甲型流感病毒與NLRP3炎性小體激活的研究進展
2021-04-12 09:26:28潘文亮江寧王松邱世昕
畜牧與獸醫 2021年4期
關鍵詞:小鼠
潘文亮,江寧,王松,邱世昕
(福建農林大學動物科學學院(蜂學學院)/閩臺動物病原生物學福建省高校重點實驗室,福建 福州 350002)
甲型流感病毒(influenza A virus,IAV)長期以來對公共衛生安全和畜牧業造成了全球性的威脅,廣泛地導致人類與畜禽的死亡,值得注意的是1918至1920年由H1N1病毒引起的“西班牙流感”大流行,據研究估計感染造成5千萬至1億人死亡;1957年的H2N2病毒造成超過200萬人死亡,1968年的H3N2病毒殺死了約100萬人[1-2]。禽類感染的甲型流感病毒H5N1和H7N9均可傳染給人類,H5N1致死率在60%,H7N9致死率在20%左右,但是其相對于H5N1而言具有更高的傳播速度[3],所以闡釋甲型流感病毒的致病機理在對其預防和治療方面尤為重要。甲型流感病毒感染所引發的嚴重疾病和造成的死亡,與宿主體內異常或者失調的細胞因子、以及炎癥反應有關[4]。細胞因子NOD樣受體蛋白3(NOD-like receptor protein 3,NLRP3)炎性小體是一種宿主天然免疫的蛋白復合物,其與病毒感染時大量促炎細胞因子分泌并造成細胞因子風暴,進而誘發過度炎癥反應有密切的關聯。本文闡述了 NLRP3 炎性小體在甲型流感病毒感染中的作用和現今研究發展,為甲型流感病毒感染引發的細胞因子風暴提供潛在的治療方向。
1 甲型流感病毒
流感病毒屬于正黏病毒科家族(Orthomyxoviridae),分為甲、乙、丙、丁或A、B、C、D四型。甲型和乙型流感病毒為具有8個基因組片段的單鏈負股RNA病毒,編碼10種必需的病毒蛋白,包含血凝素(hemagglutinin,HA)、神經氨酸酶(neuramidinase,NA)、核蛋白(nucleoprotein,NP)、非結構蛋白1(non-structural protein 1,NS1)、非結構蛋白2(nonstructural protein 2,NS2)、基質蛋白1(matrix protein 1,M1)、基質蛋白2(matrix protein 2,M2)、聚合酶堿性蛋白1(polymerase basic protein 1,PB1)、聚合酶堿性蛋白2(polymerase basic protein 2,PB2)和聚合酶酸性蛋白(polymerase acid protein,PA),以及若干個輔助蛋白轉錄本,例如與病毒毒力密切相關的非結構蛋白PB1-F2蛋白等[5-6]。另外,丙型和丁型流感病毒的遺傳物質由7個基因組片段組成,在病毒表面表達血凝素酯酶融合(hemagglutinin esterase fusion,HEF)蛋白來代替HA和NA[7]。其中,甲型流感病毒是導致人類以及畜禽流感大流行的主要病原,因此得到了廣泛的研究。
甲型流感病毒基因組長約1.35萬個堿基,由8個片段組成,能夠編碼10種必需的病毒蛋白,根據HA和NA蛋白的分子結構和遺傳特征,在人類、畜禽等宿主中發現有16種HA亞型和9種NA亞型,且近年來在蝙蝠體內檢測出H17N10和H18N11[8]。在一種甲型流感病毒與其他不同來源的流感病毒株共同感染情況下,可能會導致病毒抗原轉移,從而產生全球暴發的新型流感病毒[9]。甲型流感病毒可感染宿主肺部的免疫細胞,例如巨噬細胞和樹突狀細胞,分泌一系列促炎細胞因子和趨化因子[10],造成細胞因子風暴,其感染還能使得免疫細胞大量向肺部浸潤,從而導致急性肺組織損傷引發急性呼吸窘迫綜合征,且發病過程常常伴隨細菌感染,嚴重危害宿主的生命安全[11]。甲型流感病毒具有突變能力強,傳播速度快,對宿主的生命安全造成威脅等特點,所以對于此病毒的防治極其重要。
2 NLRP3炎性小體及其激活機制
NOD樣受體(nucleotide-binding oligomerization domain-like receptors,NLRs)是一種胞內模式識別受體(pattern recognition receptors,PRRs),它可以識別多種病原相關分子模式(pathogen-associated molecular patterns,PAMPs)和損傷相關分子模式(damage-associated molecular patterns,DAMPs)[12]。NLRP3是NOD樣受體家族中重要的一員,其炎性小體復合物由NLRP3、凋亡相關斑點樣蛋白(apoptosis-associated speck-like protein containing a CARD,ASC)和半胱天冬酶-1前體(pro-cysteinyl aspartate specific proteinase-1,pro-caspase-1)組成。NLRP3由冷誘導自身炎癥綜合征基因1(cold induced autoinflammatory syndrome 1,CIAS1)所編碼,包含一個N末端的吡啶結構域(pyrin domain,PYD)、一個中央核苷酸結合寡聚結構域(nucleoside triphosphatase domain,NACTH domain)和一個C末端富含亮氨酸的重復序列(leucine-rich repeat,LRR)[13]。NLRP3感應到危險信號時,通過LRR使得其N末端的吡啶結構域PYD與ASC的PYD相互作用并結合,之后ASC通過自身的募集結構域(caspase activation and recruitment domain,CARD)來募集同樣具有CARD的pro-caspase-1,從而聚集成NLRP3炎性小體[14]。激活的炎性小體可促使ASC將pro-caspase-1切割成具有活性的caspase-1,導致促炎細胞因子白介素1β(interleukin-1β,IL-1β)和白介素18(interleukin-18,IL-18)的成熟與分泌,并誘導細胞死亡[15-16]。甲型流感病毒感染時會激活NLRP3炎性小體,這是宿主自身的一種先天防御手段,以執行抗病毒作用。然而,在嚴重感染患者中觀察到,NLRP3炎性小體被過度激活,并造成大量的促炎細胞因子表達,進而引發細胞因子風暴造成肺組織損傷[11],這也是甲型流感病毒感染造成宿主死亡的主要原因。
在甲型流感病毒感染期間,NLRP3炎性小體由兩種危險信號來激活。第一危險信號為:Toll樣受體3(Toll-like receptor 3,TLR3)、Toll樣受體7(Toll-like receptor 7,TLR7)和維甲酸誘導基因I(retinoic acid-inducible gene-I,RIG-I),識別病毒RNA,Toll樣受體4(Toll-like receptor 4,TLR4)可識別由于甲型流感病毒感染而表達量增加的高遷移率族蛋白B1(high mobility group protein box-1,HMGB1)、然后激活核轉錄因子(nuclear factor-κB,NF-κB),促使NLRP3、pro-caspase-1、ASC蛋白、pro-IL-1β和pro-IL-18的表達上調[17-18]。第二危險信號為:病毒引發的鉀離子外流,溶酶體成熟和活性氧(reactive oxygen species,ROS)的產生,并促使NLRP3炎性小體的組裝[19]。在甲型流感病毒的研究中觀察到,其ssRNA、M2蛋白以及PB1-F2蛋白可激活NLRP3炎性小體[20-24],而NS1蛋白能夠抑制NLRP3炎性小體的第一危險信號和第二危險信號,從而抑制NLRP3炎性小體的形成[25-27]。為了更直觀地看出IAV是如何影響NLRP3炎性小體的激活的,筆者做圖以清晰明示(圖1)。由于流感病毒對NLRP3的活化發揮著雙重作用,因此闡明流感病毒感染對NLRP3活化過程的影響十分關鍵。下面將闡述流感病毒自身的ssRNA或蛋白以及病毒感染時激活的信號通路對NLRP3炎性小體活化的影響。
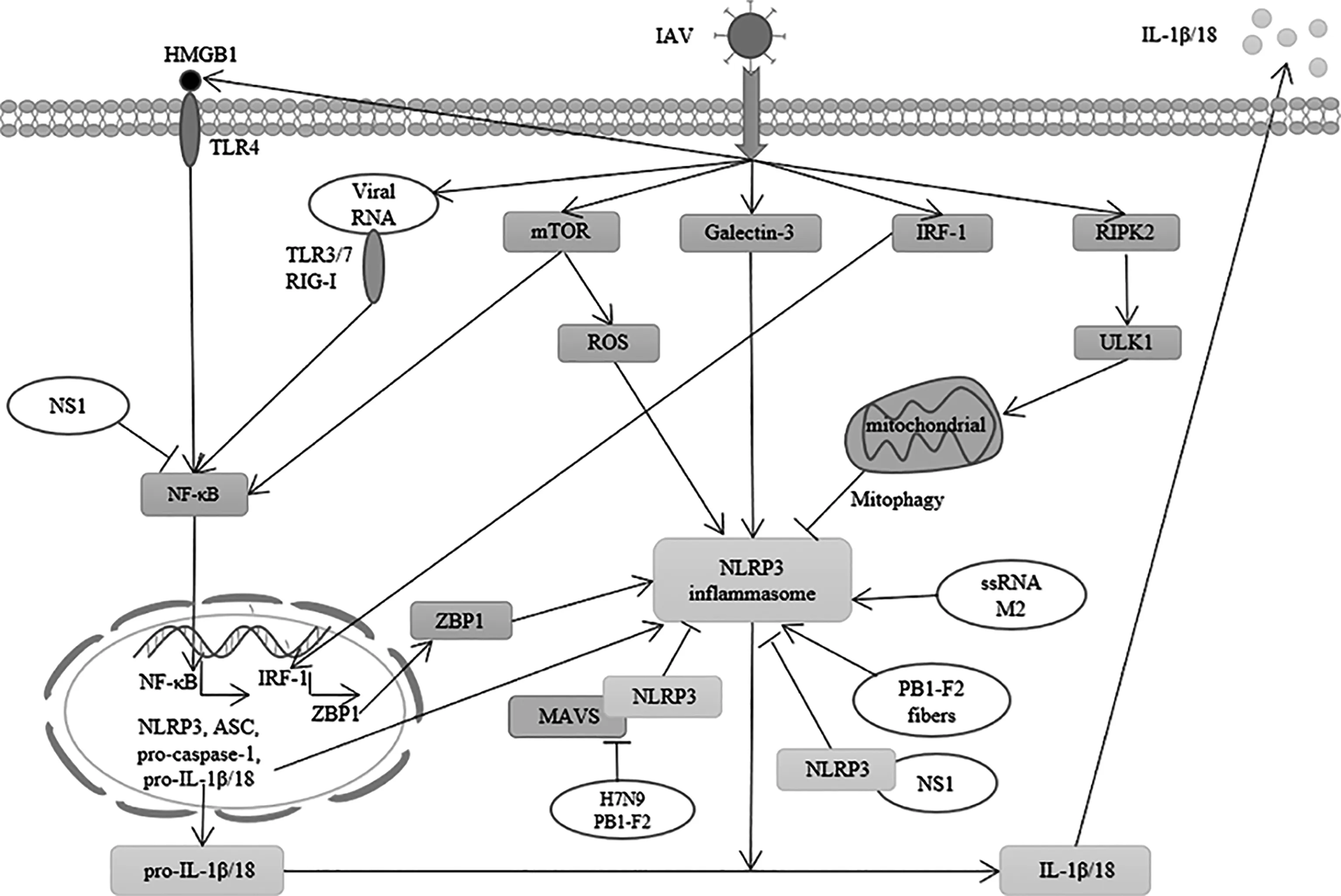
圖1 甲型流感病毒調控NLRP3炎性小體的機制
3 甲型流感病毒自身因素對NLRP3炎性小體的影響
3.1 甲型流感病毒ssRNA和M2蛋白對NLRP3炎性小體的激活作用
甲型流感病毒自身存在的一些因素,可以刺激NLRP3炎性小體的激活,從而促進IL-1β和IL-18的成熟和分泌。報道顯示,使用甲型流感病毒PR8的ssRNA處理小鼠和小鼠骨髓來源樹突狀細胞(bone marrow-derived dendritic cells,BMDCs),皆可引起caspase-1依賴性的IL-1β分泌[20]。在脂多糖(lipopolysaccharide,LPS)刺激提供第一危險信號的條件下,利用慢病毒包裝的方式,在BMDC中表達離子通道蛋白M2,觀察到IL-1β以及IL-18的分泌增加,證明甲型流感病毒M2蛋白可以通過激活第二危險信號來激活NLRP3炎性小體[21],進一步通過引入M2突變體或使用金剛烷胺(amantadine)[22]處理來阻斷M2蛋白,能有效抑制IL-1β的成熟和分泌。以上說明甲型流感病毒中的ssRNA和M2蛋白是激活NLRP3炎性小體的重要因素。
3.2 甲型流感病毒PB1-F2蛋白對NLRP3炎性小體的調控作用
甲型流感病毒中的PB1-F2蛋白常以單體、低聚物或纖維的形式存在,感染細胞后主要定位在線粒體中,可以誘導宿主細胞線粒體途徑的細胞凋亡[24]。PB1-F2纖維形式的形成取決于其C端區域是否能夠表達全長,這也是高致病性甲型流感病毒的特征之一[28]。在骨髓來源巨噬細胞(bone marrow-derived macrophages,BMMs)中處理LPS,并表達甲型流感病毒PR8(A/PuertoRico/8/34(H1N1))的PB1-F2 C端序列寡肽,能夠引發IL-1β的成熟和分泌,未使用LPS刺激的BMM中則沒有出現IL-1β的分泌,進一步對caspase-1、ASC和NLRP3基因敲除的3種不同的BMM做相同處理,均沒有誘導IL-1β的分泌[23],說明在高致病性甲型流感病毒感染過程中,其纖維形式的PB1-F2蛋白,能夠提供NLRP3炎性小體第二危險信號。有意思的是,在細胞內表達高致病性H7N9的PB1-F2蛋白,可以通過抑制線粒體抗病毒信號蛋白(mitochondrial antiviral signaling protein,MAVS)與NLRP3蛋白之間的相互作用,抑制NLRP3炎性小體的形成[29]。
3.3 甲型流感病毒的NS1蛋白對NLRP3的抑制作用
非結構蛋白NS1不僅能抑制干擾素(interferons,IFNs)的抗病毒作用,還能抑制NLRP3炎性小體的活性和IL-1β的分泌。通過研究NS1蛋白及其突變體在體外表達的方式,揭示了NS1蛋白在正常人支氣管上皮細胞(normal human bronchial epithelial cells,NHBEs)和THP-1巨噬細胞中的作用。通過抑制NF-κB的激活導致第一危險信號傳導的阻斷,使NLRP3、pro-caspase-1、ASC、pro-IL-1β和pro-IL-18的mRNA表達水平降低[25-27]。此外,NS1還能通過與NLRP3蛋白相互作用,以及抑制ASC的泛素化來抑制NLRP3炎性小體的組裝[25-26]。這些結果表明,甲型流感病毒的NS1蛋白,通過參與抑制第一和第二危險信號,減少NLRP3炎性小體依賴性促炎細胞因子的分泌。
4 甲型流感病毒介導的信號通路及其分子對NLRP3炎性小體的影響
4.1 mTOR對NLRP3炎性小體的激活作用
哺乳動物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)是一種絲氨酸/蘇氨酸激酶,由mTORC1與mTORC2兩種不同的蛋白質復合物組成,在細胞存活、增殖和代謝中起關鍵作用[30]。mTOR異常激活常發生在促炎性疾病的發病機理中,包括脂多糖誘導的急性肺損傷、敗血癥、動脈粥樣硬化和神經退行性疾病,故mTOR成為這些疾病的重要治療靶點[31]。病毒中的研究發現,甲型流感病毒pdm09(pH1N1)感染BALB/c小鼠時,能夠激活mTOR使下游NF-κB活化導致NLRP3、ASC、pro-caspase-1、pro-IL-1β和pro-IL-18的表達上調,并且mTOR激活會誘發ROS的釋放,為NLRP3炎性小體提供第二危險信號,在使用抗流感病毒藥物奧司他韋(oseltamivir)和mTOR特異性抑制劑西羅莫司(sirolimus)聯合喂食且延遲喂食BALB/c小鼠后,有效抑制了病毒介導的NLRP3炎性小體的第一和第二危險信號,降低了pH1N1的復制,且明顯地減輕了肺部炎性細胞的浸潤和肺損傷的情況[32]。以上結果確定,mTOR是甲型流感病毒感染介導激活NLRP3炎性小體的一個重要蛋白,對甲型流感病毒感染引起的肺炎具有一定的治療意義。
4.2 Galectin-3對NLRP3炎性體的激活作用
半乳凝素3(Galectin-3)屬于β-半乳糖苷結合動物凝集素家族,由約135個氨基酸的保守碳水化合物結合域構成[33]。研究顯示,在H5N1感染的小鼠肺部勻漿中,Galectin-3的mRNA和蛋白水平顯著上調,使用H5N1分別感染正常的和Galectin-3敲除的BMM,對比發現:后者在病毒感染之后IL-1β的分泌量大幅減少,ASC的低聚物水平下降;在人胚腎293T細胞(human embryonic kidney 293T cells,HEK293Ts)中過表達Galectin-3,其細胞上清液中IL-1β的分泌量增加,且通過免疫熒光檢測觀察到ASC斑點增多以及ASC與NLRP3的締合增強[34]。這些結果說明,在甲型流感病毒感染期間能夠上調Galectin-3的表達,從而促進NLRP3炎性小體的組裝和IL-1β的分泌,揭示Galectin-3在甲型流感病毒感染引發炎癥過程中的作用,為炎癥的治療提供了一些理論基礎。
4.3 IRF-1對NLRP3炎性小體的激活作用
干擾素調節因子1(interferon regulatory factor-1,IRF-1)是IRF轉錄因子家族的第一個成員,為IFN和IFN刺激基因(interferon-stimulated genes,ISGs)的調節因子,在抗病毒過程中起到作用[35]。在感染甲型流感病毒PR8的IRF1-/-骨髓衍生巨噬細胞(bone marrow derived macrophages,BMDMs)中,發現由NLRP3介導的caspase-1的活化水平與野生型(wild type,WT)組別相比有了明顯地下降,并且降低IL-1β和IL-18的成熟和分泌,使用甲型流感病毒(X31和WSN毒株)感染IRF1-/-BMDM也得到了相同的結果。此外,在感染甲型流感病毒(X31和WSN毒株)的NLRC4和AIM2缺陷細胞中,caspase-1裂解均正常發生,進一步證明了IRF-1在甲型流感病毒感染過程中,具有促進NLRP3炎性體激活的特定作用[35]。IRF-1是Z-DNA結合蛋白1(Z-DNA binding protein 1,ZBP1)的轉錄調節因子,在甲型流感病毒感染期間能夠上調ZBP1蛋白的表達,且ZBP1/DAI是流感病毒固有的傳感器,能夠在甲型流感病毒感染期間激活NLRP3炎性小體[36-37]。所以,在甲型流感病毒感染期間是通過激活IRF-1/ZBP1/NLRP3信號通路來引起炎癥反應的。
4.4 RIPK2對NLRP3炎性小體的抑制作用
受體相互作用蛋白激酶2(receptor-interacting protein kinase 2,RIPK2)是細菌感染引起炎癥反應的關鍵介質,能夠調控NF-κB以及MAPK信號通路,以激活炎癥細胞并促進促炎細胞因子和趨化因子的分泌[38]。微管相關蛋白1輕鏈3-II (microtubule-associated protein 1 light chain 3-II,LC3-II)是自噬相關蛋白,通過檢查LC3的表達水平是檢測自噬活性的一種手段。研究發現,在甲型流感病毒(PR8或X31)感染的RIPK2-/-BMDM中,其LC3-II蛋白表達量以及自噬體的計數,均遠低于WT組別的BMDM,且caspase-1的激活得到了增強,IL-18的分泌量增多,RIPK2下游的Unc-51樣自噬激活激酶1(Unc-51 like autophagy activating kinase 1,ULK1)的磷酸化水平降低。使用ULK1-/-BMDM感染PR8,觀察到相對于WT組別,細胞中線粒體自噬水平下降且caspase-1的活性有明顯的上調[39]。這說明RIPK2在甲型流感病毒感染時對NLRP3炎性小體的激活起到負調控作用。
然而,調控NLRP3炎性小體激活的細胞因子還有許多,比如蛋白激酶R(protein kinase R,PKR)能夠通過激活NF-κB以促進NLRP3炎性小體的活化[40];微小RNA-233(microRNA-233,miR-233)能夠通過抑制NLRP3蛋白的表達從而抑制炎性小體產生IL-1β[41];小二聚體伴侶(small heterodimer partner,SHP)能夠與NLRP3結合并競爭性地抑制NLRP3和ASC在線粒體上的相互作用從而抑制NLRP3炎性小體的激活[42];NLRP3炎癥小體可經干細胞因子復合體亞單位F-box和富含亮氨酸的重復蛋白2(F-box and leucine-rich repeat protein 2,FBXL2),介導的泛素化作用后被蛋白酶體降解[43]等,但是這些細胞因子在甲型流感病毒感染宿主時,對NLRP3炎性小體激活的調控機制尚未清楚。
5 NLRP3炎性小體在甲型流感病毒感染期間的抗病毒能力
NLRP3炎性小體在抗甲型流感病毒免疫應答中扮演關鍵的角色[20,44-45]。研究發現,感染PR8的NLRP3-/-、caspase-1-/-和ASC-/-小鼠與同樣感染PR8的WT小鼠相比,支氣管肺泡灌洗液(bronchoalveolar lavage fluid,BALF)中IL-6、腫瘤壞死因子- β(tumor necrosis factor- β,TNF-β)、角質形成細胞衍生細胞因子(keratinocyte-derived cytokines,KC)和巨噬細胞炎癥蛋白2(macrophage inflamma-tory protein-2,MIP-2)等的表達量以及病毒清除率降低。在這些小鼠中,還觀察到嗜中性粒細胞和單核細胞向肺部浸潤減少、小鼠死亡率增加的現象[44]。caspase-1-/-小鼠與WT小鼠相比,在感染后顯示出更嚴重的病毒性肺炎,伴有彌漫性肺泡損傷,廣泛性上皮壞死,以及甲型流感病毒抗原在肺泡上皮中的表達增加[45]。NLRP3-/-的小鼠在感染甲型流感病毒后,有明顯的上皮壞死和氣道阻塞的現象,其中纖維蛋白和壞死的細胞碎片出現膠原蛋白的沉積[20]。這些研究表明,NLRP3炎性小體,會限制甲型流感病毒感染后的病毒復制程度,并減輕病毒性肺炎和組織損傷。
6 NLRP3炎性小體的激活對甲型流感病毒致病力的促進作用
機體在感染甲型流感病毒后能夠激活NLRP3炎性小體從而促使IL-1β和IL-18成熟和分泌,IL-1β和IL-18分別與多種細胞類型上表達的細胞表面受體IL-1R和IL-18R相結合,以激活NF-kB,轉錄出炎癥因子從而誘導炎癥反應[46],這是機體對抗病毒的一種先天性免疫反應。NLRP3炎性小體的激活能夠募集免疫細胞和促炎細胞因子以對抗病毒入侵,然而持續升高的促炎細胞因子濃度能夠造成細胞因子風暴,表現為 IL-1β、IL-18、IL-6、IL-17、TNF-α、IFN-γ、干擾素誘導蛋白10(IFN-γ-inducible protein-10,IP-10)和粒細胞巨噬細胞集落刺激因子(granulocyte macrophage colony stimulating factor,GM-CSF)等細胞因子的異常高表達[47],以加劇甲型流感病毒誘發肺部損傷的程度。高濃度IL-1β的分泌可以減少細胞上皮鈉離子通道,導致肺水腫,并且與急性呼吸窘迫綜合征的發生有關[48],這是嚴重的甲型流感病毒感染患者的臨床特征。IL-18能夠誘導T細胞和自然殺傷細胞(natural killer cells,NKs)產生IFN-γ,以及促炎細胞因子引起炎癥反應[49]。因此,肺和血清中高濃度的IL-1β和IL-18,與高致病力的甲型流感病毒感染引起強烈的炎癥反應有關。
7 治療甲型流感病毒引發NLRP3依賴性炎癥的研究
MCC950是一種針對NLRP3的特異性抑制劑,能夠阻止NLRP3炎性小體依賴性caspase-1活化引起的IL-1β和IL-18的成熟和分泌。Coates等[50]研究發現,使用MCC950處理甲型流感病毒A/WSN/33(H1N1)毒株感染的THP-1巨噬細胞,對其細胞上清液進行了ELISA分析。結果顯示,caspase-1和IL-1β活化水平受到了明顯的抑制,在感染甲型流感病毒WSN毒株后接受MCC950治療的小鼠,體重減輕的情況得到改善,存活率高于未接受治療的小鼠,且小鼠肺部caspase-1和IL-1β的活化有明顯的下調,說明MCC950可以抑制甲型流感病毒感染引起的NLRP3依賴性炎癥反應。巨噬細胞是甲型流感病毒感染期間NLRP3炎性小體激活因子的主要來源,使用MCC950處理被甲型流感病毒WSN毒株感染的肺泡巨噬細胞(alveolar macrophages,AMs)能夠抑制NLRP3炎性小體的激活,在小鼠攻毒試驗中發現,MCC950不會影響WSN感染時I型干擾素的產生和病毒的清除率[50]。因此,MCC950可做為標靶藥物治療甲型流感病毒感染引發的炎癥反應。
8 結語與展望
甲型流感病毒是一種危害人類與動物健康安全,且具有極高傳播速度的危險病毒,病毒感染宿主時會激活NLRP3炎性小體,促使IL-1β和IL-18成熟和分泌,引起炎癥反應,這是宿主的先天抗病毒手段。過度激活的NLRP3炎性小體會對肺部造成嚴重損傷,這也是甲型流感病毒感染導致宿主死亡的主要原因。研究發現,甲型流感病毒的ssRNA、M2蛋白以及纖維形式的PB1-F2蛋白均能夠促進NLRP3炎性小體的激活,NS1蛋白和H7N9的PB1-F2蛋白能夠抑制NLRP3炎性小體激活。在病毒感染時,能夠通過激活mTOR,以及上調Galectin-3、IRF-1和RIPK2的表達來調控炎癥反應。近年來發現,NLRP3特異性抑制劑MCC950,對甲型流感病毒感染造成的NLRP3依賴性炎癥,在小鼠中具有一定的治療效果。雖然甲型流感病毒感染調控NLRP3炎性小體的機制研究已經取得了較為理想的突破,但是仍然有一系列的問題需要去探索,如NLRP3炎性小體活化機制的進一步研究,在甲型流感病毒感染時,如何通過把控對NLRP3炎性小體激活的正調控和負調控因素,來治療因炎性小體過度活化而導致的肺部炎癥和病理損傷等。本文通過描述甲型流感病毒感染如何調控NLRP3炎性小體的活化機制,為治療甲型流感病毒引起的炎癥反應提供了具有潛力的策略。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
