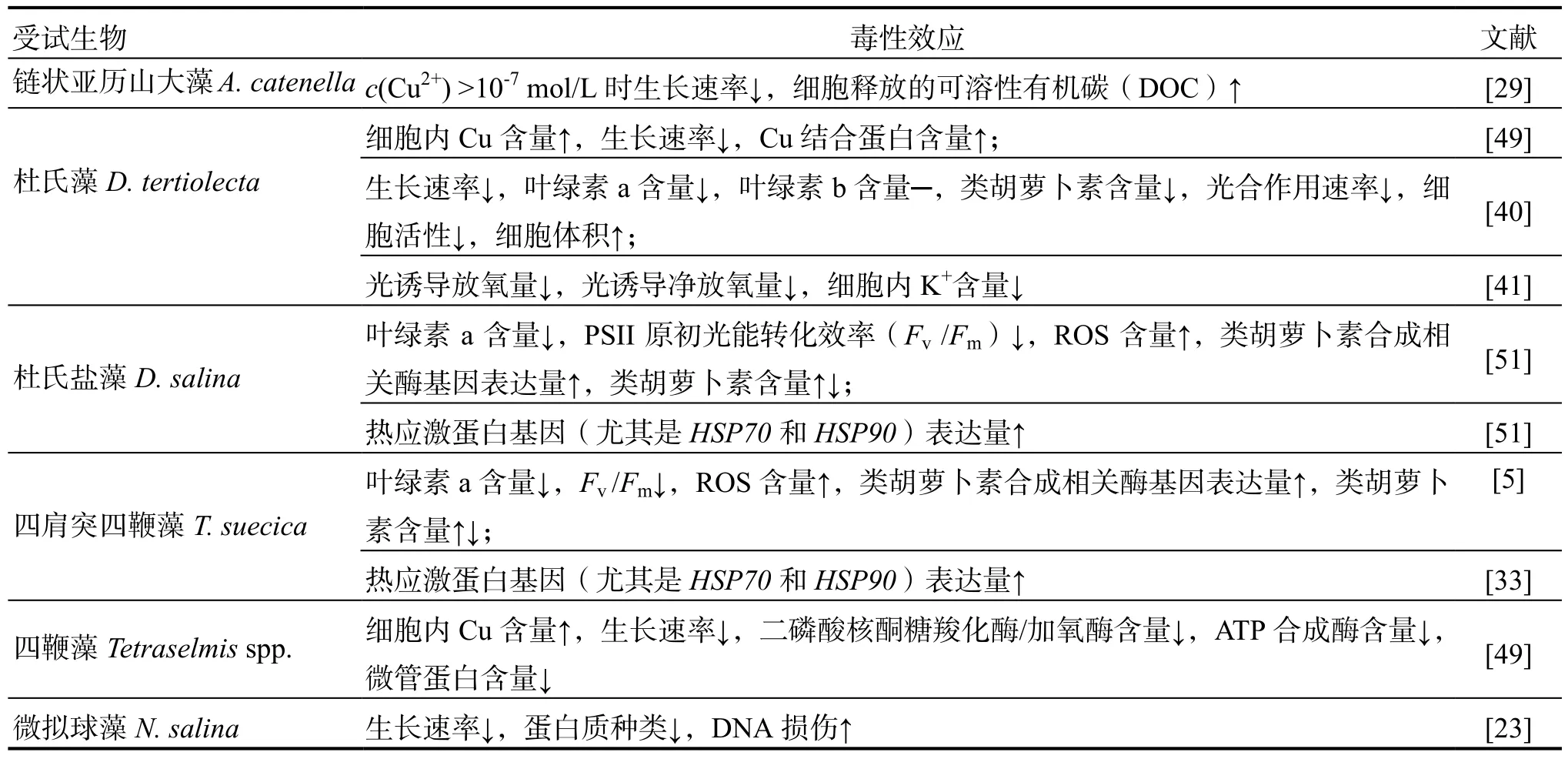
表2 重金屬Cu對海洋浮游植物的毒性效應Table 2 The toxicological effects of heavy metals (copper) on marine phytoplankton
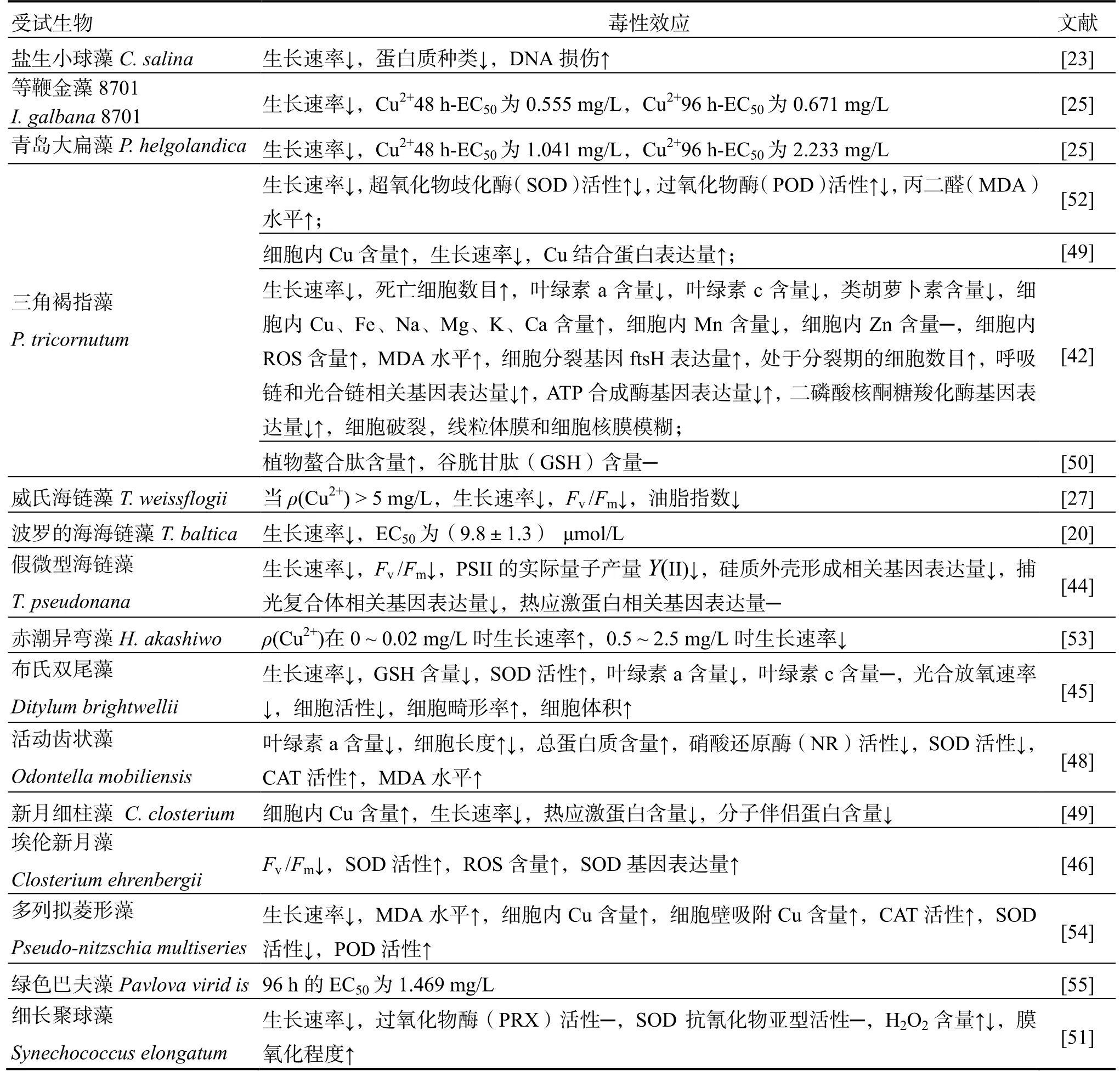
續表2(Continued)
2.2 Cu的毒性機制
Cu2+水溶性很高,在自然水體中極易遷移,一直是海洋環境研究中一個不容忽視的問題,因此后文對銅毒性機制的討論主要以Cu2+為代表。
Cu對海洋浮游植物的毒性作用機制大致可以概括為:1)Cu2+與氨基酸的羧基、羥基、氨基、巰基具有高度親和性[49],Cu2+與這些基團的結合會使蛋白質失去活性[42,48-50],干擾生理生化過程。2)Cu屬于過渡性金屬,可以與細胞內的脫氧劑反應生成Cu+,Cu+是Fenton和Haber-Weiss反應的催化劑,直接加速細胞內ROS的生成[56],誘導氧化脅迫。3)Cu2+和Cu+都可以與其他結合在特定位點的必需金屬離子發生替換,對海洋浮游植物的生長發育產生毒性效應。
2.2.1Cu與高度親和性基團結合 海洋浮游植物細胞壁和細胞膜上存在豐富的多糖類、蛋白質、脂質類物質,Cu2+具有較強的極化性,極易與這些物質的羧基、羥基、氨基、巰基等官能團結合。海洋環境中Cu2+濃度增大時,首先被吸附到浮游植物細胞壁上[54],當超出細胞壁容納能力之后,過量的Cu2+通過細胞膜進入細胞,從而產生細胞毒性[54]。海洋浮游植物細胞的膜結構由于具有豐富的不飽和脂肪酸和蛋白質,因而成為Cu2+優先攻擊的部位之一。Cu2+可與膜蛋白的官能團(如羧基、羥基、氨基等)或磷脂分子層的磷脂類物質結合,導致膜蛋白失去活性,影響膜的選擇透過性[41]和流動性,進而影響各細胞器與細胞質之間、細胞與外界之間的信號傳遞和物質運輸等生理過程,甚至會導致細胞膜破裂,細胞解體[40,42]。Cu2+還可以與細胞內含有巰基的分子如GSH[45]、植物螯合肽[50]結合。植物螯合肽主要功能是作為必需微量金屬元素的緩沖劑,螯合并運輸過量的重金屬至液泡。植物螯合肽合成的前體是GSH,其合成將消耗GSH,導致GSH含量下降。三角褐指藻(P.triconut)在受到Cu2+脅迫時,細胞內的植物螯合肽含量增加,GSH含量略有下降[50]。GSH的消耗意味著生物體的抗氧化潛力降低,這對處于過渡性重金屬(如Cu)脅迫下的海洋浮游植物影響極大。
2.2.2Cu誘導氧化脅迫 Cu是具有3種價態(0、+1、+2)的過渡金屬元素,在生物體內可以直接催化H2O2反應生成ROS。對多種海洋浮游植物的研究表明,Cu2+脅迫可導致細胞內ROS含量增加,使抗氧化防御系統失衡,引起氧化損傷[5,42,45-48,51-52,54]。ROS攻擊的主要目標之一是質膜和細胞器膜中的不飽和脂肪酸,引起膜脂過氧化[42,47-48,51-52,54],膜透性上升,導致細胞內K+大量外滲[41]。ROS還能夠氧化蛋白質側鏈中一些氨基酸的殘基,被ROS氧化的變性蛋白質最終為蛋白水解酶降解。這可能是Cu2+脅迫下,杜氏鹽藻D.salina[47]、四鞭藻Tetraselmisspp.[49]等細胞中蛋白質含量和種類減少的原因之一。此外,ROS可將DNA分子中的鳥嘌呤氧化,若此時DNA損傷修復失敗,則進一步導致堿基由G變為T,發生DNA突變[23]。Cu2+能導致大量ROS產生,海洋浮游植物細胞中的抗氧化防御系統能在一定范圍內清除ROS,保護細胞免受氧化脅迫。因而,在Cu2+脅迫下,海洋浮游植物細胞內的多種抗氧化酶(CAT、SOD等)活性及抗氧化劑(GSH等)含量發生變化[5,33,42,45-52,54]。
2.2.3Cu對光合作用的損傷 Cu脅迫對海洋浮游植物的光合作用的毒性作用機制是多方面的,可概括為:1)Cu2+可以替換葉綠素卟啉環中的Mg2+[9],生成銅代葉綠素,銅代葉綠素無法行使正常葉綠素的捕光功能,即有效葉綠素含量降低[5,40,42,45,47-48],導致海洋浮游植物的光合作用受到抑制。2)過量的Cu損傷葉綠體的膜系統[42],改變葉綠體或類囊體的結構。類囊體存在于葉綠體基質中,其膜上含有光合色素和電子傳遞鏈組分。Cu脅迫導致細胞葉綠體雙層膜結構遭破壞,基粒堆疊結構解體,因而提供給暗反應的ATP和還原氫減少,直接導致光合速率降低。3)抑制光合作用相關的酶活性或降低其含量,影響光合速率。例如二磷酸核酮糖羧化酶,它催化卡爾文循環中最初固定CO2的反應。Cu脅迫可以抑制該酶的活性,并降低海洋浮游植物細胞內該酶的含量[42,49]。4)降低光系統的電子傳遞效率[43-44]。電子傳遞鏈中傳遞電子的載體脫氫酶絕大多數包含鐵硫簇(Fe-S),由于Cu極化性比Fe強,因此相對Fe而言更加親巰基,所以Cu競爭性與硫結合形成“Cu-S簇”,抑制脫氫酶活性,造成電子傳遞不暢,導致光合作用受到抑制[27,42-44,46,49]。同時,溢出的電子還會與氧結合形成ROS,造成細胞氧化損傷。
除此之外,過量的Cu進入細胞核內會與DNA的磷酸羥基或二硫鍵結合,誘發DNA之間、蛋白質之間以及DNA和蛋白質之間的分子內和分子間交聯[23]。Cu脅迫還可能導致DNA甲基化異常,直接影響DNA的復制和轉錄水平,最終引起基因調控紊亂和蛋白質合成受抑制[5,23,42,44,47,49]。研究表明,在Cu的脅迫下四鞭藻Tetraselmissp.細胞內微管蛋白含量減少[49]。微管蛋白在浮游植物的生長和發育中起到維持細胞形態、促進細胞內運輸、參與細胞運動及細胞分裂的作用。這可能是Cu脅迫抑制海洋浮游植物細胞生長速率的原因之一。
3 展望
近年來關于重金屬污染對海洋浮游植物的毒性作用機制研究,主要以單種浮游植物為研究對象,測試指標以細胞生長速率、抗氧化酶活性、葉綠素熒光特性參數等生理生化水平指標為主。本研究表明海洋浮游植物對同一種污染物的響應具有物種差異性,而在自然海洋環境中往往是多種浮游植物和多種污染物共存。因此,在今后的研究中,筆者認為應該重點考察重金屬元素對多種浮游植物共存時的毒性機理研究,這樣結合實際情況才能為保護生態環境提供更大的幫助;加強重金屬元素在海洋浮游植物亞細胞水平上的遷移、轉運、積累的研究,基于高通量手段檢測生物體在重金屬脅迫下基因、蛋白質和代謝物等的全部信息變化,以便更全面系統地詮釋重金屬對海洋浮游植物的毒害機理。
猜你喜歡
核科學與工程(2021年4期)2022-01-12 06:30:26
今日農業(2020年19期)2020-12-14 14:16:52
小學生必讀(中年級版)(2020年9期)2020-12-04 02:07:22
科學大眾(2020年17期)2020-10-27 02:49:10
紅土地(2018年11期)2018-12-19 05:10:56
意林·全彩Color(2018年9期)2018-11-13 22:49:38
中學物理·高中(2016年12期)2017-04-22 11:53:03
中國衛生(2016年4期)2016-11-12 13:24:14
中國衛生(2014年4期)2014-12-06 05:57:14
小櫻桃·童年閱讀(2014年11期)2014-12-01 22:21:30
