熒光蛋白mNeonGreen,mKOk作為FRET傳能對在研究植物蛋白質互作中的應用分析
2021-04-30 09:03:12王亞琴李宗迪李晨羊李云琴胡濤謝禮周雪平
植物保護 2021年2期
王亞琴 李宗迪 李晨羊 李云琴 胡濤 謝禮 周雪平
關鍵詞蛋白質互作;熒光共振能量轉移;mNeonGreen;mKOk
熒光共振能量轉移技術(fluorescence reso-nance energy transfer,FRET)是指兩個相互靠近的熒光發色基團中,一個分子吸收一定頻率的光子被激發到更高的電子能態的同時,通過偶極子相互作用,將能量向另一分子轉移(即發生能量共振轉移)的現象。這種現象的發生有兩個前提條件:一是兩個熒光發光基團中一個熒光基團(供體分子)的發射光波長需與另一個熒光基團(受體分子)的激發光波長存在重疊區域;二是供體分子和受體分子之間的距離必須在1~10nm之間。由于熒光共振能量轉移現象需要在這兩個較為苛刻的條件下才能發生,且對供體和受體間的距離和空間取向高度敏感,因而可以作為檢測蛋白質之間直接相互作用的有效工具。
青色熒光蛋白(cyan fluorescent protein,CFP)與黃色熒光蛋白(yellow fluorescent protein,YFP)是目前蛋白質相互作用研究中應用最廣泛的FRET對,但CFP/YFP對存在一定的不足,如CFP和YFP會發生快速、多速率、可逆的光漂白,YFP可在一定條件下光轉化為青色熒光,CFP可以被YFP的激發光波長激發出熒光等原因造成CFP/YFP對背景干擾較大。此外,許多報道表明基于CFP和YFP的FRET,當生物化學反應微弱或比較短暫時,會對FRET檢測效率產生一定的挑戰。因此,找到一對低背景干擾且能量轉移效率高的熒光蛋白對,對于FRET的檢測至關重要。
mNeonGreen是一種來源于文昌魚Branchios-toma lanceolatum的新型綠色熒光蛋白,最大激發光和發射光的波長分別為504、517 nm,mKOk是一種來源于石珊瑚Verrillofungia concinna的新型橙色熒光蛋白,最大激發光波長和發射光波長分別為551、563 nm,研究發現,mNeonGreen和mKOk熒光蛋白相較于傳統的綠色和橙色熒光蛋白具有更強的熒光強度。目前關于將這兩種熒光蛋白用于FRET檢測還未見報道。基于此,本研究在植物細胞中驗證mNeonGreen-mKOk熒光蛋白對是否適用于FRET技術檢測,并探索一套基于FRET技術檢測植物蛋白間相互作用的體系。
1材料與方法
1.1材料
本氏煙Nicotiana benthamiana野生株系型由本實驗室留種繁殖;大腸桿菌Escherichia coli菌株DH5a,農桿菌Agrobacterium tume faciens菌株C58C1,含FLAG和MYC標簽的植物表達載體pGD-FLAG、pGD-MYC均由本實驗室保存;REMl.1為實驗室保存質粒;mKOk、mNeonGreen熒光蛋白由尚亞生物有限公司合成;FLAG、MYC標簽抗體購自Sig-ma公司;二抗購自ABclonal公司;KOD OneTM PCRMaster Mix購自TOYOBO公司;ClonExpress II OneStep Cloning Kit、ClonExpress MultiS One Step Clo-ning Kit購自Vazyme公司;Kan、Rif抗生素購自上海生工生物工程有限公司。
1.2方法
1.2.1 FRET載體構建
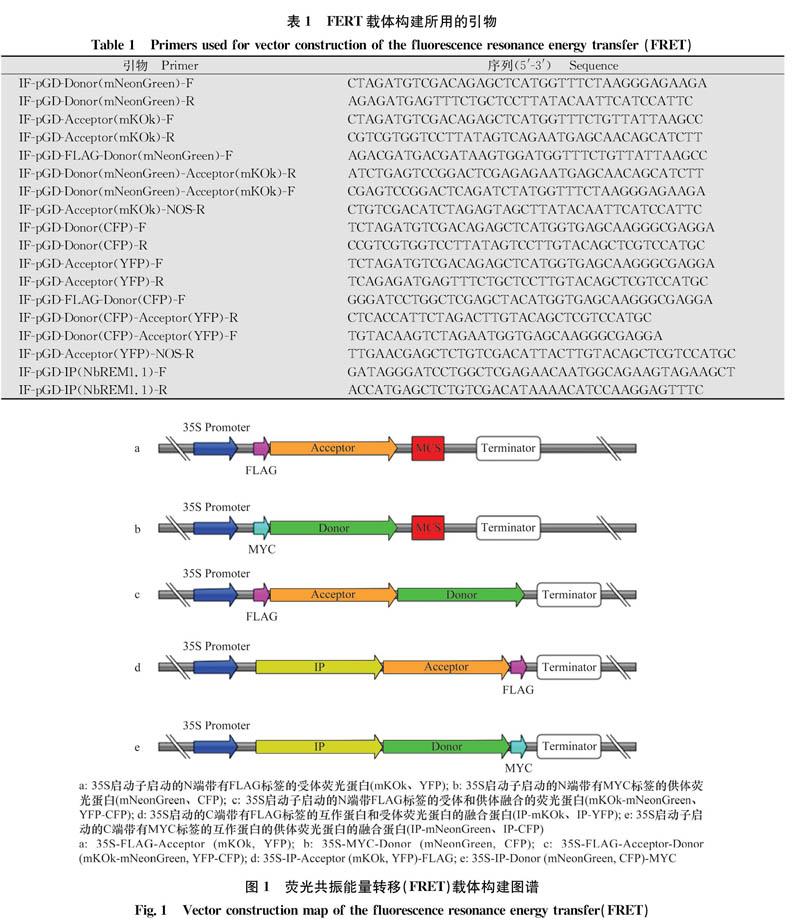
以供體熒光蛋白(donor)和受體熒光蛋白(accep-tor)為模板,分別設計特異性引物IF-pGD-Donor-F/IF-pGD-Donor-R、IF-pGD-Acceptor-F/IF-pGD-Accep-tor-R(表1),得到去除終止密碼子且帶有與pGD載體同源序列的基因片段,而后將得到的PCR產物利用單片段同源重組的方法分別克隆到pGD-35S-MYC和pGD-35S-FLAG載體上,得到pGD-35S-MYC-Donor和pGD-35S-FLAG-Acceptor(圖1a,b)。再以Donor和Acceptor為模板,分別設計特異性引物IF-pGD-FLAG-Donor-F/IF-pGD-Donor-Acceptor-R、IF-pGD-Donor-Acceptor-F/IF-pGD-Acceptor-NOS-R,得到帶有同源序列的Donor和Acceptor片段,而后將PCR產物利用多片段同源重組的方法克隆到pGD-35S-FLAG載體上,得到載體pGD-35S-FLAG-Acceptor-Donor(圖1c)。以待檢測的互作蛋白(interacting protein,IP)為模板,設計特異性引物IF-pGD-IP-F/IF-pGD-IP-R,得到去除終止密碼子且帶有與載體同源序列的基因片段,再利用同源重組的方法將PCR產物分別克隆到帶有供體和受體熒光蛋白的載體上,得到載體pGD-35S-IP-Ac-ceptor-FLAG、pGD-35S-IP-Donor-MYC(圖1d,e)。構建好的載體在測序正確后,電激轉化到農桿菌菌株C58C1中。
1.2.2農桿菌在本氏煙上的瞬時表達
將構建好的農桿菌置于含有Kanq-Rif的雙抗YFP液體培養基中,28℃,220r/min培養12 h,取2mL菌液于12000r/min下離心30s,而后加入1mL潤緩沖液重懸菌體,調節OD至0.8左右,室溫靜置2~3h。靜置好的菌液分別浸潤本氏煙葉片,于25℃下培養36~48h。
1.2.3蛋白提取、SDS-PAGE及Western blot分析
取約0.10g本氏煙葉片,經液氮處理后研磨成粉末,加入200μL植物蛋白質提取緩沖液,振蕩1min使樣品與提取緩沖液充分混合反應,12000 r/min離心5min,取上清,加入50/μ5×loading buffer,95℃金屬浴5min后使蛋白變性。將制備好的蛋白樣品進行SDS-PAGE電泳,電泳結束后,取下凝膠,轉至硝酸纖維素膜(NC膜)上,用TBST緩沖液對封閉后的膜洗3遍,每次5min,加人事先配制好的一抗溶液,在搖床上輕晃孵育1h,用TBST緩沖液洗膜3遍,每次5~10min。加入HRP標記的二抗,在搖床上孵育1h,用TBST緩沖液洗膜3~5次,每次5min,最后用Luminate Forte WesternHRP substrate(Millipore)進行化學顯影。
1.2.4激光共聚焦顯微鏡檢測FRET效率
樣品制備:檢測樣品為共表達供體熒光蛋白mNeonGreen/CFP-IP和受體熒光蛋白mKOk/YFP-IP及融合表達mNeonGreen-mKOk、CFP-YFP的本氏煙葉片;質控樣品為單獨表達供體熒光蛋白mNeonμGreen/CFP-IP或受體熒光蛋白mKOk/YFP-IP的本氏煙葉片。采用敏化發射法(sensitized emissionFRET,SE-FRET)檢測FRET現象。該法是通過供體的激發光去激發供體,若供體和受體的空問距離在1~10nm范圍內,供體的發射光就能激發出受體的熒光,則可以檢測到FRET信號。檢測步驟如下:
第一步:調試參數。首先設定供體熒光和受體熒光的掃描參數。由于mNeonGreen供體熒光的激發峰和發射峰分別為504、517nm mKOk受體熒光的激發峰和發射峰分別為551、563 nm,因此采用488 nm波長的激發光激發mNeonGreen熒光蛋白,并采用490~550nm波長的發射光接收發射光;對于受體mKOk熒光蛋白,采用561nm波長的激發光進行激發,并采用575~625nm波長的發射光檢測發射光信號。另外,當FRET熒光蛋白對變為CFP/YFP時,由于CFP供體熒光的激發峰和發射峰分別為436、476 nm,YFP受體熒光的激發峰和發射峰分別為516、529nm,因此采用405nm波長的激發光可以激發出CFP的熒光,并采用455~515nm波長的發射光接收發射光,YFP的熒光可以采用514nm波長的激發光進行激發,采用520~620nm波長的發射光進行檢測。
第二步:采用多軌道(multi track)FRET圖像采集模式。其中,Track中有兩個通道,待檢測的蛋白均用供體的激發光進行激發,通道工收集供體熒光的發射信號,通道Ⅱ收集FRET的發射信號。TrackⅡ只有一個受體熒光的通道,采用受體的激發光進行激發,并使用這個通道單獨收集受體熒光的發射信號。調整激發光的強度,獲得理想的熒光圖像。
第三步:FRET效率的分析。從獲得的FRET熒光圖像中,選擇感興趣的區域(ROD,采用Xia和Liu的方法,利用卡爾蔡司AIM軟件的LSMFRET軟件自動計算出每個ROI的FRET效率,即N-FRET值。
第四步:統計學分析。采用t檢測的方法分析試驗組和對照組的N-FRET值是否存在差異,利用GraphPad Prism 7軟件進行統計分析。
2結果與分析
2.1熒光蛋白mNeonGreen、mKOk的光譜分析
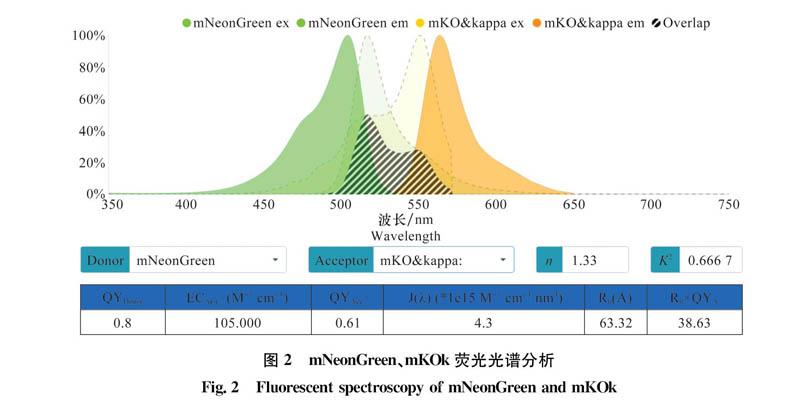
通過在線網站(https://WWW.fpbase.org/fret/)對mNeonGreen和mKOk的熒光光譜進行分析,結果顯示,mNeonGreen最大激發光和發射光的波長分別為504、517 nm,mKOk最大激發光波長和發射光波長分別為551、563nm,且二者的光譜存在重疊區域(圖2),其中mNeonGreen可作為供體熒光,mKOk作為受體熒光。
2.2重組載體鑒定
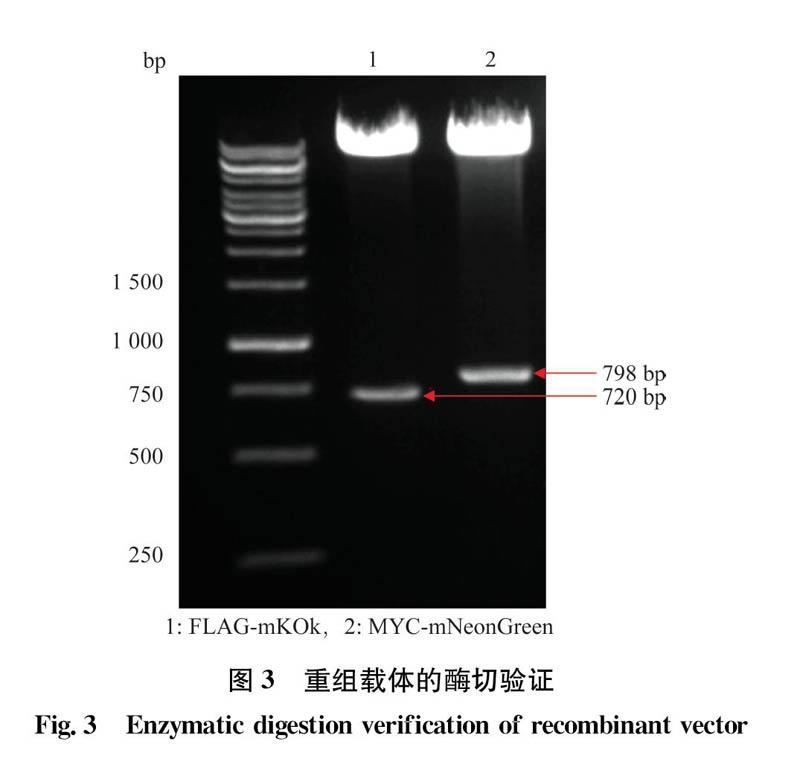
以熒光蛋白mNeonGreen,mKOk為模板,利用設計的特異性引物,通過PCR擴增和酶切篩選獲得的陽性克隆,經測序驗證正確后,對相應的目的片段進行雙酶切,酶切產物經1%的瓊脂糖凝膠電泳,得到大小分別為798 bp和720 bp的特異性條帶(圖3),與已報道的目標基因大小符合,證明熒光蛋白mNeon-Green.mKOk分別成功地插入到目的載體上。
2.3重組蛋白鑒定
將構建好的熒光蛋白載體在本氏煙葉片上進行瞬時表達,36~48h后提取蛋白并進行Westernblot分析,利用MYC和FLAG抗體分別進行檢測,結果顯示,MYC和FLAG抗體可檢測到mNeon-Green和mKOk的條帶,用FLAG抗體可檢測到融合蛋白的條帶(圖4),證實所構建的熒光蛋白載體可以在本氏煙葉片中成功表達。
2.4 mNeonGreen/mKOk熒光蛋白對適用于FRET技術檢測
將C58Cl-pGD-mNeonGreen-MYC、C58Cl-pGD-mKOk-FLAG等比例混合后浸潤野生型本氏煙,作為陰性對照組,將融合表達載體C58C1-pGD-FLAG-mNeonGreen-mKOk浸潤野生型本氏煙葉片,作為陽性對照組,在本氏煙葉片上瞬時表達48h后進行FRET效率檢測。
為了排除串色對FRET效率計算的干擾,首先收集僅表達供體mNeonGreen和受體mKOk的FRET圖像作為校正因子(圖5a,b),計算出串擾系數,用于后續的FRET效率分析,該計算過程由卡爾蔡司自帶的LSM FRET軟件完成。
用波長為488nm激發光和490~550nm發射光檢測共表達供體mNeonGreen和受體mKOk以及表達融合蛋白mNeonGreen-mKOk的本氏煙葉片,均可以獲得mNeonGreen熒光圖像(圖5c,d,mNeonGreen通道);用波長561nm激發光和575~625nm發射光檢測共表達供體mNeonGreen和受體mKOk以及表達融合蛋白mNeonGreen-mKOk的本氏煙葉片,也可檢測到mKOk熒光的發射光信號(圖5c,d,mKOk通道)。用波長為488nm激發光和575~625nn3.發射光檢測FRET信號時,無法在共同表達供體和受體的葉片(mNeonGreen+mKOk)中獲得FRET通道的圖像(圖5c,FRET通道),但能在表達融合蛋白mNeonGreen-mKOk的葉片上檢測到較強的FRET信號(圖5d,FRET通道)。
根據獲得的FRET圖像,用卡爾蔡司自帶的LSM FRET軟件計算FRET效率N-FRET值。結果顯示,陰性對照組的FRET效率值幾乎為零,而陽性對照組的FRET效率值遠遠高于陰性對照組,且兩者存在顯著差異(圖5e)。說明mNeonGreen和mKOk熒光蛋白在共浸潤的情況下,由于空間距離上足夠遠,不會發生FRET現象,可以作為FRET檢測的陰性對照組。而融合熒光蛋白mNeonGreen-mKOk由于在空間距離上幾乎為零,因此發生了FRET現象,說明mNeonGreen/mKOk熒光蛋白對適用于FRET技術檢測。
2.5mKOk/mNeonGreen熒光蛋白對的優勢
將C58Cl-tGDMYC-CFP、C58C1-pGDFLAG-YFP等比例混合后浸潤野生型本氏煙,作為陰性對照組,將融合表達載體C58C1-pGD-FLAG-CFP-YFP浸潤野生型本氏煙,作為陽性對照組,在本氏煙葉片上瞬時表達48h后進行FRET效率檢測。
首先,采集單獨表達供體CFP和受體YFP的FRET圖像(圖6a,b),并進行串擾系數的計算。再用波長405 nm激發光、455~515nm發射光檢測共表達供體CFP和受體YFP以及表達融合蛋白CFP-YFP的本氏煙葉片,均可以獲得青色熒光圖像(圖6c,d,CFP通道);用波長514nm激發光和520~620nm發射光檢測共表達供體和受體以及表達融合蛋白的本氏煙葉片,均能檢測到黃色熒光的發射光信號(圖6c,d,YFP通道)。用波長為405nm激發光和520~620nm發射光檢測FRET信號時,不僅能在表達融合蛋白CFP-YFP的葉片上檢測到FRET的信號(圖6d,FRET通道),在共同表達CFP和YFP的葉片中,也可以獲得較弱的FRET通道信號(圖6c,FRET通道)。
對獲得的陰性和陽性對照組進行FRET效率分析,結果表明,在共同表達CFP和YFP的情況下,FRET通道檢測到了較弱的信號,但是通過統計分析,表達融合蛋白CFP-YFP的葉片FRET效率相比于共同表達CFP和YFP的葉片FRET效率之問還是存在顯著性差異的(圖6e)。與mNeonGreen/mKOk熒光對的陰性對照組相比,CFP/YFP熒光對陰性對照組FRET效率值明顯偏高,說明CFP/YFP熒光對存在一定的背景干擾,而mNeonGreen/mKOk的陰性對照組的FRET效率值更接近于零,背景干擾較低。所以,在煙草葉片中,mNeonGreen/mKOk相比于傳統的CFP/YFP更適合作為FRET熒光蛋白對用于蛋白相互作用的檢測。
2.6 mKOk-mNeonGreen熒光蛋白對驗證植物蛋白間相互作用
Remorin蛋白是定位在植物細胞膜上的一種陸地植物特有的蛋白,它可以通過影響胼胝體的積累來調節胞間連絲孔徑的大小,在植物抵抗病原微生物過程中起著至關重要的作用。NbREMl.1是Remorin蛋白的一種,Fu等利用傳統的酵母雙雜交、雙分子熒光互補等方法鑒定到NbREMl.1存在自身互作的現象。因此,本研究選取REMl.1蛋白,采用mNeonGreen/mKOk FRET熒光蛋白對驗證其蛋白質間相互作用。
采集單獨表達REMl.1-mNeonGreen和REM1.1-mKOk的FRET圖像(圖7a,b),計算出串擾系數。再用488nm波長的激發光、490~550nm波長的發射光檢測共表達供體mNeonGreen和受體mKOk以及共表達REMl.1-mNeonGreen和REMl.1-mKOk的煙草細胞,均可以獲得mNeon-Green熒光圖像(圖7c,d,mNeonGreen通道)。同時,用561nm波長的激發光、575~625nm波長的發射光檢測共表達供體mNeonGreen和受體mKOk以及共表達REMl.1-mNeonGreen和REMl.1-mKOk的煙草細胞,均可以獲得mKOk熒光的發射光信號(圖7c,d,mKOk通道)。用波長為488 nm波長的激發光、575~625nm波長的發射光檢測FRET信號時,無法在共同表達供體mNeonGreen和受體mKOk的葉片中獲得FRET通道的圖像(圖7c,FRET通道),但能在共表達REMl.1-mNeon-Green和REMl.1-mKOk的煙草葉片上檢測到較強的FRET信號(圖7d,FRET通道)。
根據獲得的FRET圖像,利用軟件計算FRET效率值。結果顯示,共同表達REMl.1-mNeon-Green和REMl.1-mKOk的FRET效率值遠高于共表達mNeonGreen和mKOk的FRET效率值,并且兩者之間存在顯著性差異(圖7e),證實REMl.1可以發生自身互作,與之前文獻報道的結果一致。
3討論
在植物體內,蛋白質間的相互作用與多種生物過程相關,包括信號轉導、體內平衡控制、應激反應、植物防御和器官形成等。在分子層面上,蛋白質問的相互作用在蛋白質翻譯后修飾、轉錄輔助因子的招募、細胞骨架的組裝、轉運體的活化等方面都具有重要意義。
目前研究蛋白質互作的常規技術為酵母雙雜交技術、雙分子熒光互補技術(BiFC)、GST pull-down技術、免疫共沉淀技術等。這些研究技術的不斷發展,為蛋白一蛋白間相互作用的研究提供了極為有利的條件,但同時這些研究手段也存在不少缺陷,如傳統的酵母雙雜交系統操作周期較長,并且需要將誘餌蛋白和捕獲蛋白拉至細胞核內才能啟動下游報告信號,因此不能鑒定出在細胞核之外具有特定亞細胞定位的互作。BiFC的缺點在于假陽性較多,需要進行多重篩選等試驗進行排除。GST pull-down技術無法進行大規模篩選、且不能排除內源蛋白產生的假陽性的干擾。免疫共沉淀技術無法檢測瞬時的相互作用、無法判斷是直接還是問接的相互作用等。
近年來,FRET技術的發展為彌補上述問題提供了一個有力的工具。相比于傳統的蛋白一蛋白互作研究手段,FRET技術能夠在活細胞的正常生理條件下進行檢測,并具有靈敏度高,假陽性較低,實驗周期短的優勢,因此目前FRET技術在研究哺乳動物蛋白之間相互作用上已經得到了廣泛的應用。但由于植物葉片葉肉組織結構復雜等因素,FRET技術在植物蛋白互作研究中的應用較少。
進行FRET檢測時最關鍵的就是選擇兩個合適的熒光蛋白(供體和受體),目前應用較廣泛的是CFP和YFP,由于CFP和YFP都是在GFP的基礎上改造得來的,兩者在一定程度上可能存在著同源性,可形成微弱的相互作用,同時YFP的激發光也可以激發出CFP的熒光,這些因素導致CFP/YFP熒光對的背景干擾較大,對試驗結果產生一定的影響。本研究選取了兩種新型的熒光蛋白mNeon-Green/mKOk作為FRET傳能對,熒光光譜分析表明mKOk和mNeonGreen的光譜存在重疊區域,且兩者之間可以發生熒光共振能量轉移,其中mNeon-Green作為供體熒光,mKOk作為受體熒光。與傳統的傳能對CFP/YFP相比,mNeonGreen/mKOk背景干擾較低,更適合作為FRET熒光蛋白對用于蛋白相互作用的檢測。通過構建并表達熒光蛋白與目標蛋白的融合蛋白,利用FRET技術成功地在植物細胞中驗證了蛋白與蛋白間的相互作用。
FRET技術在研究活細胞單分子之間相互作用的能力已引起人們的廣泛興趣,這項技術現在已經成為一種顯微鏡研究的常用技術。盡管如此,用FRET檢測蛋白質互作的報道還非常有限,這表明目前FRET技術還遠非一種常規技術。在對FRET效率進行檢測的過程中,檢測儀器的敏感度、分辨率、參數設置以及計算機影像采集和分析軟件的能力等都是影響FRET效率的因素。目前,FRET技術在植物學研究中的應用非常少,植物細胞的結構與動物細胞有較大差別,熒光蛋白的選擇,ROI的選取,融合蛋白在葉肉細胞中的表達水平等都會影響FRET的效率和試驗的成敗。本研究采用敏化發射法分析FRET效率,這種方法對于激光共聚焦顯微鏡的配置要求較低,操作簡單,但是該法只能定性地判斷2個蛋白間是否存在相互作用,無法定量比較蛋白一蛋白相互作用的強弱。就目前而言分析蛋白質間的相互作用需要結合多種不同的方法相互取長補短才能得出一個科學的結論。
本研究成功構建了mNeonGreen和mKOk兩個熒光蛋白標記的載體,建立了一套基于FRET技術在植物細胞中進行蛋白一蛋白相互作用的體系,包括融合表達載體的構建和表達、激光共聚焦的圖像采集、FRET效率的分析等,為后續FRET技術在植物細胞中的應用提供了更多的技術支持和借鑒。
