克氏原螯蝦養殖群體的SLAF測序及遺傳多樣性分析
2021-04-15 22:05:45劉亞楠劉潔魏上李亞琳李明肖俊邱高峰陸穎
南方農業學報 2021年12期
劉亞楠 劉潔 魏上 李亞琳 李明 肖俊 邱高峰 陸穎

摘要:【目的】探究克氏原螯蝦養殖群體內和群體間的遺傳多樣性,為引種和群體間雜交以改善養殖群體種質提供參考依據。【方法】以湖北、江西、安徽、浙江和江蘇5省克氏原螯蝦主產地14個養殖群體的120個個體為研究對象,采用簡化基因組測序技術——特定區點擴增片段測序技術(SLAF-seq)進行基因組測序,獲得基因組SNP基因型數據,構建群體系統發育進化樹,并進行群體結構、主成分和遺傳多樣性分析。【結果】共鑒定出741147個單核苷酸多態性(SNP)位點,群體系統進化分析表明,14個群體的種源主要來自浙江金華和江蘇宿遷,然后再向各地引種遷徙。群體結構分析結果與系統進化分析結果相吻合。主成分分析結果揭示了安徽長豐和滁州群體、湖北荊州龍口及和平2個群體,以及浙江東陽群體等5個群體與其他群體間存在相對較遠的親緣關系,是雜交引種的潛在種源。群體遺傳多樣性分析顯示,14個群體的Ho介于0.2171~0.2801,平均值為 0.2476,He介于0.3424~0.3598,平均值為0.3534,PIC介于0.2750~0.2878,群體間相差不大,接近0.25,各群體均接近遺傳多樣性中等水平的下限。【結論】14個克氏原螯蝦養殖群體內的遺傳多樣性較低,群體間的親緣關系較接近,品種單一、長期內交跡象明顯。通過群體的基因組重測序,能以較低的成本實現對養殖群體遺傳多樣性的定期監測。
關鍵詞: 克氏原螯蝦;SLAF-seq;SNP;遺傳多樣性;群體遺傳學
中圖分類號: S932? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ?文獻標志碼: A 文章編號:2095-1191(2021)12-3265-09
Analysis of genetic diversity among the farmed Procambarus clarkii populations using the SLAF-seq technology
LIU Ya-nan1,2, LIU Jie1,2, WEI Shang1,2, LI Ya-lin1,2, LI Ming3,
XIAO Jun4, QIU Gao-feng1,2, LU Ying1,2*
(1Key Laboratory of Exploration and Utilization of Aquatic Genetic Resources, Ministry of Education,Shanghai Ocean University, Shanghai? 201306, China; 2International Research Center for Marine Bioscience,Ministry of Science and Technology, Shanghai Ocean University, Shanghai? 201306, China; 3Aquatic Products Technology Extension
Station of Jinhua City, Jinhua, Zhejiang? 321017, China; 4Guangxi Academy of Fishery Sciences/Guangxi
Key Laboratory of Aquatic Genetic Breeding and Healthy Aquaculture, Nanning? 530021, China)
Abstract:【Objective】This study was designed to investigate the intra-group and inter-group genetic diversity of farmed Procambarus clarkii populations,and to provide fundamental data bases for improving the germplasm of breeding groups through required introduction and crossbreeding. 【Method】A total of 120 individuals of 14 farmed populations collected in Hubei, Jiangxi, Anhui, Zhejiang and Jiangsu were sequenced with a Specific-Locus Amplified Fragment Sequencing (SLAF-seq) technology, one of the restriction site-associated DNA sequencing technologies. The single nucleotide polymorphisms (SNPs) were identified to conduct the analyzation of phylogeny, population structure, principal component and genetic diversity. 【Result】According to the identified 741147 SNPs in this study, the phylogenic analysis suggested that the 14 populations originated from two main origins, Jinhua and Suqian. Then they were introduced and migrated to different places. The results of the population structure analysis agreed with the phylogenetic analysis. The principal component analysis revealed that the five populations, including Changfeng (Anhui), Chuzhou (Anhui), Longkou, Jingzhou (Hubei), Heping, Jingzhou (Hubei) and Dongyang (Zhejiang) had relatively distant genetic relationships to the others, which might be the potential provenances for the future hybridization. The estimated genetic diversity of the 14 populations exhibited that Ho ranged from 0.2171 to 0.2801 with a mean of 0.2476, He from 0.3424 to 0.3598 with a mean of 0.3534 and PIC from 0.2750 to 0.2878, the difference among populations was small, especially for the PIC all close to 0.25. Each population approached the lower limit of intermediate levels of genetic diversity. 【Conclusion】Comparison of sequence variations reveals a considerably low genetic diversity within and among the 14 farmed P. clarkii populations. The kinship between groups is close, with obvious signs of single breed and long-term crossing. As a low-cost and effective approach,the SLAF-seq sequencing should be suited to regularly monitor the genetic diversity of the farmed populations.
Key words: Procambarus clarkii; SLAF-seq; SNP; genetic diversity; population genetics
Foundation item: National Key Research and Development Program of “Blue Granary Science and Technology Innovation”(2018YFD0900101);Zhejiang One Million Project of Rice-fishery Integrated Planting and Raising(zjjhdy20200701);Guangxi Natural Science Foundation(2016GXNSFFA380002)
0 引言
【研究意義】克氏原螯蝦(Procambarus clarkii)俗稱小龍蝦,屬甲殼綱(Crustacea)十足目(Decapoda)鰲蝦科(Cabaridae)(劉其根等,2008),原產于北美洲墨西哥。克氏原螯蝦具有較高的營養價值及較強的環境適應能力,不僅對生長水質要求較低,甚至可以存活于高污染性的水體中。此外,其廣譜的雜食性使之還能有效清除魚類排泄物,對水體清潔有重要意義。近年來螯蝦種群已迅速蔓延到東亞、歐洲和北非尼羅河流域(宋光同等,2018;Yi et al.,2018;徐濱等,2019)。克氏原螯蝦約在20世紀30年代末由日本傳入我國(張愛軍和沈繼紅,2005),現已在我國廣泛分布。鰲蝦被作為水產資源物種以來,其養殖業發展迅速,養殖面積也越來越大,已成為一種重要的水產養殖物種(嚴維輝等,2019)。但近年來,克氏原螯蝦的養殖面臨個體變小、疫病多發、種質退化等問題,已對其養殖產業的可持續發展造成明顯影響。因此,了解不同養殖群體的遺傳多樣性信息和親緣關系,為種間交流、品種優化提供參考依據,對克氏原螯蝦養殖業的健康、可持續發展具有重要指導意義。【前人研究進展】為明確克氏原螯蝦養殖群體的遺傳信息,國內外學者主要采用微衛星序列的測序和分析。Barbaresi等(2003,2007)對歐洲5個種群的遺傳變異分析表明引種種群來自葡萄牙、法國和意大利等地區,通過分析西歐12個克氏原螯蝦群體中5個微衛星位點的遺傳變異,發現其傳播可能涉及遠距離跳躍擴散和微觀地理范圍內的自然擴散。王長忠等(2009)運用微衛星序列分析長江下游4個種群的遺傳多樣性,結果表明隨著克氏原螯蝦群體在長江流域的自然擴散和人為遷移,其遺傳多樣性出現不同程度的降低。曹玲亮等(2010)開展了安徽水系中克氏原螯蝦種群的遺傳學研究,發現該地區種群的遺傳多樣性水平和雜合度相對較高,水系間的交流是種群擴散的重要途徑。彭剛等(2010)根據7對微衛星引物的擴增產物,比較了3個克氏原螯蝦野生和養殖群體間的遺傳多樣性,得出長江流域野生群體之間具有較高遺傳多樣性的結論。邢智珺等(2014)利用8個微衛星標記對江蘇省8個地區克氏原螯蝦的遺傳多樣性進行分析,發現群體存在中等程度的分化。黃小芳等(2020)利用8對微衛星引物對克氏原螯蝦群體基因組DNA進行PCR檢測,分析其遺傳多樣性,數據顯示各群體間存在廣泛的基因交流,僅存在低度至中度的遺傳分化。Zhong等(2020)利用線粒體DNA和微衛星技術對廣西5個克氏原螯蝦群體的遺傳多樣性進行研究,結果表明遺傳變異是由種群內因素決定的,廣西南部的群體遺傳多樣性較低,應對其種質資源進行改良。從現有研究來看,普遍存在著不足之處,如調查取樣的樣品種群數量較少,樣品地理分布代表性不足;微衛星方法研究所能獲得的有效分子標記數量太少,結果的可靠性低;很少使用第二代高通量基因組測序技術和方法等。【本研究切入點】目前,國內研究極少涉及克氏原螯蝦養殖群體的遺傳多樣性分析。此外,克氏原螯蝦基因組具有高度復雜性,有94對染色體(張莎等,2018),基因組大小近3.5 Gb,參考基因組序列尚未發表。因此,采用簡化基因組測序技術——特定區點擴增片段測序技術(Specific-locus amplified fragment sequencing,SLAF-seq)(Sun et al.,2013)對其開展遺傳多樣性分析十分必要。該技術通過在基因組酶切測序后獲得的大量序列標簽上獲得的單核苷酸多態性(SNP)位點分析,能在較低測序成本下對群體遺傳多樣性實現無參分析(不依賴參考基因組的序列分析)。由于SNP的數量大,分辯率高,能適用于遺傳差異較小的養殖群體的遺傳多樣性分析。【擬解決的關鍵問題】采用基因組測序的方法對分布于湖北、江西、安徽、浙江和江蘇5省的14個克氏原螯蝦養殖群體進行測序,獲得比以往研究更豐富的基因組多態性序列標簽,以分析養殖群體間的遺傳多樣性,為判斷和指導克氏原螯蝦養殖過程中是否需要進行及時引種或群體間雜交提供重要依據。
1 材料與方法
1. 1 樣品收集和基因組DNA提取
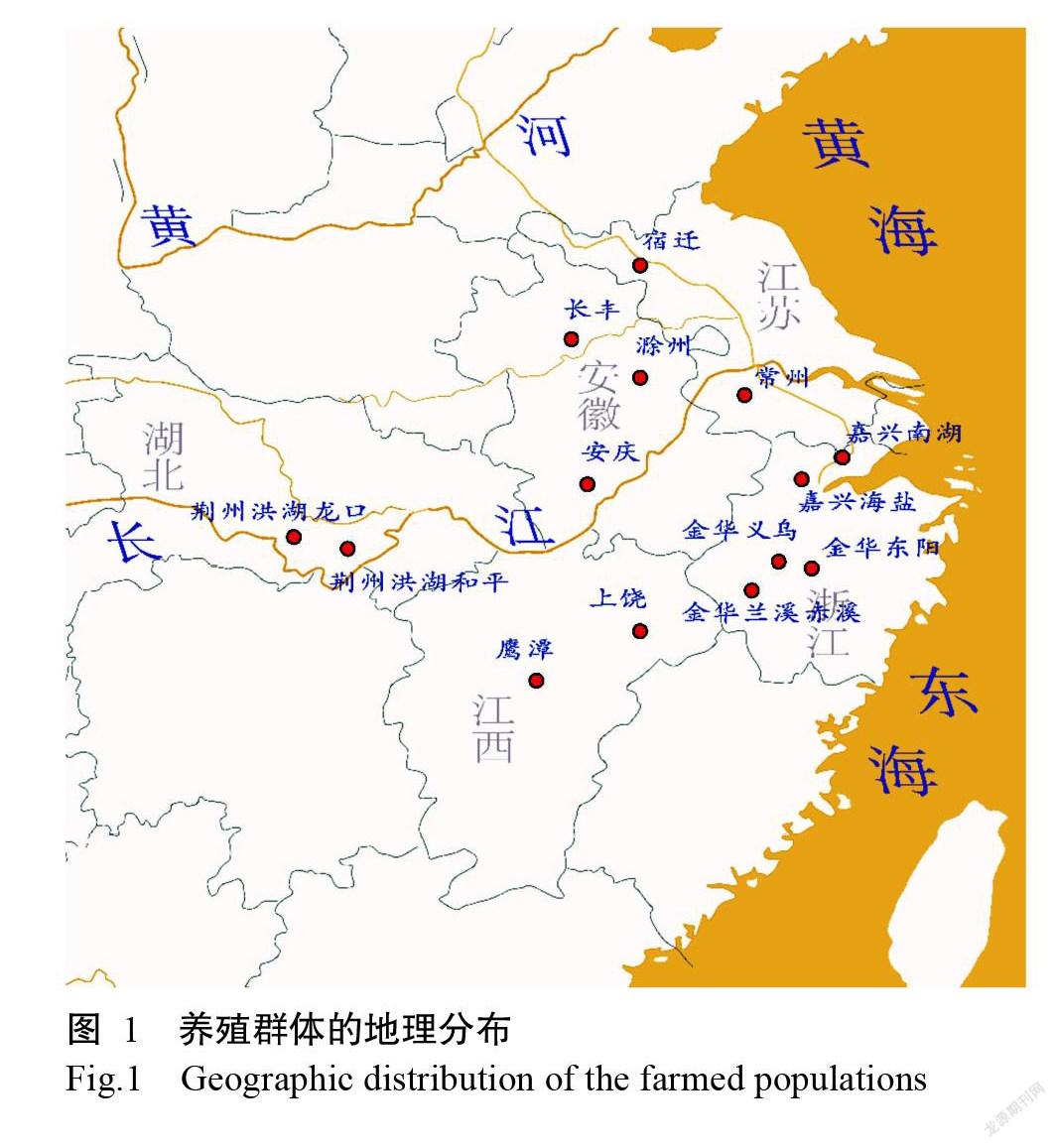
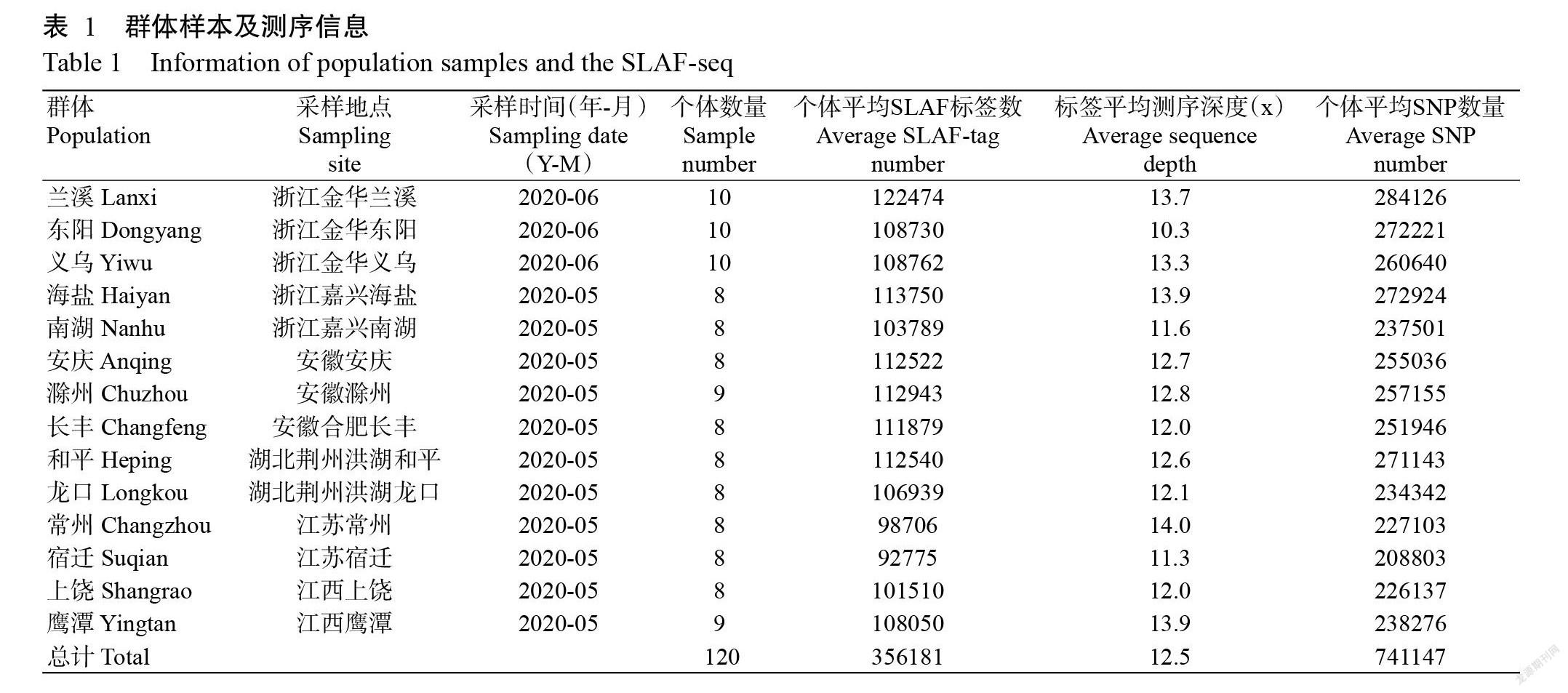
2020年5—6月采集來自浙江、安徽、湖北、江西和江蘇5省的14個養殖群體(圖1和表1),每個養殖群體選取8~10個個體的肌肉組織,共120份樣品,利用液氮速凍后,采用CTAB法提取基因組DNA(閆苗苗等,2008),用Agilent 5400檢測DNA濃度。
1. 2 簡化基因組測序和標簽SNP鑒定
預酶切實驗后,選擇限制性內切酶切HaeIII對基因組DNA進行酶切,將酶切得到的片段,即長度約400 bp的SLAF標簽,按照測序的流程構建Paire-dend測序文庫,使用HiSeq XTen測序平臺進行測序,獲得雙向長度各為150 bp的原始序列。原始序列經過去除接頭和質量評估后的干凈序列(Clean reads),按序列相似性進行聚類,來源于同一個SLAF標簽被聚類到一起。鑒定SNP時,每個SLAF標簽中測序深度最高的序列類型作為參考序列,使用BWA(Li and Durbin,2009)將Clean reads比對到對應參考序列上,使用GATK(Mckenna et al.,2010)和Samtools(Li et al.,2009)工具,默認參數下,鑒定出SNP。
1. 3 群體遺傳結構和親緣關系分析
鑒定后的SNP,使用MEGA X(Kumar et al.,2018),基于鄰接法(Neighbor-joining),采用Kimura 2-parameter模型(Davidson and Campo,2020),Bootstrap重復1000次,構建樣品的系統發育進化樹。基于SNP,使用EIGENSOFT(Price et al.,2006)進行主成分分析(PCA)。根據系統發育進化樹分析結果,計算多態信息含量(PIC),并使用Stacks v1.42計算群體觀測雜合度(Ho)和期望雜合度(He)等遺傳多樣性指數。使用ADMIXTURE(Alexander et al.,2009)分析群體結構,并進行聚類,最后根據最低交叉驗證錯誤率(CV error)值來判斷最優分群數。PIC按Botstein等(1980)的公式計算:
PIC=1-([i=1nPi2])-[i=1n-1j=i+1n2Pi2]Pj?
式中,Pi和Pj分別為第i和第j個等位基因在群體中的頻率,n為等位基因數。
2 結果與分析
2. 1 群體系統進化分析
由表1可知,用于測序的14個養殖群體共120個個體涵蓋了長江中游、下游和錢塘江等水系。對120個個體樣品進行SLAF測序后,共得到191.2 M初始Reads。序列經過聚類后,獲得超過35萬個SLAF標簽,每個標簽的平均測序深度約為12.5 x。對群體中存在多態性的SLAF標簽進行分析后,鑒定出741147個SNP位點,個體平均SNP數量在208803~284126,即使是擁有SNP數量最少的個體也攜帶16萬以上的SNP。如此數量巨大的SNP為后續準確的群體遺傳學分析打下了數據基礎。
轉換和顛換是SNP的主要類型,對這些SNP的轉換與顛換信息進行詳細統計,結果(表2)顯示,在克氏原鰲蝦各養殖群體中,轉換與顛換的比例在1.4∶1左右,高于完全隨機情形下的1∶2。同大多數后生動物相似,存在轉換——顛換的偏差(Seplyarskiy et al.,2012)。通常,在密碼子中發生的顛換更易導致氨基酸的改變,且轉換比起顛換更容易,因此這在進化上是合理的。
利用這些SNP,構建120個克氏原螯蝦個體的系統發育進化樹(圖2)。系統發育進化樹顯示樣本在遺傳上主要聚成2個簇(clade):一支由浙江金華的義烏和蘭溪群體與浙江嘉興的海鹽和南湖2個群體構成;另一支由浙江金華東陽的群體和其余4省的其他群體構成。由個體在系統發育進化樹上的分布可看出,浙江義烏、江西鷹潭、湖北洪湖和平及龍口、安徽滁州、安徽合肥長豐和江蘇常州等養殖點的個體幾乎均單獨聚集在一起,說明這些養殖點品種較單一,幾乎沒有與其他養殖點間進行過雜交。而如浙江金華蘭溪、浙江嘉興海鹽和南湖、安徽安慶、江西上饒等群體,個體的分布與其他養殖點的個體有較明顯的混雜,表明該養殖點品種與其他養殖點品種間交流較多。系統進化分析揭示某些在地理分布上有較遠距離的群體卻顯示出較近的親緣關系,如浙江金華東陽與湖北荊州的2個群體,江蘇常州與江西上饒、安徽安慶的3個群體間在進化關系上較接近。
根據所有樣品的系統進化關系,可大致預測出克氏原螯蝦養殖種群的遷徙過程(圖3)。由圖3可看出,取樣點的克氏原螯蝦最初可能來自兩個祖先群體,在這些養殖點中,目前能回溯到與這兩個祖先親緣關系最近的養殖點,一個是浙江義烏或蘭溪,另一個是江蘇宿遷。然后再由這兩個地方,分別向各地引種遷徙。第一條線路,從浙江義烏/蘭溪到嘉興海鹽再到嘉興南湖,但隨后這幾個地方的養殖群體不斷交流。第二條線路較復雜,從江蘇宿遷開始即分成兩支:一支先到江西鷹潭,再到浙江東陽,最后到湖北荊州的龍口和和平;另一支先到安徽安慶,隨后又分成三支,分別遷移至滁州、長豐至上饒,其中上饒的那支再到達常州。此外,浙江金華東陽的群體源自江蘇宿遷,與金華的另兩個群體不同。
2. 2 群體結構分析
基于群體的SNP位點,通過ADMIXTURE(Ale-xander et al.,2009)對克氏原螯蝦群體的群體結構進行分析(圖4)。從圖4可看出,當K=2時,對應的CV error值最低,表明14個群體可能來源于兩個祖先,該結果與系統進化分析結果吻合。另外,K=3時CV error值也較低,且與K=2時的CV error值很接近。而在系統進化分析中,浙江金華義烏和浙江金華蘭溪的群體親緣關系很近,很難區分哪個更接近祖先種,因此在一定程度上,較難區分所有群體是雙起源的還是三起源的,與群體結構的分析結果相對應。由于金華義烏與蘭溪的地理距離很近,可能本身就來自同一個群體。所以,更傾向于這些種群最初來自于兩個種源群體。
2. 3 群體主成分分析
為進一步明確群體間親緣關系,利用741147個SNP位點通過EIGENSOFT進行PCA分析(圖5)。從三維聚類后的情況看出(第一、二、三主成分分別為PC1、PC2和PC3),安徽長豐和滁州群體親緣關系較近,聚集成1個簇(Cluster);湖北荊州洪湖的2個群體及浙江東陽的群體,分別聚集成3個獨立的簇。這4個簇都游離于由剩下的9個群體形成的主簇(Main cluster)之外。在主簇中,江蘇常州和浙江金華蘭溪群體的親緣關系最遠,與系統發育的結果相似。PCA分析結果表明:湖北的2個群體,安徽的長豐和滁州及浙江東陽與其他群體間存在相對較遠的親緣關系。
2. 4 群體遺傳多樣性分析
為評估群體的遺傳多樣性,通過計算Ho、He和PIC等參數來考察各個群體的遺傳變異程度,是遺傳資源保護的基礎研究內容(Talebi et al.,2008),常用來表示群體遺傳多樣性情況(Pan et al.,2016)。從基于SNP位點計算得到14個群體的Ho、He和PIC數值統計結果(圖6)來看,Ho介于0.2171~0.2801,平均值為0.2476,He介于0.3424~0.3598,平均值為0.3534。其中,浙江嘉興南湖群體的He最高,但Ho最低,表明這個群體經過較長時間的內交,遺傳多樣性較低。14個群體的PIC均處于0.2750~0.2878,群體間相差不大,接近0.25,表明這14個養殖群體的遺傳變異,雖然還居于中等水平(劉楚吾等,2010),但已非常接近中等水平的下限。從14個群體的He和PIC來看,浙江群體的遺傳多樣性大多較低,可能會面臨更多的種質退化風險。根據PCA分析結果和遺傳多樣性分析結果,當浙江群體需要進行引種時,可選擇遺傳多樣性相對較高及單獨成簇的安徽長豐和滁州的群體。
3 討論
與傳統的微衛星等分子標記相比,通過鑒定SNP檢測遺傳多樣性,不再以DNA長度差異作為檢測手段,而是直接通過檢測序列變化,是更加高效的遺傳標記,在大多數基因組中,這是最豐富和穩定的遺傳變異形式(Liu et al.,2012)。對于缺少參考基因組的物種,特別是基因組復雜的物種來說,簡化基因組測序技術不僅節省費用,測序分析周期短,而且獲得的大量多態性位點分析結果較接近深度全基因組測序結果(胡亞亞等,2018)。以往的克氏原螯蝦群體遺傳多樣性研究中,多采用SSR標記和ISSR標記,利用的微衛星位點數很少,而且這些方法適用于個體差異較顯著的野生群體,不適用于個體差異較小的養殖群體。克氏原螯蝦作為一種重要的水產物種,人工養殖規模大,養殖群體的遺傳多樣性檢測直接與生長實踐相關聯,定期監測養殖群體對水產養殖的可持續發展極為重要。
定期檢測養殖群體的生物多樣性,有助于及時采取措施,降低種質退化風險。與國內其他重要經濟養殖品種如中華絨螯蟹、中國對蝦等相比(劉萍等,2004;馬海濤等,2007),克氏原螯蝦的遺傳多樣性相對偏低。過去對克氏原螯蝦的研究主要關注生態和引種種源方面,對克氏原鰲蝦遺傳多樣性的檢測方法研究較少。定期檢測養殖群體中遺傳多樣性水平,并適時引種是解決克氏原螯蝦產業面臨的種質退化問題的有效措施。本研究以湖北、江西、安徽、浙江和江蘇5省的克氏原螯蝦養殖群體為研究對象,通過SLAF測序,利用獲得的741147個SNP位點進行遺傳多樣性分析,結果發現14個群體的PIC在0.2750~0.2878,與其他利用微衛星標記的研究結果相比相對較低。相比于以往利用微衛星標記評估的報告結果(譚云飛等,2020),本研究計算所得的PIC值較低,推測養殖群體的遺傳多樣性已明顯低于野生群體,與此前報道關于克氏原螯蝦養殖群體的數據出入較大的原因可能是所選養殖場封閉式養殖時間較長,與其他養殖點間的交流過少,近親繁殖嚴重。當然,雜交親本的選擇還需要更多更細致的遺傳差異數據進行分析。
此外,群體的聚類分析結果表明,其系統發育關系并沒有按照長江中游、中下游和錢塘江等水系的地理位置劃分,說明養殖群體的遷移可能是人為引種或養殖的結果。與預測的群體遷移結果相似的是,從PCA分析中發現的游離在外的5個群體,均位于遷移過程的末梢。國內這些養殖點的種源相對較單一,不同養殖群體間的交流較少,許多養殖點可能已經或將要面臨因過度近親繁殖而帶來的負面影響。如成年體型越來越小,生長緩慢,抗病力降低、病害頻發、種質退化(舒新亞,2010)。近年來廣泛發生的“五月瘟”(魏文燕等,2020),也是種群遺傳多樣性降低的結果。纖毛蟲病(鐘亮等,2021)也是養殖過程中的常見病害之一,患病后,蟲體寄生在龍蝦外殼上,影響正常蛻殼生長,進而失去商業價值。因此,及時引種、提高養殖群體的遺傳多樣性,是目前解決這些問題的最便捷方式。
基于生物大數據分析,對不同地區克氏原螯蝦養殖群體的差異、群體內個體的差異進行及時調查,適時引種及加強不同地區群體間的交流,才能加強克氏原螯蝦優良種質資源的保護,且定期監測還能為引種提供重要參考依據。本研究中首次發現浙江金華東陽的群體與另外2個金華的養殖群體有較大的遺傳差異,種質來源不同,將來可作為一個就近引種的選擇。同時,也發現浙江群體的遺傳多樣性低于其他省的養殖群體,可能與其內交及與其他地區交流過少有關。為保證各地區克氏原螯蝦的遺傳多樣性,建議可適當從遺傳多樣性較高的安徽和湖北等地引種進行雜交,選育出多樣性更加豐富的群體進行繁殖,加強不同地區、不同水系間群體的交流。通過大數據監測,一方面有利于保護好現有的克氏原螯蝦物種基因庫;另一方面有利于充分利用其遺傳多樣性,挖掘育種潛力,用科學方法及時準確掌握克氏原螯蝦的種質狀況,以促進其養殖業的健康穩定發展。
4 結論
14個克氏原螯蝦養殖群體內的遺傳多樣性較低,群體間的親緣關系較接近,品種單一、長期內交跡象明顯,面臨較高的種質退化風險。通過群體的基因組重測序獲得的多態性信息遠多于傳統的微衛星方法,能以較低的成本實現對養殖群體遺傳多樣性的全面定期監測。
參考文獻:
曹玲亮,周立志,張保衛. 2010. 安徽三大水系入侵物種克氏原螯蝦的種群遺傳格局[J]. 生物多樣性,18(4):398-407. [Cao L L,Zhou L Z,Zhang B W. 2010. Genetic patterns of an invasive Procambarus clarkii population in the three river basins of Anhui Province[J]. Biodiversity Science,18(4):398-407.] doi:10.3724/SP.J.1003.2010.398.
胡亞亞,劉蘭服,冀紅柳,韓美坤,焦偉靜,高志遠,馬志民. 2018. 簡化基因組測序技術研究進展[J]. 江蘇師范大學學報(自然科學版),36(4):63-68. [ Hu Y Y,Liu L F,Ji H L,Han M K,Jiao W J,Gao Z Y,Ma Z M. 2018. Research progress on the reduced-representation genome sequencing technique[J]. Journal of Jiangsu Normal University(Natural Science Edition),36(4):63-68.] doi:10.3969/ j.issn.2095-4298.2018.04.012.
黃小芳,唐章生,劉俊丹,張宏燕,鐘一治,盧智發,侯樹鑒,王大鵬,陸專靈. 2020. 廣西不同地區克氏原螯蝦群體遺傳多樣性微衛星分析[J]. 南方農業學報,51(2):437-444.[Huang X F,Tang Z S,Liu J D,Zhang H Y,Zhong Y Z,Lu Z F,Hou S J,Wang D P,Lu Z L. 2020. Genetic diversity microsatellite analysis of Procambarus clarkii populations in different regions of Guangxi[J]. Journal of Sou-thern Agriculture,51(2):437-444.] doi:10.3969/j.issn.2095- 1191.2020.02.025.
劉楚吾,黎錦明,劉麗,郭昱嵩. 2010. 中國龍蝦微衛星標記的篩選及遺傳多樣性分析[J]. 遺傳,32(7):737-743. [Liu C W,Li J M,Liu L,Guo Y S. 2010. Screening and gene-tic diversity analysis of microsatellite markers in Chinese lobster(Panulirus stimpsoni)[J]. Hereditas,32(7):737-743.] doi:10.3724/SP.J.1005.2010.00737.
劉萍,孟憲紅,何玉英,孔杰,李健,王清印. 2004. 中國對蝦(Fenneropenaeus chinensis)黃、渤海3個野生地理群遺傳多樣性的微衛星DNA分析[J]. 海洋與湖沼,35(3):252-257. [Liu P,Meng X H,He Y Y,Kong J,Li J,Wang Q Y. 2004.? Genetic diversity in three wild populations of Fenneropenaeus chinensis in Yellow and Bohai Seas as revealed by microsatellite DNA[J]. Oceanologia et Limnologia Sinica,35(3):252-257.] doi:10.3321/j.issn:0029-814X.2004.03.009.
劉其根,李應森,陳藍蓀. 2008. 克氏原螯蝦的生物學[J]. 水產科技情報,35(1):21-23. [Liu Q G,Li Y S,Chen L S. 2008. Ecological culture of red swamp crawfish Procambarus clarkii(I)[J]. Fisheries Science & Technology Information,35(1):21-23.] doi:10.3969/j.issn.1001-1994. 2008.01.011.
馬海濤,常玉梅,于冬梅,孫效文. 2007. 利用微衛星分子標記分析四個中華絨螯蟹群體的遺傳多樣性(英文)[J]. 動物學研究, 28(2):126-133. [Ma H T,Chang Y M,Yu D M,Sun X W. 2007. Microsatellite variations among four populations of Eriocheir sinensis[J]. Zoological Research,28(2):126-133.] doi:10.3321/j.issn:0254-5853.2007.02. 003.
彭剛,劉偉杰,李佳佳,嚴維輝,唐建清. 2010. 長江流域3個克氏原螯蝦野生群體遺傳結構的微衛星分析[J]. 江蘇農業學報,26(5):1115-1117. [Peng G,Liu W J,Li J J,Yan W H,Tang J Q. 2010. Microsatellite DNA analysis of genetic structure of three wild populations of Procambarus clarkii in the Yangtze River basin[J]. Jiangsu Journal of Agricultural Sciences,26(5):1115-1117.] doi:10.3969/j.issn.1000-4440.2010.05.040.
舒新亞. 2010. 克氏原螯蝦產業發展及存在的問題[J]. 中國水產,(8):22-25. [Shu X Y. 2010. The development and existing problems of clarkiis original crayfish industry[J]. China Fisheries,(8):22-25.] doi:10.3969/j.issn.1002- 6681.2010.08.011.
宋光同,何吉祥,吳本麗,陳靜,黃龍,汪翔,武松. 2018. 克氏原螯蝦雌性生殖系統發育及組織結構觀察[J]. 江西農業學報,30(2):68-75. [Song G T,He J X,Wu B L,Chen J,Huang L,Wang X,Wu S. 2018. Study on development of female reproductive system and histological structure of Procambarus clarkii[J]. Acta Agriculturae Jiangxi,30(2):68-75.] doi:10.19386/j.cnki.jxnyxb.2018.02.16.
譚云飛,蓬國輝,熊禮靜,彭波,吳毅博,宋朝偉,白旭峰. 2020. 長江中下游流域13個克氏原螯蝦群體遺傳多樣性和遺傳結構分析[J]. 華中農業大學學報,39(2):33-39. [Tan Y F,Peng G H,Xiong L J,Peng B,Wu Y B,Song C W,Bai X F. 2020. Genetic diversity and structure ana-lysis of 13 red swamp crayfish(Procambarus clarkii)popu-lations in Yangtze River basin[J]. Journal of Huazhong Agricultural University,39(2):33-39.] doi:10.13300/j.cnki.hnlkxb.2020.02.005.
王長忠,李忠,梁宏偉,呼光富,吳勤超,鄒桂偉,羅相忠. 2009. 長江下游地區4個克氏原螯蝦群體的遺傳多樣性分析[J]. 生物多樣性,17(5):518-523.[Wang C Z,Li Z,Liang H W,Hu G F,Wu Q C,Zou G W,Luo X Z. 2009. Genetic diversity in four Procambarus clarkii populations in the lower reaches of the Yangtze River[J]. Biodiversity Science,17(5):518-523.] doi:10.3724/SP.J.1003. 2009.09017.
魏文燕,楊馬,劉家星,李良玉,吳艷蓉. 2020. 小龍蝦“五月瘟”成因及防控措施[J]. 科學養魚,(6):50-51. [Wei W Y,Yang M,Liu J X,Li L Y,Wu Y R. 2020. Causes and control measures of crayfish “May plague”[J]. Scientific Fish Farming,(6):50-51.] doi:10.3969/j. issn1004-843X. 2020.06.026.
邢智珺,姜虎成,陸偉,錢照君,于宏偉,李家樂. 2014. 江蘇8個克氏原螯蝦群體遺傳多樣性微衛星分析[J]. 上海海洋大學學報, 23(5):656-662. [Xing Z J,Jiang H C,Lu W,Qian Z J,Yu H W,Li J L. 2014. Genetic diversity analysis of eight procambarus clarkii stocks in Jiangsu Province based on microsatellites[J]. Journal of Shanghai Ocean University,23(5):656-662.]
徐濱,李忠,魏開金,馬寶珊,朱祥云,徐進. 2019. 6個克氏原螯蝦群體形態學分析[J]. 淡水漁業,49(6):27-32. [Xu B,Li Z,Wei K J,Ma B S,Zhu X Y,Xu J. 2019. Morphological variations among six populations of Procambarus clarkia[J]. Freshwater Fisheries,49(6):27-32.] doi:10.13721/j.cnki.dsyy.2019.06.005.
閆苗苗,魏光成,潘效紅,馬懷雷,李偉振. 2008. 一種適用于動物與植物總DNA提取的方法——改良CTAB法(英文)[J]. Agricultural Science & Technology,9(2):39-41. [Yan M M,Wei G C,Pan X H,Ma H L,Li W Z. 2008. A method suitable for extracting genomic DNA from animal and plant—Modified CTAB method[J]. Agricultural Science & Technology,9(2):39-41.] doi:10.16175/j.cnki. 1009-4229.2008.02.001.
嚴維輝,唐建清,許志強,李佳佳. 2019. 克氏原螯蝦大棚育苗試驗總結[J]. 水產養殖,40(12):8-9. [Yan W H,Tang J Q,Xu Z Q,Li J J. 2019. Summary of greenhouse seedling experiment of Procambarus clarkii[J]. Journal of Aquaculture,40(12):8-9.] doi:10.3969/j.issn.1004-2091. 2019.12.005.
張愛軍,沈繼紅. 2005. 龍蝦的綜合加工利用[J]. 中國資源綜合利用,(9):35-36. [Zhang A J,Shen J H. 2005. The comprehensive machining utilization of Chinese lobster[J]. China Resources Comprehensive Utilization,(9):35-36.] doi:10.3969/j.issn.1008-9500.2005.09.011.
張莎,俞樹惠,邱高峰. 2018. 克氏原螯蝦染色體及其核型[J]. 水產學報,42(10):1513-1519. [Zhang S,Yu S H,Qiu G F. 2018. Chromosome and karyotype of the crayfish(Procambarus clarkii)[J]. Journal of Fisheries of China,42(10):1513-1519.] doi:10.11964/jfc.20171211084.
鐘亮,黃小麗,徐銘,李良玉. 2021. 小龍蝦纖毛蟲病的診斷與防治[J]. 科學養魚,(1):50-51. [Zhong L,Huang X L,Xu M,Li L Y. 2021. Diagnosis and control of ciliosis in crayfish[J]. Scientific Fish Farming,(1):50-51.] doi:10.3969/j.issn1004-843X.2021.01.027.
Alexander D H,Novembre J,Lange K. 2009. Fast model-based estimation of ancestry in unrelated individuals[J]. Genome Research,19(9):1655-1664. doi:10.1101/gr.094 052.109.
Barbaresi S,Fani R,Gherardi F,Mengoni A,Souty-Grosset C. 2003. Genetic variability in European populations of an invasive American crayfish:Preliminary results[J]. Biolo-gical Invasions,5(3):269-274. doi:10.1023/A:10261335 19707.
Barbaresi S,Gherardi F,Mengoni A,Souty-Grosset C. 2007. Genetics and invasion biology in fresh waters:A pilot study of Procambarus clarkii in Europe[M]//Gherardi F. Biological invaders in inland waters:Profiles, distribution, and threats. Dordrecht:Springer:381-400.doi:10.1007/978-1-4020-6029-8_20.
Botstein D,White R L,Skolnick M,Davis R W. 1980. Construction of a genetic linkage map in man using restriction fragment length polymorphisms[J]. American Journal of Human Genetics,3(3):314-331.
Davidson R,Campo A M D. 2020. Combinatorial and computational investigations of neighbor-joining bias[J]. Frontiers in Genetics,(11):584785. doi:10.3389/fgene.2020. 584785.
Kumar S,Stecher G,Li M,Knyaz C,Tamura K. 2018. MEGA X:Molecular evolutionary genetics analysis across computing platforms[J]. Molecular Biology and Evolution,35(6):1547-1549. doi:10.1093/molbev/msy096.
Li H,Durbin R. 2009. Fast and accurate short read alignment with Burrows-Wheeler transform[J]. Bioinformatics,25(14):1754-1760. doi:10.1093/bioinformatics/btp324.
Li H,Handsaker B,Wysoker A,Fennell T,Ruan J,Homer N,Marth G,Abecasis G,Durbin R. 2009. The sequence alignment/map format and SAMtools[J]. Bioinformatics,25(16):2078-2079. doi:10.1093/bioinformatics/btp352.
Liu J,Huang S M,Sun M Y,Liu S Y,Liu Y M,Wang W X,Zhang X R,Wang H Z,Hua W. 2012. An improved allele-specific PCR primer design method for SNP marker analysis and its application[J]. Plant Methods,8(1):34. doi:10.1186/1746-4811-8-34.
Mckenna A,Hanna M,Banks E,Sivachenko A,Cibulskis K,Kernytsky A,Garimella K,Altshuler D,Gabriel S,Daly M,DePristo M A. 2010. The Genome Analysis Toolkit:A MapReduce framework for analyzing next-generation DNA sequencing data[J]. Genome Research,20:1297-1303. doi:10.1101/gr.107524.110.
Pan Y Z,Wang X Q,Sun G L,Li F S,Gong X. 2016. Application of RAD sequencing for evaluating the genetic diversity of domesticated Panax notoginseng(Araliaceae)[J]. PLoS One,11(11):e0166419. doi:10.1371/journal.pone.0166419.
Price A L,Patterson N J,Plenge R M,Weinblatt M E,Shadick N A,Reich D. 2006. Principal components analysis corrects for stratification in genome-wide association studies[J]. Nature Genetics,38(8):904-909. doi:10.1038/ ng1847.
Seplyarskiy V B,Kharchenko P,Kondrashov A S,Bazykin G A. 2012. Heterogeneity of the transition/transversion ratio in Drosophila and hominidae genomes[J]. Molecular Biology and Evolution,29(8):1943-1955. doi:10.1093/molbev/mss071.
Sun X W,Liu D Y,Zhang X F,Li W B,Liu H,Hong W G,Jiang C B,Guan N,Ma C X,Zeng H P,Xu C H,Song J,Huang L,Wang C M,Shi J J,Wang R,Zheng X H,Lu C Y,Wang X W,Zheng X K. 2013. SLAF-seq:An efficient method of large-scale De Novo SNP discovery and genotyping using high-throughput sequencing[J]. PLoS One,8(3):e58700. doi:10.1371/journal.pone.0058700.
Talebi R,Naji A M,Fayaz F. 2008. Geographical patterns of genetic diversity in cultivated chickpea(Cicer arietinum L.)characterized by amplified fragment length polymorphism[J]. Plant,Soil and Environment,54(10):447-452. doi:10.17221/399-PSE.
Yi S K,Li Y H,Shi L L,Zhang L,Li Q B,Chen J. 2018. Characterization of population genetic structure of red swamp crayfish,Procambarus clarkii,in China[J]. Scientific Reports,8(1):5586. doi:10.1038/s41598-018-23986-z.
Zhong Y Z,Tang Z S,Huang L M,Wang D P,Lu Z L. 2020. Genetic diversity of Procambarus clarkii populations based on mitochondrial DNA and microsatellite markers in different areas of Guangxi,China[J]. Mitochondrial DNA Part A,31(2):48-56. doi:10.1080/24701394.2020. 1721484.
(責任編輯 王 暉)
